Insulin Treatment Forces Arteriogenesis in Diabetes Mellitus by Upregulation of the Early Growth Response-1 (Egr-1) Pathway in Mice

 , ,
, , 
Abstract
:1. Introduction
2. Results
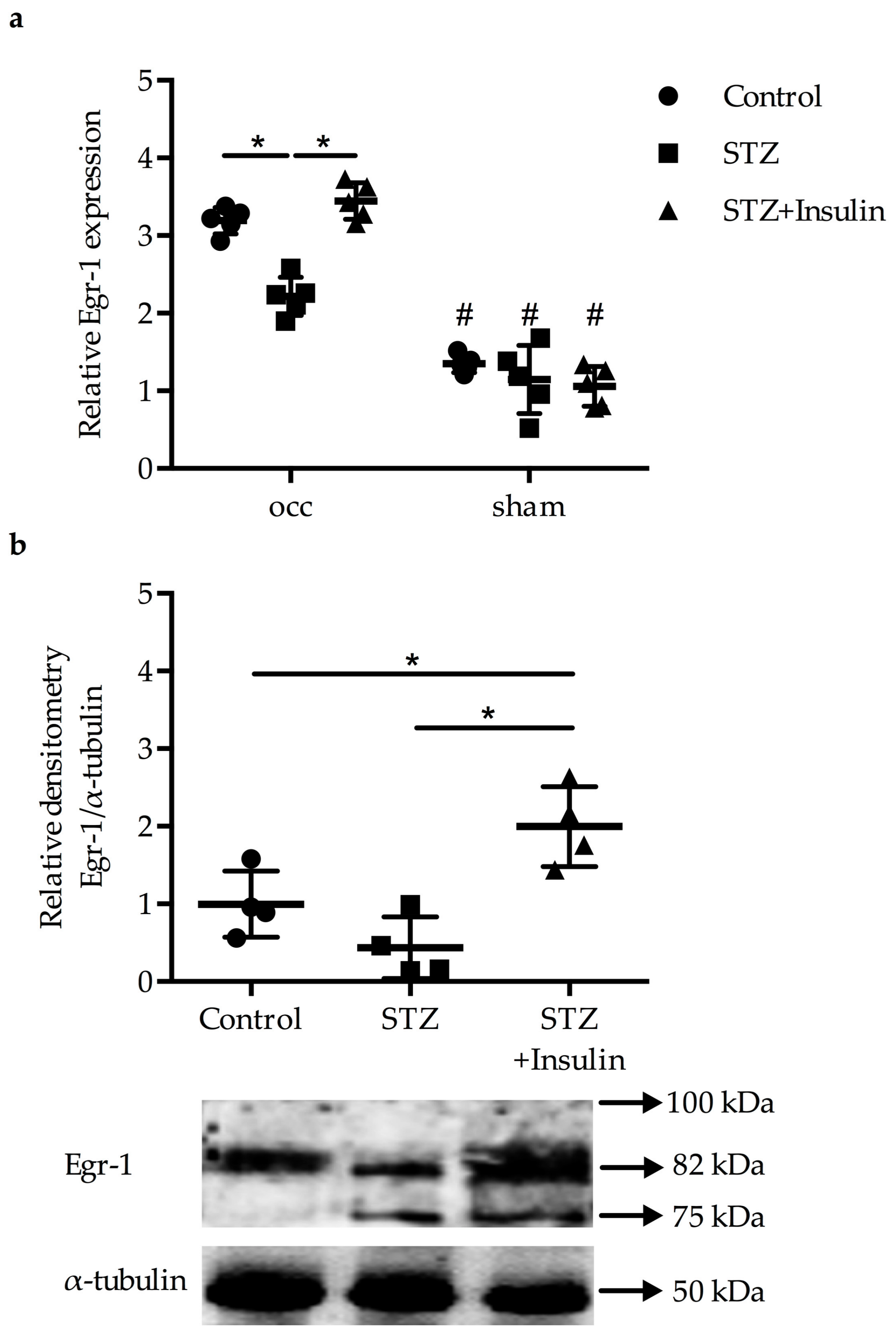
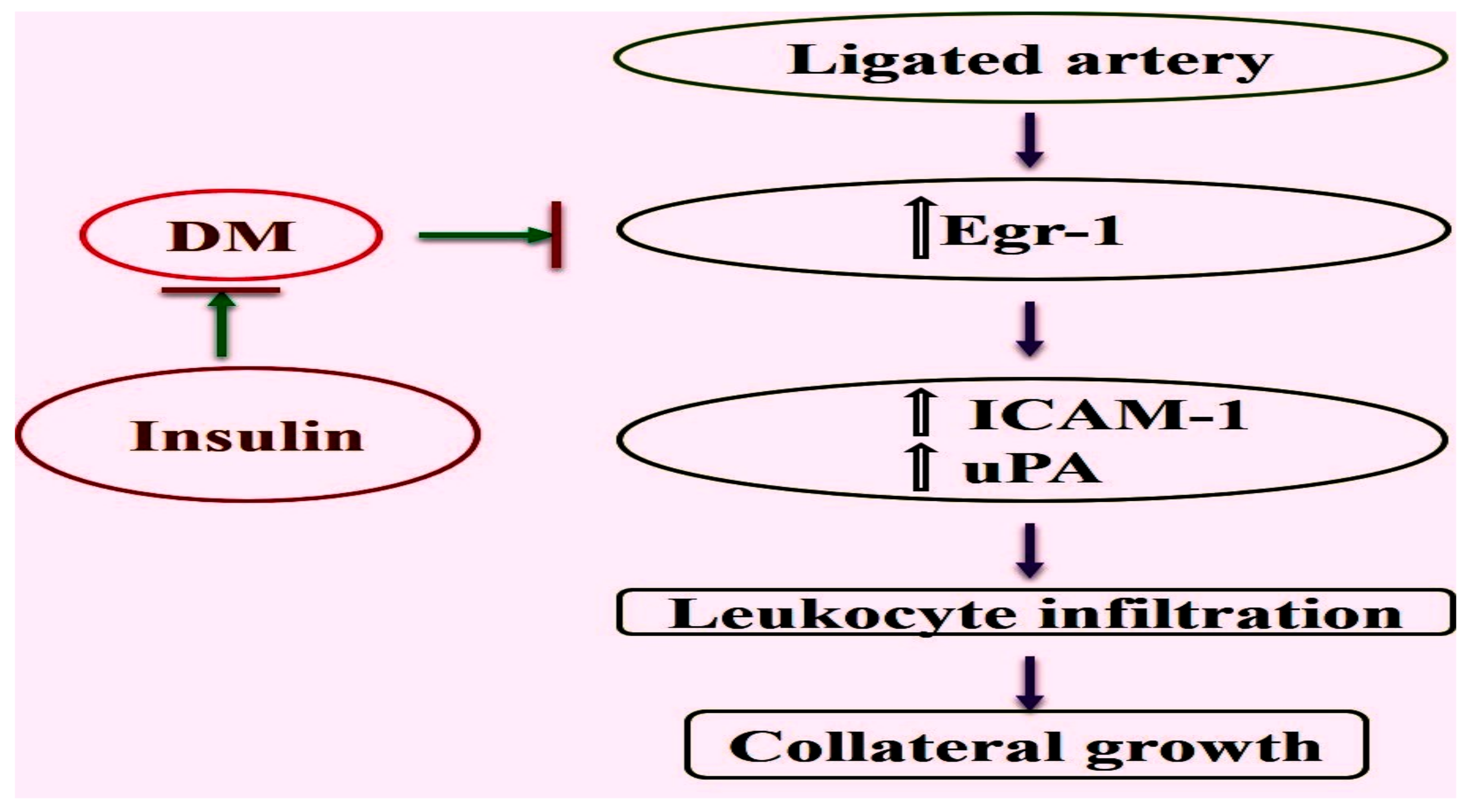
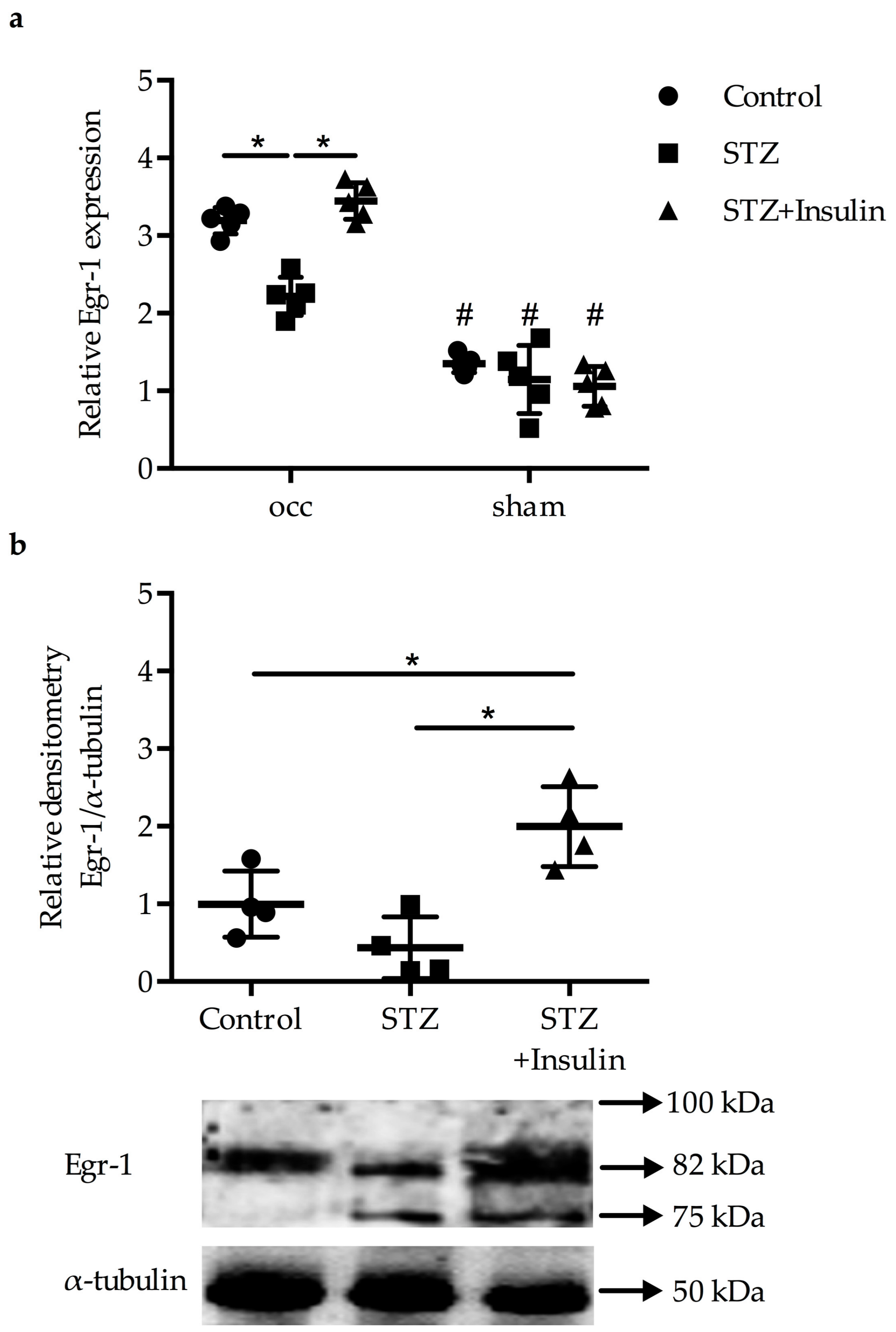
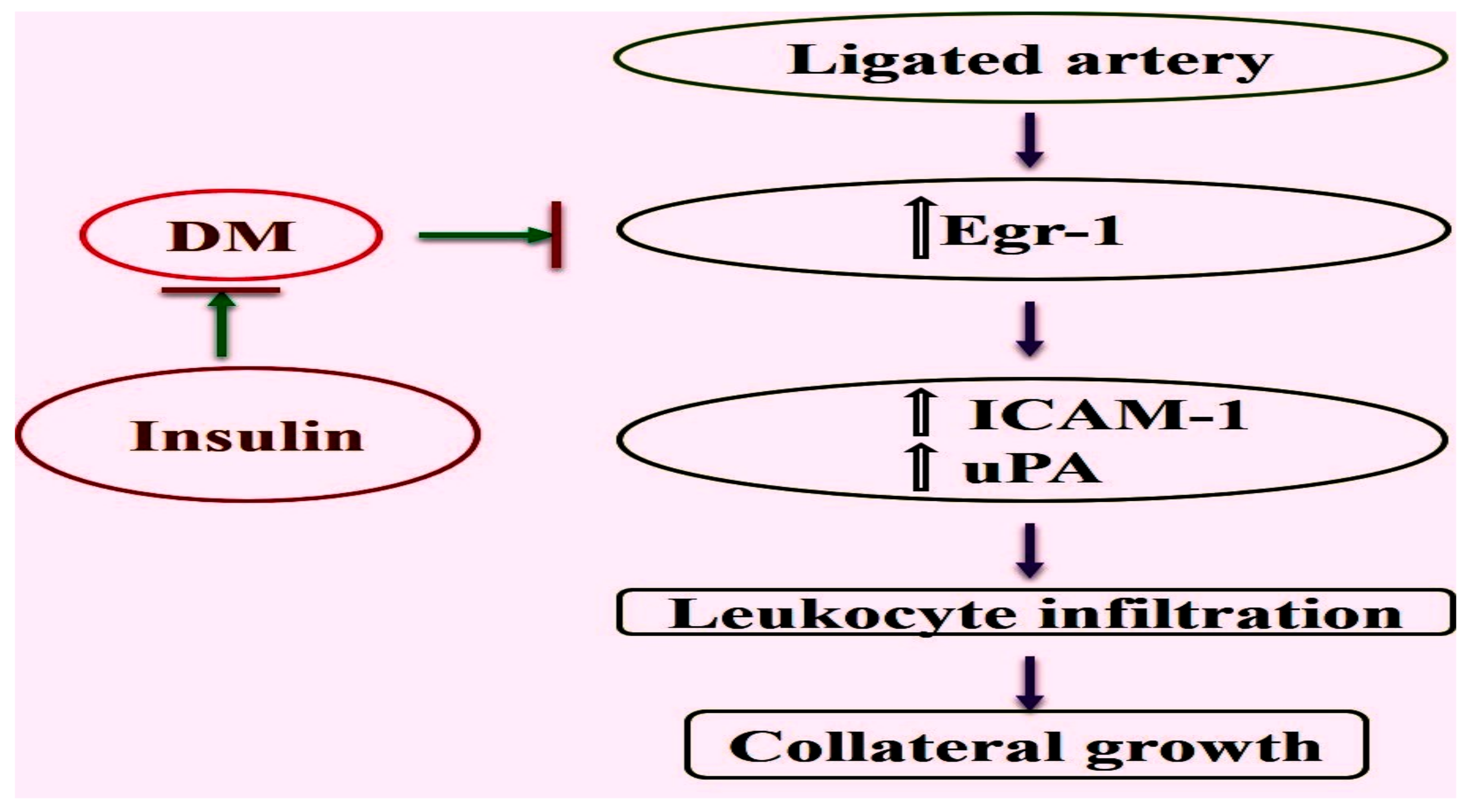
2.1. Reduced Upregulation of Egr-1 in Growing Collaterals of Diabetic Mice
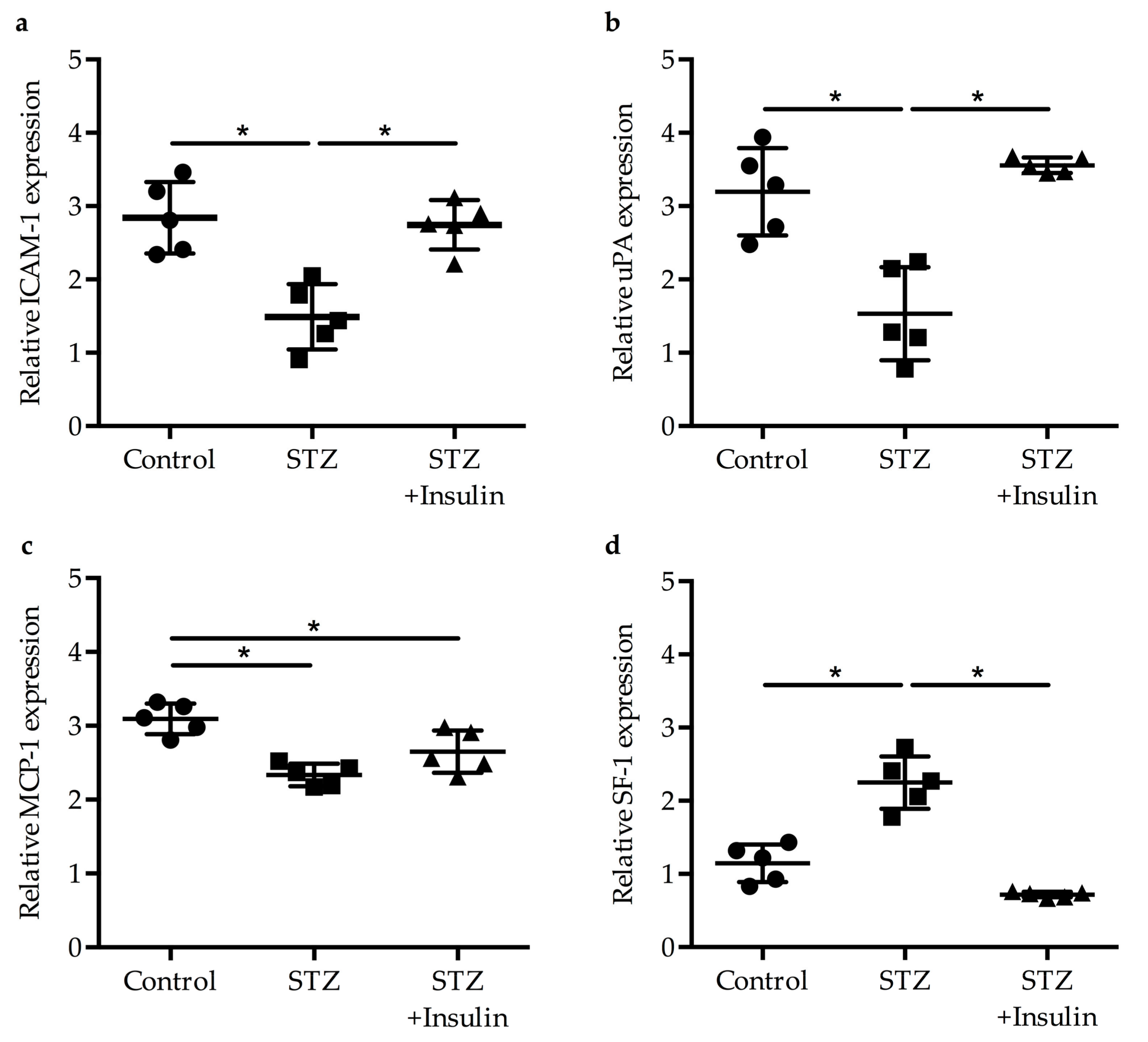
2.2. Expression of Egr-1 Downstream Genes in Collaterals of Diabetic Mice Was Restored by Insulin Treatment
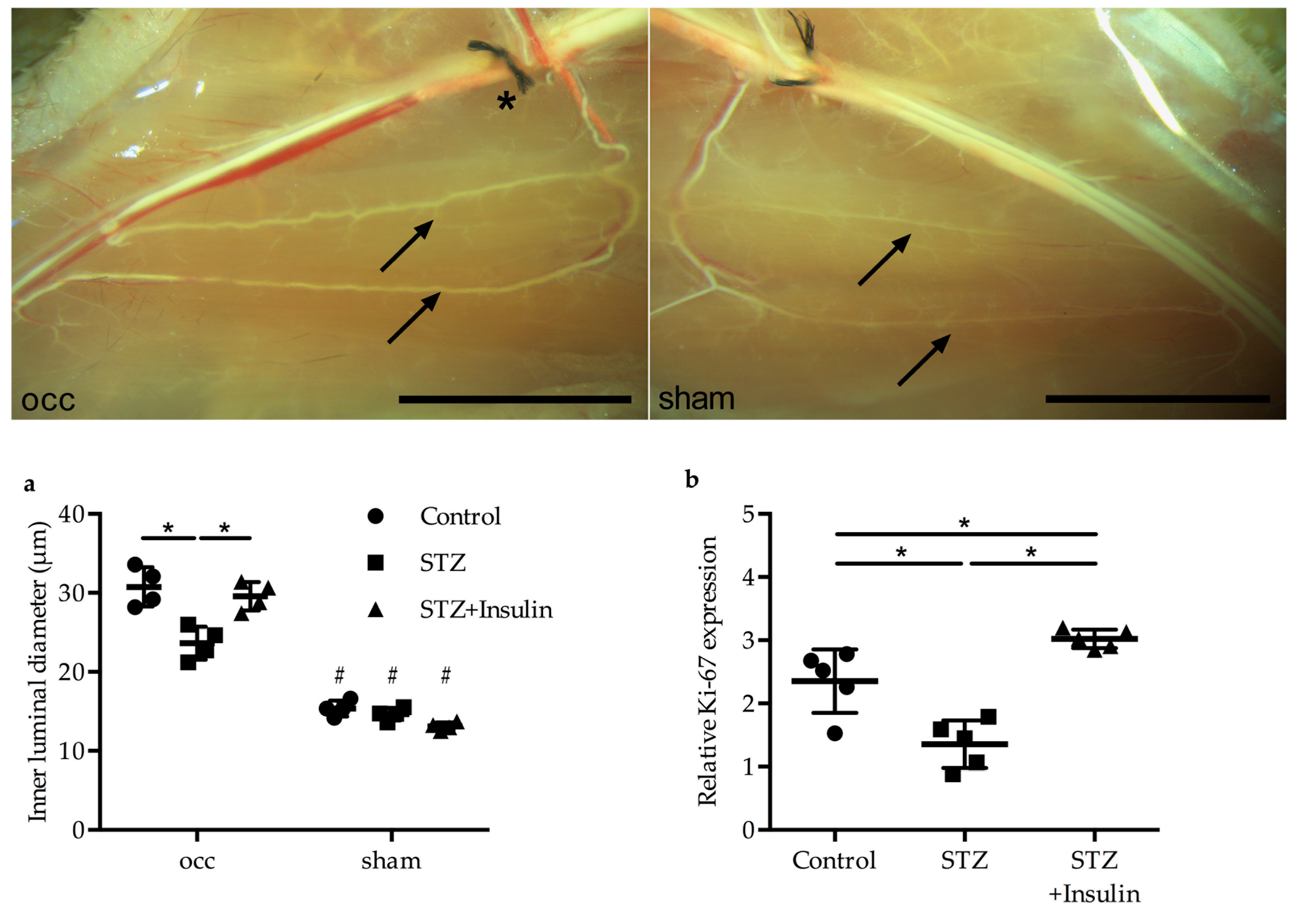
2.3. Insulin Treatment Restored Vessel Growth in Diabetic Mice
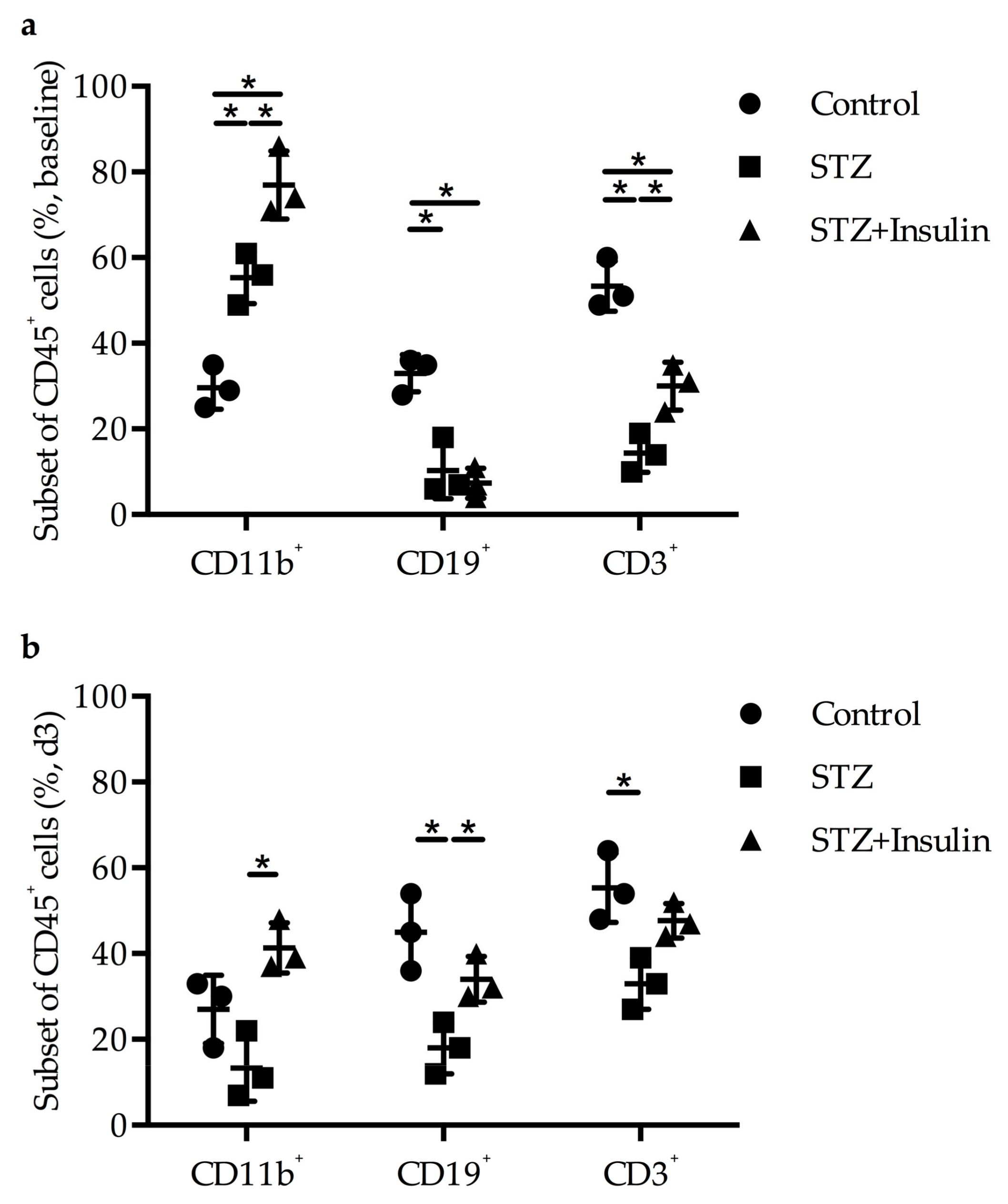
2.4. Diminished Leukocyte Infiltration Was Improved in Diabetic Mice by Insulin Treatment
3. Discussion
4. Materials and Methods
4.1. Animal Studies
4.2. Femoral Artery Ligation and Collection of Collaterals
4.3. RNA Isolation and Quantitative Real-Time PCR Studies
4.4. FACS Analyses of Blood and Muscle Tissue
4.5. Histological Analyses
4.6. Western Blot
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SF-1 | Splicing Factor 1 |
| DM | Diabetes Mellitus |
| NDM | Non-Diabetes Mellitus |
| Egr-1 | Early growth response-1 |
| uPA | urokinase Plasminogen Activator |
| MCP-1 | Monocyte Chemoattractant Protein-1 |
| ICAM-1 | Intercellular Adhesion Molecule-1 |
| FAL | Femoral Artery Ligation |
| STZ | Streptozotocin |
| PAD | Peripheral Artery Disease |
References
