1. Introduction
The emergence of Zika virus (ZIKV) in 1947 from Rhesus monkey in the Zika forest of Uganda has resulted in new challenges to global public health. Transmission of ZIKV in French Polynesia and South America in previously infection-naïve territories, has been associated with the development of pathologic neurological symptoms and central nervous system abnormalities in the human fetuses including microcephaly [
1,
2,
3,
4]. Viral persistence in the infected hosts has long been recognized as a common theme in flavivirus biology. Persistently flavivirus-infected cells hidden in tissues are often difficult to detect and may lead to devastating medical consequences. For example, the transmission of West Nile Virus (WNV), another flavivirus, from WNV-infected organ donors who were evidently no longer viremic during blood testing to recipients through organ transplantation is well-documented [
5,
6]. To effectively prevent ZIKV transmission through transfusion of infected blood and particularly, through transplantation of infected organs or tissues, a better understanding of the human cells that could potentially be harboring the infectious virus and serving continuously as a ZIKV reservoir over a prolonged time is needed. The dynamic process of ZIKV clonal selection with specific gene mutations during persistent viral infection and the adaptive mechanisms of human host cells that confer ZIKV persistence will also need to be further explored.
In a study of experimentally ZIKV-infected Rhesus monkeys, ZIKV RNA (v-RNA) was found to persist with the highest levels in hemato-lymphatic tissues, such as lymph nodes and spleen, after they were no longer viremic [
7]. Two other studies involving experimentally ZIKV-infected rhesus monkeys also reported that lymphoid tissues and central nervous system (CNS) cells had prolonged viral persistence [
8,
9]. To investigate possible ZIKV persistence in cells of the hemato-lymphatic system and blood, our laboratory has examined the relative susceptibility of human hematopoietic cell lines with different developmental characteristics to infection by the prototype MR 766 ZIKV strain with a focus on the duration as well as the outcome(s) of the infection [
10]. Our study revealed that human B lymphoblastoid cell lines were highly susceptible to ZIKV infection and developed prominent cytopathic changes and fulminant cell necrosis in the culture, while T lymphoblastoid cell lines were very resistant to the ZIKV infection [
10]. We reported the human monocytic leukemia/histiocytic lymphoma-originated U937 cell line [
11], initially showing a slow rate and only low-grade infection using the ZIKV MR 766 strain, could develop into persistently ZIKV-infected cell lines in which most, if not all, of the cells in the culture were positive for ZIKV antigens despite no apparent cytopathological changes [
10]. The ZIKV-infected U937 cell cultures produced ZIKV RNA (v-RNA) and infectious ZIKV persistently (persistent ZIKVs) with distinct infectivity and pathogenicity when tested using various kinds of host cells, in comparison with the original MR766 strain ZIKV [
10]. We conducted comparative studies between the original/prototype ZIKVs produced by the acutely infected human U937 cells and the persistently ZIKV-infected human U937 cells, and discussed the genomic variations developed in the persistent ZIKVs [
10].
In this study, we similarly examined the relative susceptibility of human hematopoietic cell lines with various developmental characteristics to infection by the prototype ZIKV strains isolated during the contemporary epidemics in America with a focus on the duration as well as the outcome(s) of viral infection. We found that the American isolates of the PRV (Puerto Rico) and FLR (Colombia) ZIKV strains not only failed to infect human T lymphoblastoid cell lines, but also failed to infect human hematopoietic monocytic/histiocytic U937 cells. On the other hand, our study revealed the two American ZIKV strains like the prototype MR 766 ZIKV strain could effectively infect cultured HEK293 human epithelial cell line with documented kidney and neuronal differentiation properties [
12].
Consistent with a previous report and the observations that the prototype African ZIKV isolate MR 766 appeared to be more infectious and cytopathogenic to various human cells than the ZIKVs circulating in the Americas [
7,
11], MR 766 strain ZIKV-infected HEK293 cells completely sloughed off from the flask after developing fulminant cytopathic effects (CPE) and cytolysis in culture. In contrast, small populations of HEK293 cells infected with the PRV and FLR strains of ZIKVs continued to survive the virus-induced cytocidal or cytopathic effects and regrew when they were provided with fresh culture media in the parallel study. Significantly, regrowth of the surviving HEK293 cells in culture resulted in HEK293 cell lines that were persistently infected with the PRV and FLR strains of ZIKV and constitutively producing ZIKV v-RNA genomes and infectious ZIKV virions. We again conducted comparative studies involving genomic variations and alterations of infectivity and cytopathogenicity between persistent ZIKVs produced by the persistently ZIKV-infected human HEK293 cells and the original inoculum or PRV and FLR strains of ZIKV produced by the acutely infected HEK293 cells.
