The Effect of the Repression of Oxidative Stress on Tenocyte Differentiation: A Preliminary Study of a Rat Cell Model Using a Novel Differential Tensile Strain Bioreactor

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
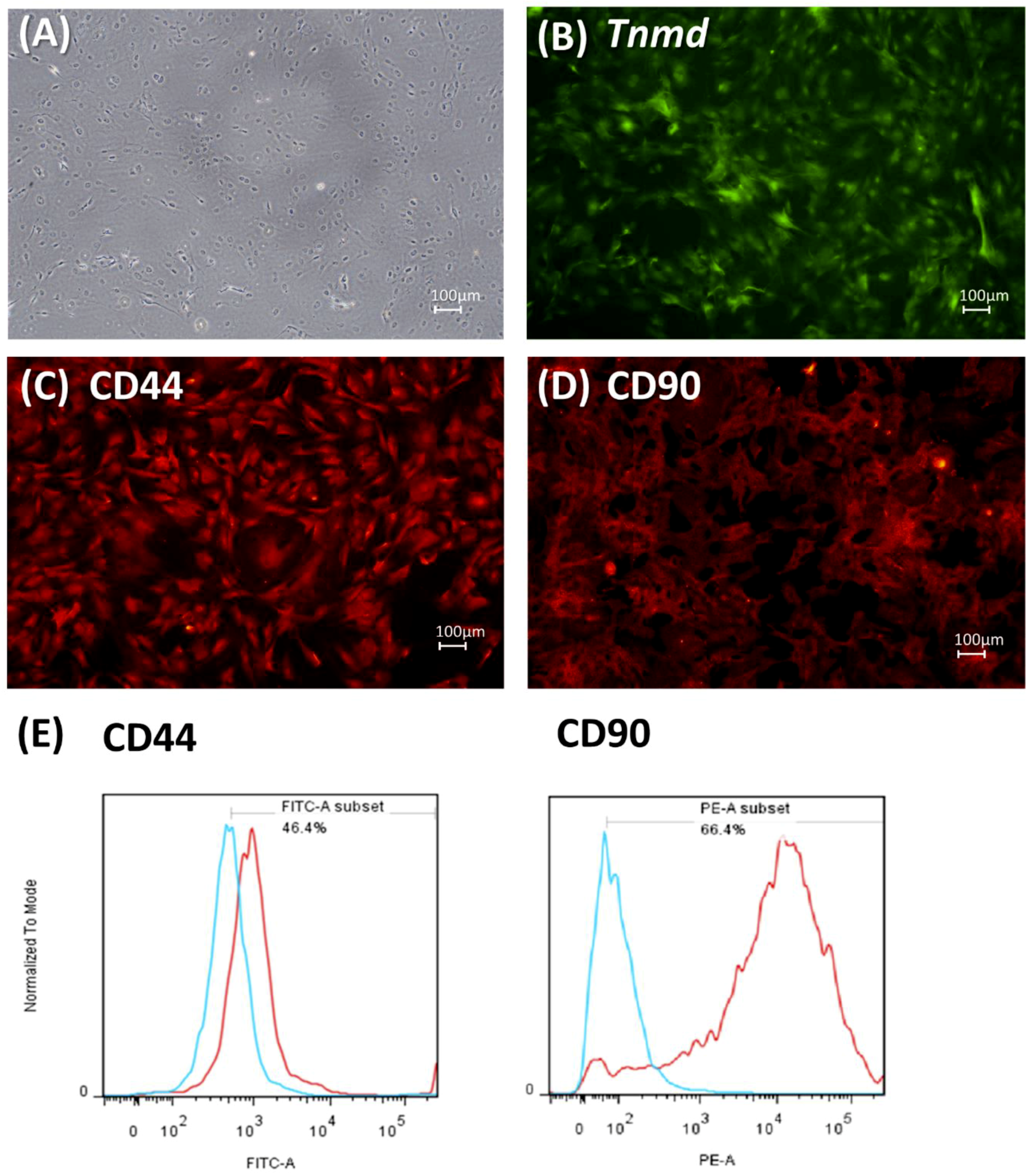
2.1. Characteristics of Tendon-Derived Cells (TDCs)
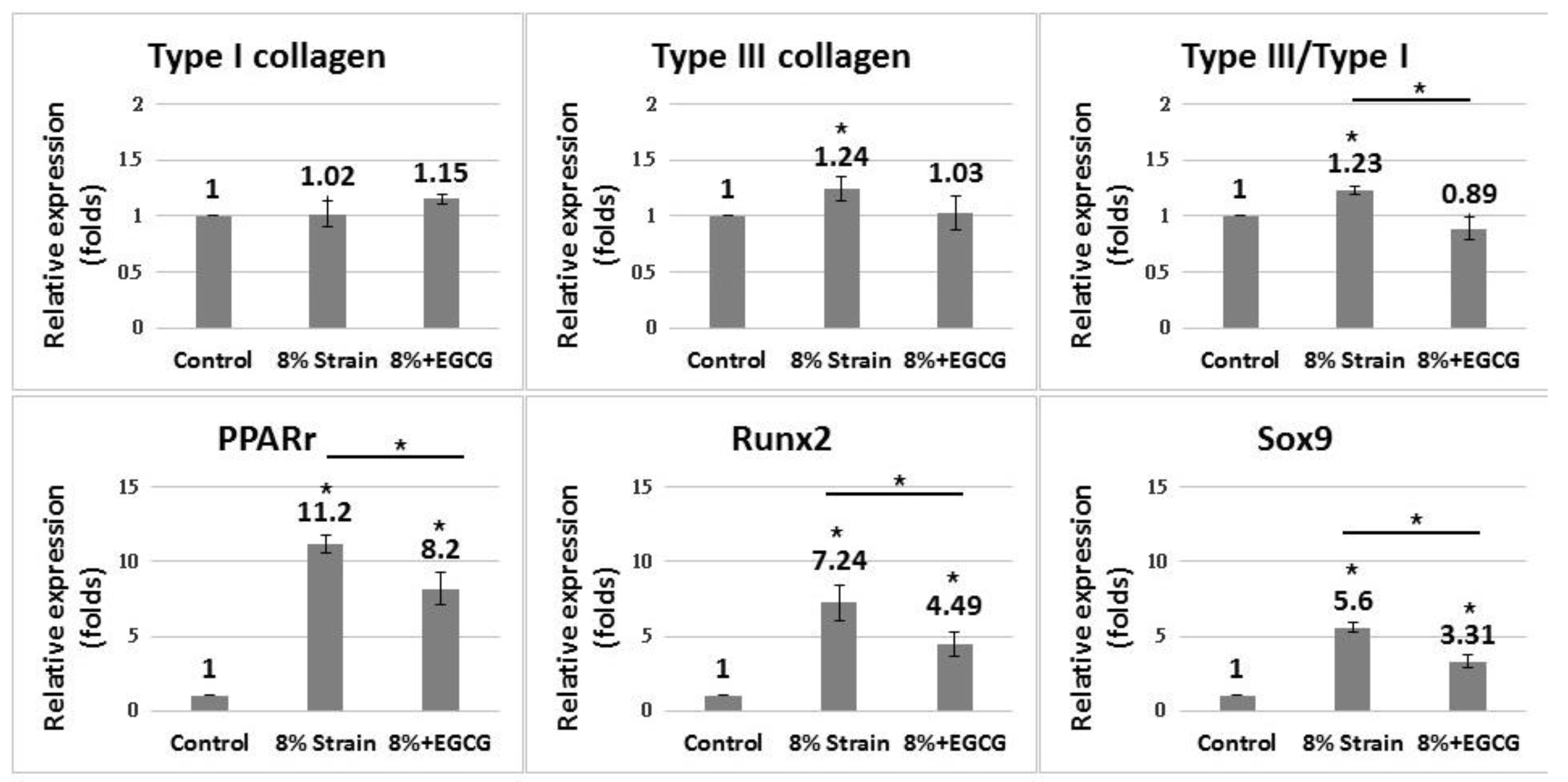
2.2. Effect of Cyclic Uniaxial Stretching on the Gene Expression of TDCs
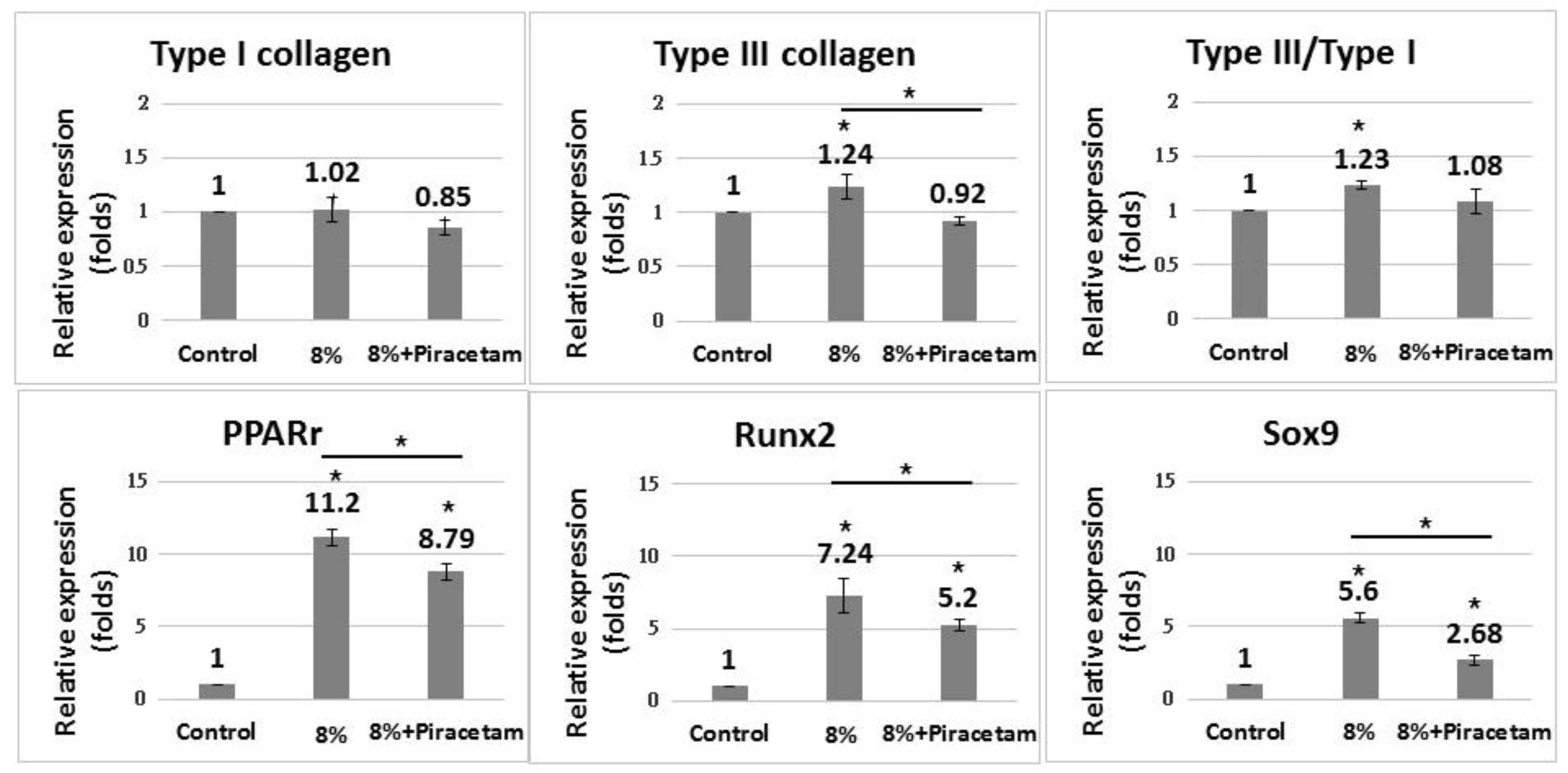
2.3. Effect of EGCG and Piracetam on the Gene Expression of TDCs Subjected to 8% Uniaxial Cyclic Stretching
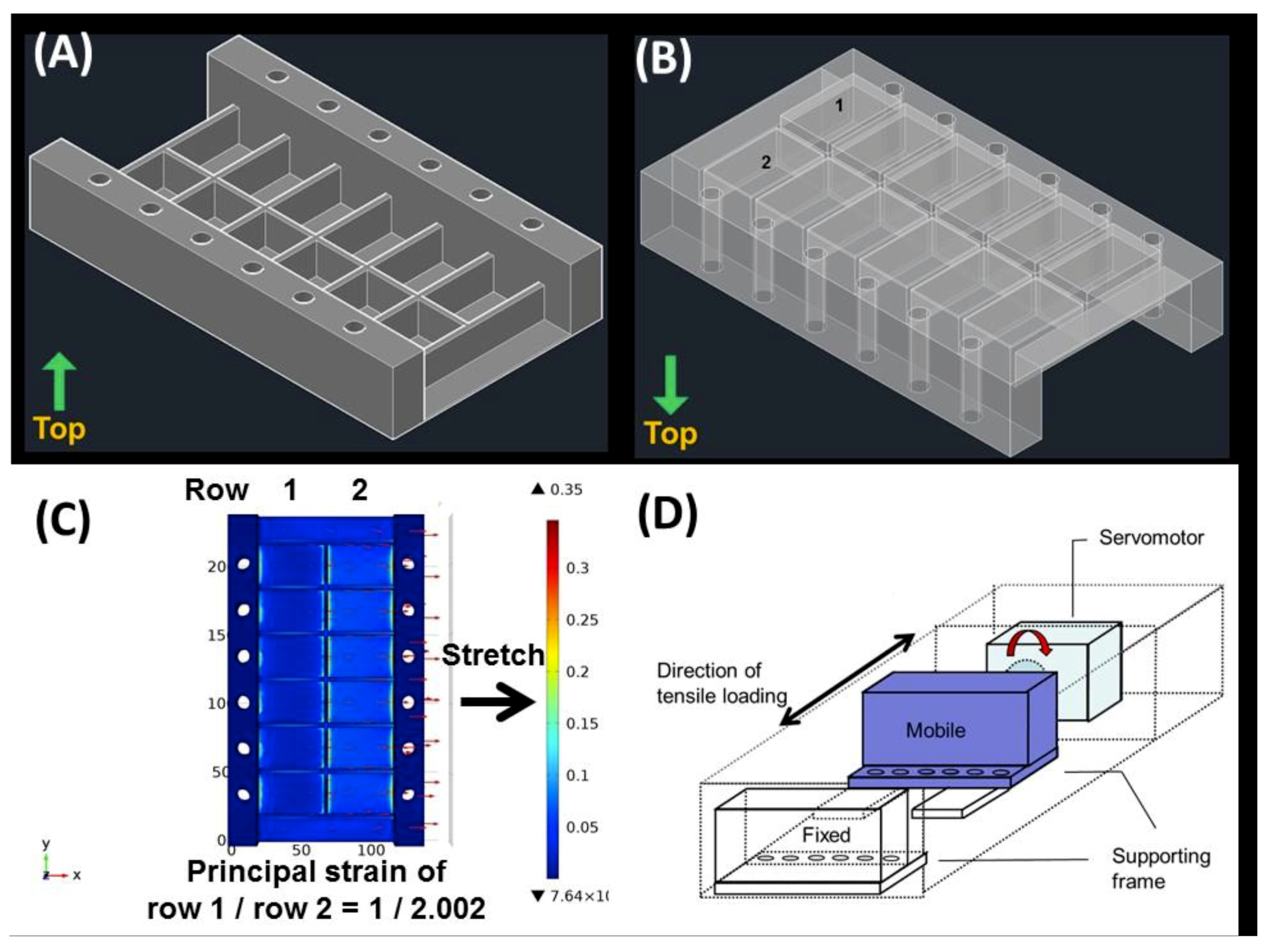
2.4. Differential Uniaxial Stretch Culture Device
3. Discussion
4. Materials and Methods
4.1. Study Protocol
4.2. Differential Uniaxial Stretch Culture Device
4.3. Isolation and Culture of TDCs
4.4. Flow Cytometry Assay
4.5. Immunohistochemistry
4.6. Quantitative PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EGCG | epigallocatechin gallate |
| TDCs | tendon-derived cells |
| ECM | extracellular matrix |
| ROS | reactive oxygen species |
| PCR | polymerase chain reaction |
| PBS | phosphate-buffered saline |
| FBS | fetal bovine serum |
| PDMS | polydimethylsiloxane |
| PE | phycoerythrin |
| FITC | fluorescein isothiocyanate |
| FACS | fluorescence-activated cell sorting |
References
- Kaux, J.F.; Forthomme, B.; Goff, C.L.; Crielaard, J.M.; Croisier, J.L. Current opinions on tendinopathy. J. Sports Sci. Med. 2011, 10, 238–253. [Google Scholar] [PubMed]