- American Diabetes Association. Peripheral Arterial Disease in People with Diabetes. Diabetes Care 2003, 26, 3333–3341. [Google Scholar] [CrossRef] [PubMed]
- Dolan, N.C.; Liu, K.; Criqui, M.H.; Greenland, P.; Guralnik, J.M.; Chan, C.; Schneider, J.R.; Mandapat, A.L.; Martin, G.; McDermott, M.M. Peripheral artery disease, diabetes, and reduced lower extremity functioning. Diabetes Care 2002, 25, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Prompers, L.; Schaper, N.; Apelqvist, J.; Edmonds, M.; Jude, E.; Mauricio, D.; Uccioli, L.; Urbancic, V.; Bakker, K.; Holstein, P.; et al. Prediction of outcome in individuals with diabetic foot ulcers: Focus on the differences between individuals with and without peripheral arterial disease. The EURODIALE Study. Diabetologia 2008, 51, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Santulli, G. Update on peripheral artery disease: Epidemiology and evidence-based facts. Atherosclerosis 2018, 275, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.A.; Creager, M.A.; Libby, P. Diabetes and atherosclerosis: Epidemiology, pathophysiology, and management. JAMA 2002, 287, 2570–2581. [Google Scholar] [CrossRef] [PubMed]
- Lanzer, P. Topographic distribution of peripheral arteriopathy in non-diabetics and type 2 diabetics. Z. Kardiol. 2001, 90, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, R.T.; Coffman, J.D. Cardiovascular morbidity and mortality in peripheral arterial disease. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2004, 4, 209–217. [Google Scholar] [CrossRef]