3. Discussion
The recent ZIKA virus epidemic is associated with fetal brain lesions and other serious birth defects (including microcephaly, fetal growth restriction, stillbirth, ocular disorders, and CNS injury, etc.) classified as congenital ZIKV syndrome. Postnatal ZIKV infection in infants and children has been reported, and one study has shown that postnatal ZIKV infection is associated with persistent abnormalities in brain structure, function, and behavior in infant macaques [
15]. However, there is no scientific documentation of ZIKV-associated birth defects in Africa, where the virus historically originated. The study by Zhang et al. claimed that microcephalies caused by American isolates/strains of ZIKV in neonatal mice are more severe compared to that caused by an old Asian strain of ZIKV [
16].
Previous studies conducted by our laboratory showed that the prototype MR 766 strain ZIKV initially developed only a low-grade infection at a slow rate in the human monocytic/histiocytic U937 cell line without causing CPE changes or cytolysis could lead to ZIKV persistency. The continuously growing persistently ZIKV-infected U937 cell lines were constantly producing ZIKV v-RNA genomes and infectious virions with alterations of infectivity and cytopathogenicity [
10]. Our current study revealed the American isolates including the PRV and FLR ZIKV strains failed to infect the human hematopoietic U937 cell line. However, the American PRV and FLR ZIKV isolates could effectively infect human epithelial HEK293 cell lines derived from human embryonic kidney cells known of exhibiting many properties of immature neurons [
12]. Infections by the PRV and FLR ZIKV strains quickly resulted in prominent CPE changes and extensive cytolysis in cultures of HEK293 cells. Interestingly, persistently ZIKV-infected HEK293 cell lines could also be established from PRV and FLR strains of ZIKV-infected cultures by providing the few surviving cells in the ZIKV-infected cultures with fresh media. The persistently ZIKV-infected HEK293 cell lines also constantly produced ZIKV v-RNA genome and infectious virions in culture. Different from the persistently MR 766 strain ZIKV-infected U937 cells previously established, some of the cells in the continuously growing PRV and FLR strains of ZIKV-infected HEK293 cell cultures would evidently continue to undergo apoptosis.
The findings of only a small population of PRV and FRL strains ZIKV-infected HEK293 cells could survive and upon favorable conditions, would gradually propagate to confluency in culture over time were reproduced multiple times. The growing HEK293 cells that survived infections of these ZIKV strains in the culture were found persistently infected by the virus with positive IFA for Zika viral specific antigen. A similar finding was reported previously in the other flavivirus-infected cultures of HEK293 cells and Vero cells [
17,
18]. The present study was conducted comparatively on persistent ZIKVs produced from a set of the persistently American strains ZIKV-infected HEK293 cell lines established in culture and then followed closely for more than 11 months. We compared the infectivity and cytopathogenicity of the persistent ZIKVs produced by persistently ZIKV-infected HEK293 cells with the parental/original ZIKVs produced by the acutely ZIKV-infected HEK293 cells to Vero cells (
Figure 5), naïve HEK293 cells (
Figure A1) and various human epithelial cell lines (HeLa cell line and A549 cell line, data not shown). The persistent PRV and FLR ZIKV strains had evidently attenuated cell infectivity and cytopathogenicity compared to the original inoculum PRV and FLR ZIKV strains of ZIKVs. Our previous study showed that the persistent ZIKVs produced by persistently MR766 strain ZIKV-infected human U937 cells established in three independent studies had properties not only distinct from the original inoculum ZIKV, but also among themselves in terms of specificity of cell infection and cytopathogenicity [
10]. Importantly, in contrast to the finding in the present study, all the persistent ZIKVs produced by the persistently MR 766 strain ZIKV-infected U937 cell lines developed in the three studies of 1st round of ZIKV persistence were found highly infectious and cytopathogenic to the naïve U937 cells. Genomic analyses of ZIKVs showed that in comparison to the original MR 766 ZIKV strain, the persistent MR 766 strains were three rather uniform subclones of ZIKVs, each with distinct sets of mutations in various key genes enabling the selected viruses to continuously infect and proliferate in U937 cells with apparently activated anti-viral mechanisms [
10].
Establishment of persistence in ZIKV-infected HEK293 cells resembles more closely to the establishment of the second round MR766 strain ZIKV persistence, which was achieved by infecting the naïve U937 cells with the three persistent ZIKVs obtained in the first round of ZIKV persistence in U937 cells. Differing from a low rate and low-grade infection by the original/parental MR766 strain ZIKV in U937 cells, large numbers of naïve U937 cells in the cultures infected by these persistent ZIKVs produced by U937 cells in the first round of ZIKV persistence quickly became IFA-positive for ZIKV antigen, developed prominent CPE changes and underwent cytolytic necrosis. Only a small number of cells could continue to survive and grow in the ZIKV-infected U937 cell culture. In similar instance, the PRV and FLR strains of ZIKV are highly infectious and cause cytopathic effects in HEK293 cells. Only the few of the HEK293 cells that could quickly adapt metabolically to accommodate viral persistence following ZIKV infection could continue to survive in the ZIKV-infected cultures. Therefore, the process of both developing the second round MR 766 strain of ZIKV persistence in U937 cells and establishing persistently PRV and FLR strains ZIKV-infected HEK293 cells appeared to mainly involve selection of the very few members of human blood or epithelial cells with adaptive metabolic functions enabling the cells to continuously survive and grow without actively undergoing apoptosis or cytolysis, despite active intra-cellular propagation of ZIKVs. The comparative genomic study of the persistent ZIKVs produced by the persistently MR 766 strain ZIKV-infected U937 cells established in the first round of ZIKV persistence without having detectable CPE and cytopathogenicity and the second round of ZIKV persistence with prominent CPE and cytopathogenicity revealed positive or negative of stable nucleotide variations or mutations in the key genes compared to the respective inoculum ZIKVs [
10].