- Reinking, M. Tendinopathy in athletes. Phys. Ther. Sport 2012, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Arnoczky, S.P.; Lavagnino, M.; Egerbacher, M. The mechanobiological aetiopathogenesis of tendinopathy: is it the over-stimulation or the under-stimulation of tendon cells? Int. J. Exp. Pathol. 2007, 88, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Riley, G. Tendinopathy—From basic science to treatment. Nat. Clin. Pract. Rheumatol. 2008, 4, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Kaleagasioglu, F.; Olcay, E. Fluoroquinolone-induced tendinopathy: Etiology and preventive measures. Tohoku J. Exp. Med. 2012, 226, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Longo, U.G.; Berton, A.; Papapietro, N.; Maffulli, N.; Denaro, V. Epidemiology, genetics and biological factors of rotator cuff tears. Med. Sport Sci. 2012, 57, 1–9. [Google Scholar] [PubMed]
- Longo, U.G.; Oliva, F.; Denaro, V.; Maffulli, N. Oxygen species and overuse tendinopathy in athletes. Disabil. Rehabil. 2008, 30, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Role of metabolic H2O2 generation: redox signaling and oxidative stress. J. Biol. Chem. 2014, 289, 8735–8741. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Stoppani, E.; Volonte, D.; Galbiati, F. Caveolin-1, cellular senescence and age-related diseases. Mech. Ageing Dev. 2011, 132, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Rui, Y.F.; Lui, P.P.; Ni, M.; Chan, L.S.; Lee, Y.W.; Chan, K.M. Mechanical loading increased BMP-2 expression which promoted osteogenic differentiation of tendon-derived stem cells. J. Orthop. Res. 2011, 29, 390–396. [Google Scholar] [CrossRef]