- Faglia, E.; Clerici, G.; Clerissi, J.; Gabrielli, L.; Losa, S.; Mantero, M.; Caminiti, M.; Curci, V.; Quarantiello, A.; Lupattelli, T.; et al. Long-term prognosis of diabetic patients with critical limb ischemia: A population-based cohort study. Diabetes Care 2009, 32, 822–827. [Google Scholar] [CrossRef]
- Kamalesh, M.; Shen, J. Diabetes and peripheral arterial disease in men: Trends in prevalence, mortality, and effect of concomitant coronary disease. Clin. Cardiol. 2009, 32, 442–446. [Google Scholar] [CrossRef]
- Malmstedt, J.; Leander, K.; Wahlberg, E.; Karlstrom, L.; Alfredsson, L.; Swedenborg, J. Outcome after leg bypass surgery for critical limb ischemia is poor in patients with diabetes: A population-based cohort study. Diabetes Care 2008, 31, 887–892. [Google Scholar] [CrossRef]
- Currie, C.J.; Morgan, C.L.; Peters, J.R. The epidemiology and cost of inpatient care for peripheral vascular disease, infection, neuropathy, and ulceration in diabetes. Diabetes Care 1998, 21, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Moraes, F.; Paye, J.; Mac Gabhann, F.; Zhuang, Z.W.; Zhang, J.; Lanahan, AA.; Simons, M. Endothelial cell-dependent regulation of arteriogenesis. Circ. Res. 2013, 113, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Arras, M.; Ito, W.D.; Scholz, D.; Winkler, B.; Schaper, J.; Schaper, W. Monocyte activation in angiogenesis and collateral growth in the rabbit hindlimb. J. Clin. Invest. 