The comparative genome sequence analyses of persistent ZIKVs produced by the persistently PRV and FLR ZIKV strains ZIKV-infected HEK293 cells and the original inoculum of ZIKV or ZIKVs produced during the acute infection of HEK293 cells showed that the genomes of both persistent PRV and FLR ZIKVs were genetically more diverse and heterogenous with more nucleotide variations in different nucleotide positions of different genes (
Table 1 and
Table 2). In the prolonged process of establishing persistently ZIKV-infected cultures, the populations of ZIKVs would gradually become more heterogenous without a definite selection pressure on the infecting inoculum viruses. Importantly, our genomic analyses revealed that there were apparently different processes of viral selection in the intricate interplays of between infecting PRV and FLR ZIKV strains with the HEK293 host cells. The genomes of persistent ZIKVs, Zp_H, produced by the persistently PRV strain ZIKV-infected HEK293 cell line developed after an initial fulminant cytopathic ZIKV infection did not show any consistent nucleotide changes or stable variations. In this context, a recent study of ZIKV genomes in the persistently ZIKV-infected culture of human fetal astrocytes also reported no high degree of stable variation found [
19]. In contrast, the genomes of persistent ZIKVs, Zf_H, produced by the persistently FLR strain ZIKV-infected HEK293 cell line developed after highly cytopathic ZIKV infection had stable variations at several positions in various NS genes and one in precursor protein gene (Red shallow in
Table 2). Specific clones of FLR strain ZIKV with the mutations or stable variations in these genes were evidently selected in the process of developing viral persistence in the ZIKV-infected HEK293 cells. Further studies are needed to elucidate how these nucleotide variations could affect the cell infectivity and cytopathogenicity of the persistent ZIKV, Zf_H, on Vero cells and other human epithelial cells. The findings of fixed patterns of mutations or at times lack of fixed patterns of mutations in genomes of the 2 persistent ZIKVs produced by the persistently FLR and PRV strains ZIKV-infected HEK293 cells showed there is more than one way to establish viral persistence in the ZIKV-infected human host cells.
The present study and our previous study [
10] revealed that both low grade with slow rate ZIKV infections that produced no detectable CPE and high grade with rapid rate ZIKV infections that produced prominent CPE as well as extensive cell necrosis could lead to ZIKV persistence. However, the mechanisms of developing viral persistence for the ZIKVs in infected human cells could be very different. Both viral strains/isolates and cellular mechanisms of infected host cells play an important role in the development of ZIKV persistence in human cells of different developmental characteristics. The processes and the outcomes of establishing ZIKV persistence are highly complex. Our studies revealed that infections by ZIKVs with low infectivity and cytopathogenicity could lead to selection of more virulent ZIKVs, some with even altered cell infection specificity, produced by the persistently ZIKV-infected cells. On the other hand, infections by ZIKV with high infectivity and cytopathogenicity in human epithelial cells may also lead to selection of less virulent viruses produced by the persistently ZIKV-infected cells.
The possible clinical relevance of the study results includes the apparent new challenge in detection of the persistent ZIKVs with genomic sequence variations in the persistently ZIKV-infected cells in tissues. In addition, the finding that infections of HEK293 cells with neuronal differentiation properties by the two American isolates of ZIKV, but not the prototype ZIKV MR766 strain, could result in viral persistence in the cells may also have its clinical relevance [
12]. The two seemingly less pathogenic American isolates of ZIKV that can establish persistence in the human cells with neuronal developmental characteristics may be playing a previously unappreciated role in the recent ZIKA virus epidemic associated with fetal microcephaly of congenital ZIKV syndrome [
12]. Our on-going comparative gene expression study for neuronal differentiation and development in HEK293 cells with or without persistent ZIKV infections may shed some lights on the relevance of ZIKV-related neurological abnormalities in recent ZIKV epidemic. In this context, it is also unclear how PRV and FLR isolates with similar genetic background showed two dissimilar patterns of genomic changes during the persistent infection. Further study will also be needed to verify if the dissimilar patterns found are indeed strain-specific. Evidently, the cell culture study will likely be over simplistic in comparison with the complexity of various clinical conditions. However, the cell culture study could yield important and useful information that may not be easily attenable clinically. We anticipate that the study will further contribute to the understanding of the fundamental biology of adaptive mutations and selection for both the infecting viruses and the infected human host cells during development of ZIKV persistence. The persistently ZIKV-infected human cell lines that we have developed should also be useful for investigating critical molecular pathways of ZIKV persistence and to study drugs or countermeasures against ZIKV infections and transmission.