- Shi, Y.; Fu, Y.; Tong, W.; Geng, Y.; Lui, P.P.; Tang, T.; Zhang, X.; Dai, K. Uniaxial mechanical tension promoted osteogenic differentiation of rat tendon-derived stem cells (rTDSCs) via the Wnt5a-RhoA pathway. J. Cell. Biochem. 2012, 113, 3133–3142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, J.H. Mechanobiological response of tendon stem cells: implications of tendon homeostasis and pathogenesis of tendinopathy. J. Orthop. Res. 2010, 28, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, G.; Zhang, X.; Wang, L.; Du, Y.; Lu, T.J.; Xu, F. Engineering cell alignment in vitro. Biotechnol. Adv. 2014, 32, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Lui, P.P.; Chan, K.M. Tendon-derived stem cells (TDSCs): from basic science to potential roles in tendon pathology and tissue engineering applications. Stem Cell Rev. 2011, 7, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Safer, A.M.; Menon, M. Green tea polyphenols and their potential role in health and disease. Inflammopharmacology 2015, 23, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, S.A.; Serafim, K.G.; Mizokami, S.S.; Hohmann, M.S.; Casagrande, R.; Verri, W.A., Jr. Analgesic activity of piracetam: effect on cytokine production and oxidative stress. Pharmacol. Biochem. Behav. 2013, 105, 183–192. [Google Scholar] [CrossRef]
- Verma, D.K.; Joshi, N.; Raju, K.S.; Wahajuddin, M.; Singh, R.K.; Singh, S. Metabolic enhancer piracetam attenuates rotenone induced oxidative stress: a study in different rat brain regions. Acta Neurobiol. Exp. (Wars) 2015, 75, 399–411. [Google Scholar]