1998, 101, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Chillo, O.; Kleinert, E.C.; Lautz, T.; Lasch, M.; Pagel, J.I.; Heun, Y.; Troidl, K.; Fischer, S.; Caballero-Martinez, A.; Mauer, A.; et al. Perivascular Mast Cells Govern Shear Stress-Induced Arteriogenesis by Orchestrating Leukocyte Function. Cell Rep. 2016, 23, 2197–2207. [Google Scholar] [CrossRef] [PubMed]
- De Vivo, S.; Palmer-Kazen, U.; Kalin, B.; Wahlberg, E. Risk factors for poor collateral development in claudication. Vasc. Endovasc. Surg. 2005, 39, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Van Weel, V.; de Vries, M.; Voshol, P.J.; Verloop, R.E.; Eilers, P.H.; van Hinsbergh, V.W.; van Bockel, J.H.; Quax, P.H. Hypercholesterolemia reduces collateral artery growth more dominantly than hyperglycemia or insulin resistance in mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Lambiase, P.D.; Edwards, R.J.; Anthopoulos, P.; Rahman, S.; Meng, Y.G.; Bucknall, C.A.; Redwood, S.R.; Pearson, J.D.; Marber, M.S. Circulating humoral factors and endothelial progenitor cells in patients with differing coronary collateral support. Circulation 2004, 109, 2986–2992. [Google Scholar] [CrossRef] [PubMed]
- Gashler, A.; Sukhatme, V. Egr-1: Prototype of a zinc finger family of transcription factors. Prog. Nucleic Acid. Res. Mol Biol. 1995, 50, 191–224. [Google Scholar]
- Yan, S.F.; Harja, E.; Andrassy, M.; Fujita, T.; Schmidt, A.M. Protein kinase C beta/early growth response-1 pathway: A key player in ischemia, atherosclerosis, and restenosis. J. Am. Coll. Cardiol. 2006, 48, A47–A55. [Google Scholar] [CrossRef]