4. Materials and Methods
4.1. Virus and Cells
Zika virus stocks, PRVABC59 strain (Puerto Rico strain, NR-50244) and FLR strain (Colombia strain, NR-50241) were obtained from BEI Resources, NIAID/NIH (ATCC, Manassas, VA, USA). Cell lines used in this study include: human embryonic kidney cells, HEK293 (ATCC CRL-1573), African green monkey kidney cells, Vero (ATCC CCL-81). Cell lines were purchased from ATCC (Manassas, VA, USA). Vero cells were maintained in RPMI 1640 medium containing L-glutamine and 25 mM HEPES (Corning, Corning, NY, USA) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin solution (P/S) at 37 °C with 5% CO2; all the other cell lines were grown in Minimum Essential Medium (DMEM, Corning) supplemented with 10% FBS and 1% P/S solution.
4.2. Propagation of the ZIKV PRV and ZIKV FLR Working Stocks
The working Zika virus stocks were prepared as we reported previously [
10]. Vero cells were cultured in a T-75 culture flask to 90% confluent. After aspirating medium from the cell culture, Vero cells were inoculated with ZIKV suspension in 2 mL of RPMI 1640 medium containing 2% FBS and 1% P/S (R2 medium). Viral inoculums were prepared from ZIKV stocks obtained from BEI Resources at multiplicity of infection (MOI) of 0.1 ± 0.01 according to the package description. The infected Vero cell cultures were incubated at 37 °C for 1 h with gentle rocking very 15 min. After viral absorption, 10 mL of R2 medium was added to the cell culture, the ZIKV-infected Vero cell cultures were incubated in a CO
2 incubator at 37 °C and monitored daily for the appearance of viral CPE. When ~80% of cells detached from the culture surface of the flask after developing CPE, the culture supernatants were harvested by centrifuging for 10 min at 1000×
g. The culture supernatants were passed through a 0.22 µm filter to remove cell debris, aliquoted, tittered and stored at −80 °C as the ZIKV working stocks to be used in the subsequent experiments.
4.3. Infection of HEK293 Cell Line with Zika Virus
HEK293 cells were seeded in a T-75 tissue culture flask, cells grew to full confluence, which contained approximately 2–3 × 107 cells per flask. To infect HEK293 cells, 3 mL of R2 medium containing ~3 × 106 p.f.u. of ZIKV PRV or ZIKV FLR working stock was added to each flask, at MOI of about 0.1, and incubated at 37 °C for 2 h with frequent rocking. After incubation, cells were washed with PBS, and 30 mL of DMEM culture medium supplemented with 10% FBS was added to the cell culture. The infected cell cultures were maintained at 37 °C with 5% CO2. Cell cultures were monitored daily to observe any cell morphology changes and CPE. To harvest acute-infected virus produced in HEK293 cells, culture supernatants were collected by centrifugation 4–5 days after infection. Culture supernatants were stored at −80 °C for viral titration and viral genome copy estimation.