- Winblad, B. Piracetam: A review of pharmacological properties and clinical uses. CNS Drug Rev. 2005, 11, 169–182. [Google Scholar] [CrossRef]
- Fatima, S.; Al-Mohaimeed, N.; Al-Shaikh, Y.; Tyagi, P.; Banu, N.; Hasan, S.; Arjumand, S. Combined treatment of epigallocatechin gallate and Coenzyme Q10 attenuates cisplatin-induced nephrotoxicity via suppression of oxidative/nitrosative stress, inflammation and cellular damage. Food Chem. Toxicol. 2016, 94, 213–220. [Google Scholar] [CrossRef]
- Krupkova, O.; Handa, J.; Hlavna, M.; Klasen, J.; Ospelt, C.; Ferguson, S.J.; Wuertz-Kozak, K. The Natural Polyphenol Epigallocatechin Gallate Protects Intervertebral Disc Cells from Oxidative Stress. Oxid. Med. Cell. Longev. 2016. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Hu, M.; Zha, Y.H.; Li, Z.C.; Zhao, B.; Yu, L.L.; Yu, M.; Qian, Y. Piracetam ameliorated oxygen and glucose deprivation-induced injury in rat cortical neurons via inhibition of oxidative stress, excitatory amino acids release and P53/Bax. Cell. Mol. Neurobiol. 2014, 34, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Kalkan, E.; Keskin, F.; Kaya, B.; Esen, H.; Tosun, M.; Kalkan, S.S.; Erdi, F.; Unlü, A.; Avunduk, M.C.; Cicek, O. Effects of iloprost and piracetam in spinal cord ischemia-reperfusion injury in the rabbit. Spinal Cord 2011, 49, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Tuncer, S.; Ayhan, S.; Findikcioglu, K.; Ergun, H.; Tuncer, I. Effect of systemic piracetam treatment on flap survival and vascular endothelial growth factor expression after ischemia-reperfusion injury. J. Reconstr. Microsurg. 