- Sarateanu, C.S.; Retuerto, M.A.; Beckmann, J.T.; McGregor, L.; Carbray, J.; Patejunas, G.; Nayak, L.; Milbrandt, J.; Rosengart, T.K. An Egr-1 master switch for arteriogenesis: Studies in Egr-1 homozygous negative and wild-type animals. J. Thorac. Cardiovasc. Surg. 2006, 131, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Lee, J.Y.; Li Kim, K.; Shin, I.S.; Suh, W.; Choi, J.H.; Jeon, E.S.; Byun, J.; Kim, D.K. Adenoviral-mediated delivery of early growth response factor-1 gene increases tissue perfusion in a murine model of hindlimb ischemia. Mol. Ther. 2005, 12, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Pagel, J.I.; Ziegelhoeffer, T.; Heil, M.; Fischer, S.; Fernández, B.; Schaper, W.; Preissner, K.T.; Deindl, E. Role of early growth response 1 in arteriogenesis: Impact on vascular cell proliferation and leukocyte recruitment in vivo. Thromb. Haemost. 2012, 107, 562–574. [Google Scholar] [PubMed]
- McMahon, S.B.; Monroe, J.G. The role of early growth response gene 1 (egr-1) in regulation of the immune response. J. Leukoc. Biol. 1996, 60, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Yan, SF.; Fujita, T.; Lu, J.; Okada, K.; Shan Zou, Y.; Mackman, N.; Pinsky, DJ.; Stern, DM. Egr-1, a master switch coordinating upregulation of divergent gene families underlying ischemic stress. Nat. Med. 2000, 6, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Scholz, D.; Ito, W.; Fleming, I.; Deindl, E.; Sauer, A.; Babiak, A.; Bühler, A.; Wiesnet, M.; Busse, R.; Schaper, J.; et al. Ultrastructure and molecular histology of rabbit hindlimb collateral artery growth (arteriogenesis). Virchows. Arch. 2000, 436, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Hoefer, I.E.; van Royen, N.; Rectenwald, J.E.; Deindl, E.; Hua, J.; Jost, M.; Grundmann, S.; Voskuil, M.; Ozaki, C.K.; et al. Arteriogenesis proceeds via ICAM-1/Mac-1- mediated mechanisms. Circ. Res. 2004, 94, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.N.; Phukan, S.; Harada, S. Differential regulation of early growth response gene-1 expression by insulin and glucose in vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Gousseva, N.; Kugathasan, K.; Chesterman, C.N.; Khachigian, L.M. Early growth response factor-1 mediates insulin-inducible vascular endothelial cell proliferation and regrowth after injury. J. Cell. Biochem. 2001, 81, 523–534. [Google Scholar] [CrossRef]
- Vedantham, S.; Thiagarajan, D.; Ananthakrishnan, R.; Wang, L.; Rosario, R.; Zou, S.; Goldberg, I.J.; Yan, S.F.; Schmidt, A.M.; Ramasamy, R. Aldose Reductase drives hyperacetylation of Egr-1 in hyperglycemia and consequent upregulation of proinflammatory and prothrombotic signals. Diabetes 2014, 63, 761–774. [Google Scholar] [CrossRef]
- Aliperti, V.; Sgueglia, G.; Aniello, F.; Vitale, E.; Fucci, L.; Donizetti, A. Identification, Characterization, and Regulatory Mechanisms of a Novel EGR-1 Splicing Isoform. Int. J. Mol. Sci. 2019, 20, 1548. [Google Scholar] [CrossRef]