To establish persistently ZIKV-infected cell cultures, fresh culture medium was replenished to the ZIKV-infected HEK293 cell cultures after degenerated cells detached from culture surface were removed. The cell cultures were continuously maintained until surviving cells re-populated the cultures and the cultures could be passed continuously.
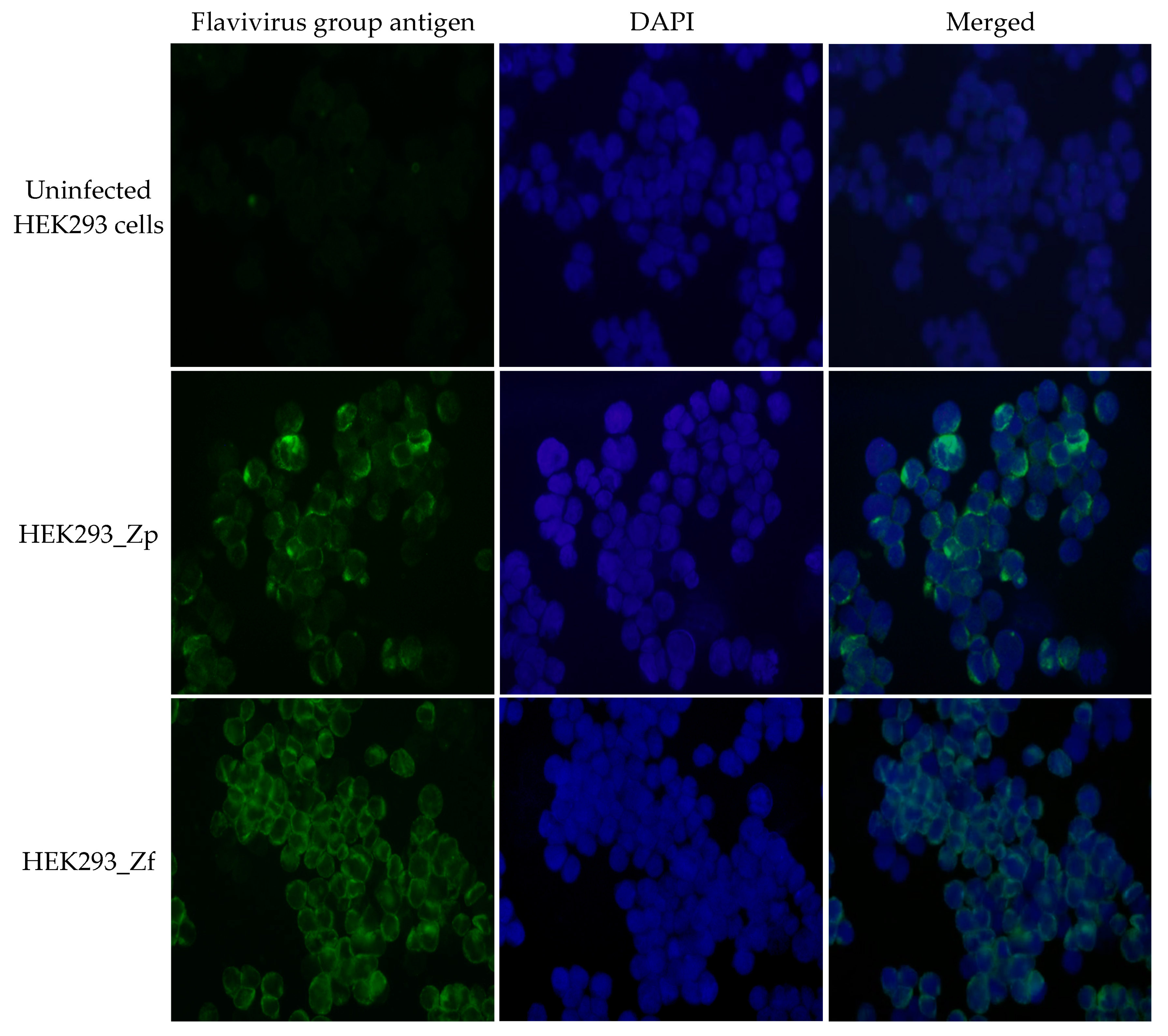
4.4. Immunofluorescence Assay (IFA)
To prepare cell samples for immunofluorescence assay (IFA), cell culture was trypsinized, cell suspension was centrifuged (1000×
g, 10 min), and cell pellet was washed once with phosphate buffered saline (PBS). After centrifugation, the washed cell pellet was re-suspended in 10–20 µL of PBS and cell suspension was dotted on a microscopic slide. The slides were air-dried and then fixed with a mixture of methanol: acetone (1:1) for 5–10 min at room temperature. The procedure for IFA detection of ZIKV-specific antigen has been previously described [
10]. Anti-Flavivirus Group antigen monoclonal antibody (Clone D1-4G2-4-15, BEI Resources) was used as the primary antibody for IFA to detect cells that were positive for producing ZIKV antigen, in order to detect the presence of ZIKV antigenic proteins in cells. Cell dots were blocked for 30–60 min with a blocking solution (KPL, Gaithersburg, MD, USA), and 1% bovine serum albumin (BSA) in PBS containing 100 µg/mL of human immunoglobulin. After removing the remaining liquid, cell dots were then incubated with primary antibody (1:300 dilutions in 1% BSA-PBS) at room temperature for 30–60 min in a humidified chamber. After washing with PBS 3 times, the cell smears were incubated with a secondary antibody, Alexa Fluor 488 conjugated goat-anti-mouse IgG (1:400) (Jackson ImmunoResearch Labs, West Grove, PA, USA), for 15–20 min. The slide was again washed with PBS 3 times and then once with distilled water. The slides were air dried and mounted with glycerol-PBS mixture containing DAPI (4′,6-diamidino-2-phenylindole, Sigma, St. Louis, Mo, USA). The cell images were captured using an up-right immunofluorescent microscope with attached digital camera system (Olympus BX51, Olympus, Center Valley, PA, USA).
4.5. Assay of Apoptosis
Apoptosis of cells was detected using TUNEL (TdT-mediated dUTP-biotin nick end-labeling) method [
13]. Cell cultures were seeded on a circular cover-glass in a 12-well tissue culture plate and cultured for 3–4 days. Cell slides were washed twice with PBS and fixed with 3.7% formaldehyde in PBS for 2 h at room temperature. Fixed cells were washed with PBS and permeabilized with 0.1% Triton X-100 PBS containing 1% BSA and 50 mM glycine for 30 min at room temperature. To stain the apoptotic cells, an ApopTag Peroxidase In Situ Apoptosis Detection Kit (MilliPore, Burlington, MA, USA) was used following the manufacturer’s suggested protocols. Cell slides were incubated with a True Blue peroxidase substrate solution (KPL) to visualize the apoptotic cells. The stained cell slides were imaged using a research microscope with digital camera system (Olympus BX51, Olympus).