2011, 27, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Tuglu, D.; Yuvanc, E.; Ozan, T.; Bal, F.; Yilmaz, E.; Atasoy, P.; Kisa, U.; Batislam, E. Protective effects of udenafil citrate, piracetam and dexmedetomidine treatment on testicular torsion/detorsion-induced ischaemia/reperfusion injury in rats. Andrologia 2016, 48, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Joo, S.D.; Han, S.B.; Im, J.; Lee, S.H.; Sonn, C.H.; Lee, K.M. Isolation and expansion of synovial CD34(-)CD44(+)CD90(+) mesenchymal stem cells: Comparison of an enzymatic method and a direct explant technique. Connect. Tissue Res. 2011, 52, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Elsaadany, M.; Winters, K.; Adams, S.; Stasuk, A.; Ayan, H.; Yildirim-Ayan, E. Equiaxial Strain Modulates Adipose-derived Stem Cell Differentiation within 3D Biphasic Scaffolds towards Annulus Fibrosus. Sci. Rep. 2017, 7, 12868. [Google Scholar] [CrossRef]
- Lui, P.P.; Chan, L.S.; Lee, Y.W.; Fu, S.C.; Chan, K.M. Sustained expression of proteoglycans and collagen type III/type I ratio in a calcified tendinopathy model. Rheumatology (Oxford) 2010, 49, 231–239. [Google Scholar] [CrossRef]
- Samiric, T.; Parkinson, J.; Ilic, M.Z.; Cook, J.; Feller, J.A.; Handley, C.J. Changes in the composition of the extracellular matrix in patellar tendinopathy. Matrix Biol. 2009, 28, 230–236. [Google Scholar]
- Chaudhury, S.; Carr, A.J. Lessons we can learn from gene expression patterns in rotator cuff tears and tendinopathies. J. Shoulder Elbow Surg. 2012, 21, 191–199. [Google Scholar] [CrossRef]