- Reichel, C.A.; Uhl, B.; Lerchenberger, M.; Puhr-Westerheide, D.; Rehberg, M.; Liebl, J.; Khandoga, A.; Schmalix, W.; Zahler, S.; Deindl, E.; et al. Urokinase-type plasminogen activator promotes paracellular transmigration of neutrophils via Mac-1, but independently of urokinase-type plasminogen activator receptor. Circulation 2011, 124, 1848–1859. [Google Scholar] [CrossRef] [PubMed]
- Deindl, E.; Ziegelhöffer, T.; Kanse, S.M.; Fernandez, B.; Neubauer, E.; Carmeliet, P.; Preissner, K.T.; Schaper, W. Receptor-independent role of the urokinase-type plasminogen activator during arteriogenesis. FASEB J. 2003, 17, 1174–1176. [Google Scholar] [CrossRef] [PubMed]
- Cattaruzza, M.; Schafer, K.; Hecker, M. Cytokine-induced down-regulation of zfm1/splicing factor-1 promotes smooth muscle cell proliferation. J. Biol. Chem. 2002, 277, 6582–6589. [Google Scholar] [CrossRef]
- Cattaruzza, M.; Nogoy, N.; Wojtowicz, A.; Hecker, M. Zinc finger motif-1 antagonizes PDGF-BB-induced growth and dedifferentiation of vascular smooth muscle cells. FASEB J. 2012, 26, 4864–4875. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Shen, N.; Zhang, M.L.; Pan, F.Y.; Wang, C.; Jia, W.P.; Liu, C.; Gao, Q.; Gao, X.; Xue, B.; et al. Egr-1 decreases adipocyte insulin sensitivity by tilting PI3K/Akt and MAPK signal balance in mice. EMBO J. 2011, 30, 3754–3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawerbaum, A.; White, V. Animal models in diabetes and pregnancy. Endocr. Rev. 2010, 31, 680–701. [Google Scholar] [CrossRef]