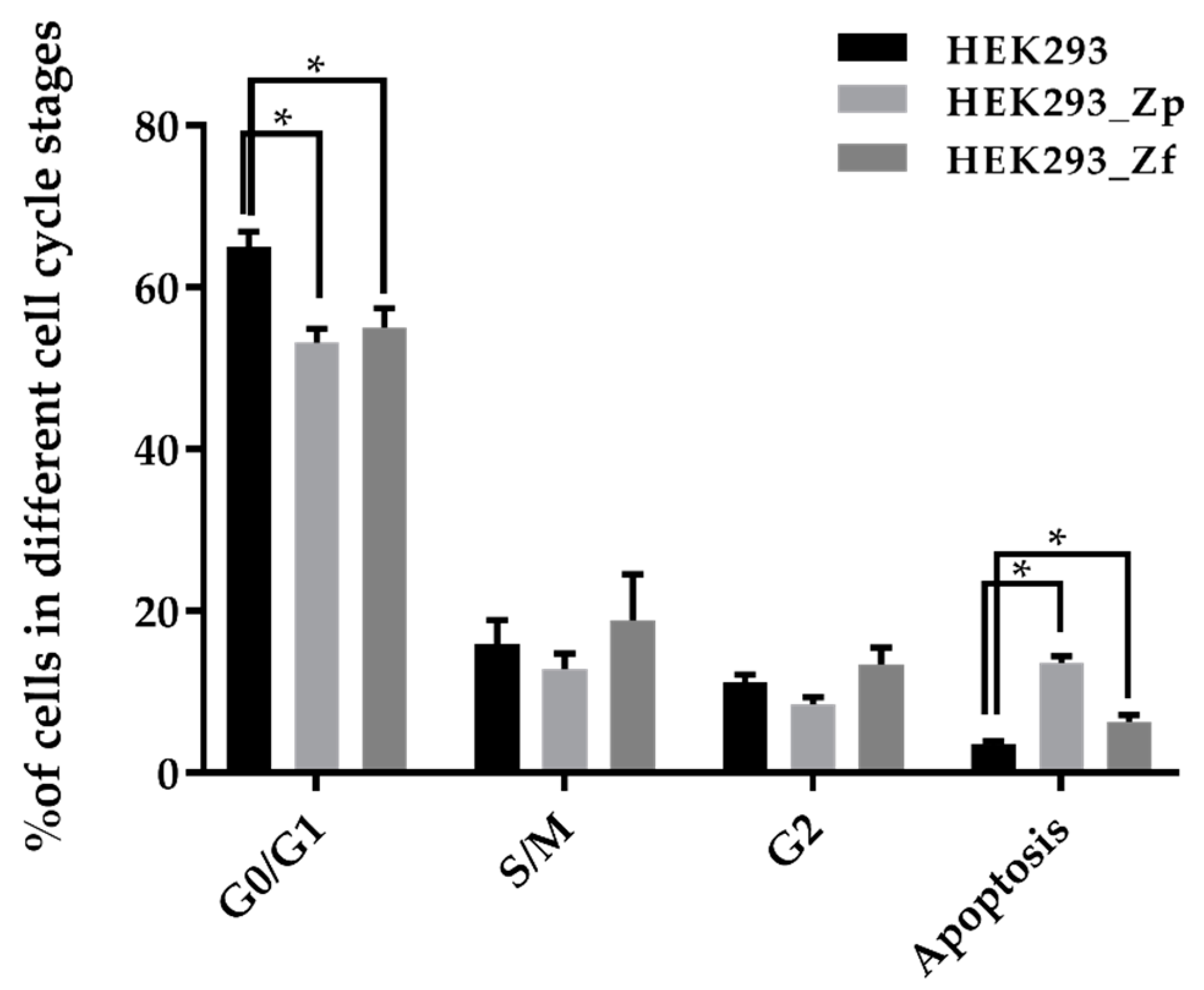
4.6. Cell Cycle Analysis
To perform cell cycle analysis, HEK293 cells (infected or non-infected) were seeded in T-25 cell culture flasks at 1–2 × 106 cells per flask and incubated at 37 °C for 3 days. The total cells in each flask were harvested by trypsinization, cell pellets were washed with PBS and then fixed with cold (−20 °C) 70% ethanol by adding ethanol drop by drop into each cell pellet while gently vortexing. The fixed cell suspensions were stored in a −20 °C freezer. To conduct cell cycle analysis, the fixed cell pellets were washed first with PBS, and then with staining buffer (2% FBS, 0.1% NaN3 in 1 × PBS) by centrifuging for 10 min at 1500 ×g and aspirating the supernatant. Approximately 1 × 106 cells were re-suspended in 0.1 mL of staining buffer, 20 μL 7-AAD (BD, Franklin Lakes, NJ, USA) was added into each tube and incubated for 15 min in the dark at room temperature. Cell cycle analysis was performed using a flow cytometer (BD LSRFortessa X-20, BD). Assay was performed in triplicate.