- Lui, P.P. Histopathological changes in tendinopathy—Potential roles of BMPs? Rheumatology (Oxford) 2013, 52, 2116–2126. [Google Scholar] [CrossRef] [PubMed]
- Lui, P.P.Y.; Fu, S.C.; Chan, L.S.; Hung, L.K.; Chan, K.M. Chondrocyte Phenotype and Ectopic Ossification in Collagenase-induced Tendon Degeneration. J. Histochem. Cytochem. 2009, 57, 91–100. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, M.-Y.; Lin, P.-C.; Liao, W.-H.; Chen, W.-S.; Hsu, C.-H.; He, C.-K.; Wu, Y.-W.; Gefen, A.; Iafisco, M.; Liu, L.; et al. The Effect of the Repression of Oxidative Stress on Tenocyte Differentiation: A Preliminary Study of a Rat Cell Model Using a Novel Differential Tensile Strain Bioreactor. Int. J. Mol. Sci. 2019, 20, 3437. https://doi.org/10.3390/ijms20143437
Hsiao M-Y, Lin P-C, Liao W-H, Chen W-S, Hsu C-H, He C-K, Wu Y-W, Gefen A, Iafisco M, Liu L, et al. The Effect of the Repression of Oxidative Stress on Tenocyte Differentiation: A Preliminary Study of a Rat Cell Model Using a Novel Differential Tensile Strain Bioreactor. International Journal of Molecular Sciences. 2019; 20(14):3437. https://doi.org/10.3390/ijms20143437
Chicago/Turabian StyleHsiao, Ming-Yen, Ping-Cheng Lin, Wei-Hao Liao, Wen-Shiang Chen, Chia-Hsien Hsu, Cheng-Kun He, Ya-Wen Wu, Amit Gefen, Michele Iafisco, Lixin Liu, and et al. 2019. "The Effect of the Repression of Oxidative Stress on Tenocyte Differentiation: A Preliminary Study of a Rat Cell Model Using a Novel Differential Tensile Strain Bioreactor" International Journal of Molecular Sciences 20, no. 14: 3437. https://doi.org/10.3390/ijms20143437