- Limbourg, A.; Korff, T.; Napp, L.C.; Schaper, W.; Drexler, H.; Limbourg, F.P. Evaluation of postnatal arteriogenesis and angiogenesis in a mouse model of hind-limb ischemia. Nat. Protoc. 2009, 4, 1737–1746. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Random Plasma Glucose (mg/dL) |
|---|---|
| Control | 144 ± 10 |
| Streptozotocin | 328 ± 38 * |
| Streptozotocin + Insulin | 216 ± 35 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thulasingam, S.; Krishnasamy, S.; Raj C., D.; Lasch, M.; Vedantham, S.; Deindl, E. Insulin Treatment Forces Arteriogenesis in Diabetes Mellitus by Upregulation of the Early Growth Response-1 (Egr-1) Pathway in Mice. Int. J. Mol. Sci. 2019, 20, 3320. https://doi.org/10.3390/ijms20133320
Thulasingam S, Krishnasamy S, Raj C. D, Lasch M, Vedantham S, Deindl E. Insulin Treatment Forces Arteriogenesis in Diabetes Mellitus by Upregulation of the Early Growth Response-1 (Egr-1) Pathway in Mice. International Journal of Molecular Sciences. 2019; 20(13):3320. https://doi.org/10.3390/ijms20133320
Chicago/Turabian StyleThulasingam, Senthilkumar, Sundar Krishnasamy, David Raj C., Manuel Lasch, Srinivasan Vedantham, and Elisabeth Deindl. 2019. "Insulin Treatment Forces Arteriogenesis in Diabetes Mellitus by Upregulation of the Early Growth Response-1 (Egr-1) Pathway in Mice" International Journal of Molecular Sciences 20, no. 13: 3320. https://doi.org/10.3390/ijms20133320