4.7. Viral Production Kinetics of Persistently ZIKV-Infected HEK293 Cells
To study ZIKV production kinetics of persistently infected HEK293 cells, infected cells were seeded in 12-well tissue culture plates at 2 × 105 cells/well in 1 mL of culture medium and incubated at 37 °C with 5% CO2. Culture supernatants (0.4 mL) in 3 culture wells of each cell line were collected daily for viral titration and viral genome copy estimation. After supernatant collection, total cells in each well were harvested by trypsinization, and viable cells in each culture well were enumerated using a hemocytometer. Triplicate wells were used for each experiment. The experiment was performed for 5 days.
4.8. Endpoint Dilution Assay (TCID50)
The ZIKV infectious titer of culture supernatant was estimated as the 50% tissue culture infective dose (TCID
50), using Vero cells as the assay target. The procedure has been reported in our previous study [
10]. Exponentially growing Vero cells were seeded in 96-well culture plates at 2 × 10
4 cells/well in RPMI 1640 medium containing 10% FBS one day before performing the assay. Aliquots of cell culture supernatants to be titrated were 10-fold serially diluted in R2 medium. To perform the assay, the culture medium of the prepared 96-well plates of Vero cells was aspirated and 20 µL of serially diluted culture supernatant to be tested was rapidly added into each well. A total of 8 wells were inoculated for each serial dilution. After incubating the plates at 37 °C for 2 h for viral absorption with frequent rocking every 15–30 min, 150 µL of fresh R2 medium was added into each well. The plates were incubated in a CO
2 incubator at 37 °C and monitored for ~5 days. At the end of the assay, the numbers of culture wells for each dilution point showing clear viral CPE were recorded. The TCID
50 titer was calculated based on the method described by Hierholzer and Kilington in the first edition of the Virology Methods Manual. Assay was conducted in triplicate.
4.9. Real-Time One-Step Reverse Transcription-Quantitative PCR (RT-qPCR)
Quantification of v-RNA of ZIKV genome was performed by a real-time RT-PCR assay using a ZIKV-specific primer set suitable for detecting both PRV and FLR ZIKV strains. The assay procedure has been previously reported [
10]. Total RNA of each culture supernatant was extracted from 140 µL of sample stored at −80 °C using QIAamp Viral RNA Mini Kit (Qiagen, Dusseldorf, Germany), following the manufacturer’s protocol. The v-RNA copy numbers in purified RNA were quantified using iTaq Universal SYBR Green One-Step Kit (Bio-Rad, Hercules, CA, USA) and Bio-Rad CFX96 system (Bio-Rad). Each reaction mixture of one-step RT-qPCR contained 5 µL of purified RNA, 10 µL of iTaq universal SYBR Green reaction mix, 0.25 µL of iScript reverse transcriptase, 0.5 µL each of 10 µM forward and reverse primers, and 4.25 µL of nuclease-free water, to a final volume of 20 µL. The RT-qPCR was started from 50 °C reverse transcription step for 30 min and followed by PCR amplification with 95 °C pre-heat for 1 min and then 45 two-step thermocycles at 95 °C for 10 s and 57 °C for 30 s. The ZIKV-specific primer pair used in the RT-qPCR reaction was described previously [
20]. The sequences of the forward and reverse primers are: 5′-CCTTGGATTCTTGAACGAGGA-3′ and 5′-AGAGCTTCATTCTCCAGATCAA-3′. The standard curve for estimating v-RNA copy number present in the tested sample was calculated by fitting the Ct value to a strand curve. The standard curve was generated by RT-qPCR run against 5 RNA samples prepared from a 10-fold serial dilution of the ZIKV PRV genomic RNA solution containing 2.0 × 10
4 ZIKV RNA genome copies/µl (or an RNA solution containing 4.4 × 10
4 ZIKV RNA genome copies/µL for ZIKV FLR) provided by BEI. Quantification was performed in triplicate.
4.10. Plaque Assay
Approximately 5 × 104 Vero cells were seeded into 24-well plates 24 h before the experiment within 1 mL of RPMI 1640 medium containing 10% FBS. Supernatants from ZIKV-infected cell cultures were diluted with R2 medium to achieve equal copy number of ZIKV v-RNA in the same volume. The culture medium of the 24-well plates of Vero cells was aspirated and 200 µL of the prepared culture supernatants were added into each well (in duplicate). After incubating the plates at 37 °C for 2 h for viral absorption (with gentle rocking every 15 min), the infected cells were washed with PBS after aspirating the inoculums from the wells, 1 mL overlay medium was added into each well. The plates were incubated in a CO2 incubator at 37 °C and monitored for 3–5 days. The overlay medium was prepared by adding 0.5% agarose to RMPI 1640 medium supplemented with 2% FBS and antibiotic solution. After incubation, cells were fixed with 2 mL/well of 3.7% paraformaldehyde in PBS for 2–4 h. The agarose overlay and formaldehyde solution were removed from culture wells. Cells were rinsed with water and stained with either 0.5% crystal violet in 50% ethanol or True Blue (KPL) after immunoblotting with Anti-Flavivirus Group antigen monoclonal antibody (Clone D1-4G2-4-15, BEI Resources) followed by goat anti-mouse IgG H&L (HRP) (ab6789, Abcam, Cambridge, MA, USA).
The images of stained cell culture plates were recorded using a GE imaging system (Amersham Imager 6000, GE, Pittsburgh, PA, USA).
4.11. Infectivity of Zika Virus Produced from Persistently Infected Cells on Vero Cell Lines
Vero cells were seeded in 12-well plates at 1 × 105 cells/well in 1 mL of culture medium one day prior to conducting the experiment. At Day 0, culture medium in each culture well was aspirated, and 400 µL of various ZIKV suspensions in R2 medium was added to each well at an MOI at 0.1. After incubating the plates at 37 °C for 2 h for viral absorption (with frequent rocking), viral inoculums in each culture well were aspirated. The infected cells were rinsed with PBS, and 2 mL of fresh culture medium was added into each well. The plates were incubated in a CO2 incubator at 37 °C. Cell cultures were monitored for morphological changes daily, and 0.5 mL of culture supernatant from each well was collected for viral titration and estimation of viral genome copies. The culture plates were replenished daily with 0.5 mL of fresh culture medium after sample collection to maintain the culture volume of each well at 2 mL. A total of 5 sample of culture supernatants were collected for each well and stored at −80 °C.
4.12. Viral RNA Preparation for Viral Genome Sequencing
To prepare viral RNA for genome sequencing, we followed a previously described procedure [
10]. RNAs of viral particles were extracted from the cell culture supernatant. Twenty mL of culture supernatant from acutely or persistently infected cells was harvested. Each supernatant was centrifuged at 2000×
g for 10 min to remove cell debris and passed through a 0.22 µm filter to further remove cellular residuals. The supernatant was chilled on ice and 10 mL of 24% PEG (polyethylene glycol)-8000 solution (with 2.5M NaCl) was added to the culture supernatant. The PEG-supernatant mixture was rocked at 4 °C, overnight. The mixture was then centrifuged at 3000×
g at 4 °C for 30 min. The precipitate was re-suspended in 100 µL of PBS after the supernatant was discarded. Trizol Reagent, 300 µL, was added to the viral suspension to extract viral RNA using Direct-zol MiniPrep kit (Zymo Research, Irvine, CA, USA), following the procedures provided by the manufacturer. The extracted RNA samples were quality checked and quantified by RNA pico chips of 2100 Bioanalyzer (Agilent, Santa Clara, CA, USA).
4.13. RNA-Seq of Viral Genome
RNA-Seq of viral genomes was studied using an Illumina Miseq platform following the previously reported procedure [
10]. The viral RNAs from various culture supernatants were used to generate libraries using an Illumina TruSeq stranded total RNA sample prep kit, following the manufacturer’s standard procedures. In brief, total RNA was fragmented in the presence of divalent ions at 94 °C for 8 min. Fragmented RNAs were reverse-transcribed into first-strand cDNAs, and then second-strand cDNAs. The double strand cDNAs were adenylated at the 3′-ends, and then ligated with sequencing index adaptors. After amplifying for 15 cycles. The cDNA Libraries were loaded in one MiSeq Nano Flow Cell using MiSeq Reagent Nano kit for a 100-cycle paired-end sequencing on the MiSeq sequencer (Illumina, San Diego, CA, USA).
4.14. Analysis of the Viral Genome Variants
To study viral genomes and variation, a previously reported procedure was followed [
10]. The fastq files generated from the Illumina MiSeq sequencer were used for viral genome variant analysis. All the sequence analyses were performed using the software package of CLC Genomics Workbench version 11.0.1. The raw reads were trimmed to remove the regions with more than two ambiguous bases and regions having 5% of the bases with quality score lower than 20. The trimmed raw reads were then mapped to the reference sequence of the ZIKV PRVABC59 (Puerto Rico strain) (GenBank Accession number: KX601168) or the ZIKV FLR (Colombia strain) (GenBank Accession number: KX087102) with the mapping parameter: match score = 1, Mismatch cost = 2, insertion cost = 3, deletion cost = 3, mapped length fraction > 0.5, and similarity fraction > 0.8. The local realignment was applied to realign the unaligned ends from the reads mapping. The aligned reads tracked from the local realignment were subject to Low Frequency Variant Detection workflow with the parameter: Ploidy = 1, required variant probability = 80%, the minimum coverage = 5, minimum reads count = 2, minimum frequency = 1%.
4.15. Statistical Analysis
All continuous variables are presented as mean ± SD. Data among groups were analyzed using ANOVA followed by student-t test. Values of p < 0.05 were considered statistically significant.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}