Do DLX3 and CD271 Protect Human Keratinocytes from Squamous Tumor Development?





Abstract
:1. Introduction
2. Results and Discussion
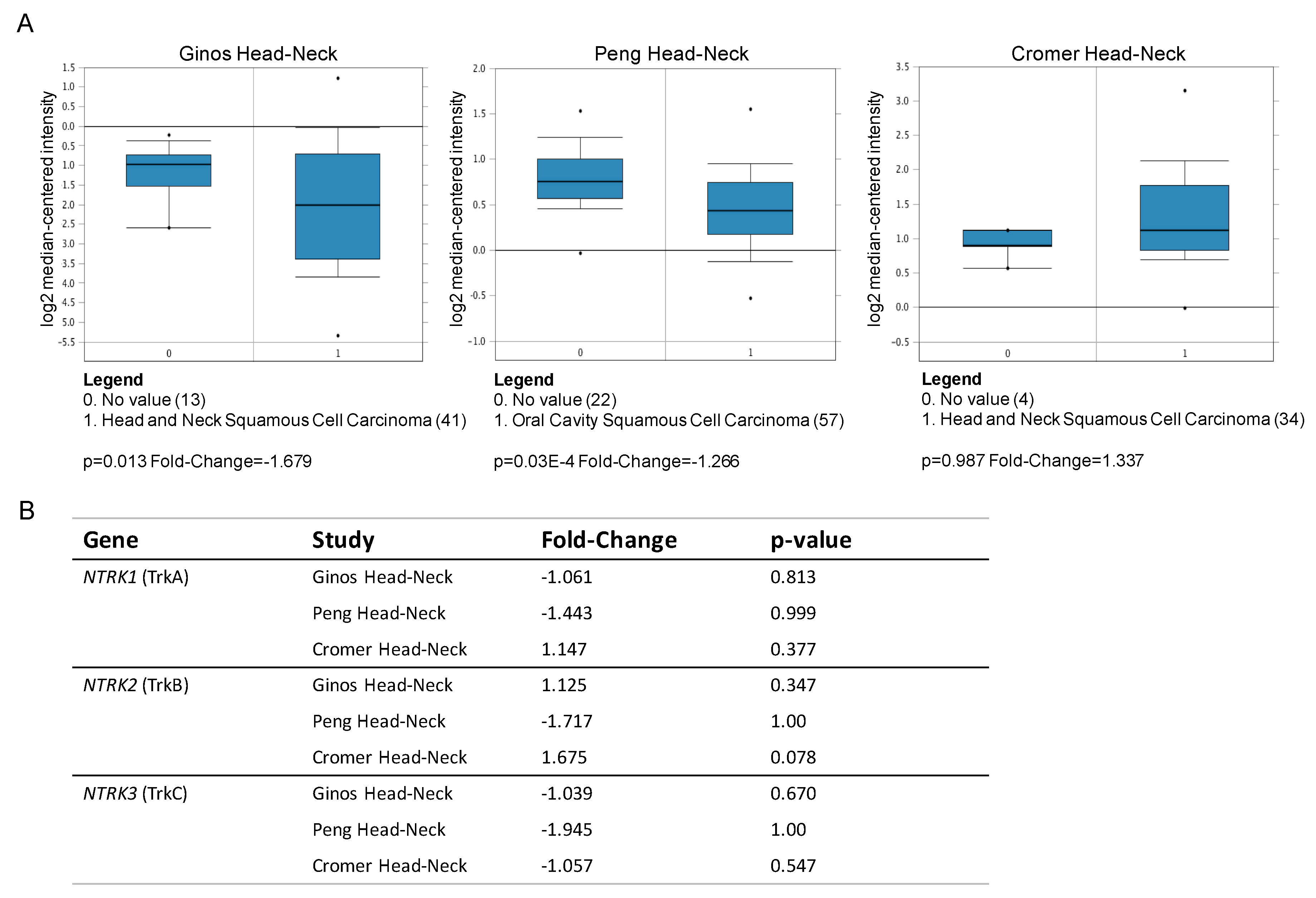
2.1. NT Receptors Are Heterogeneously Expressed in Squamous Cancer
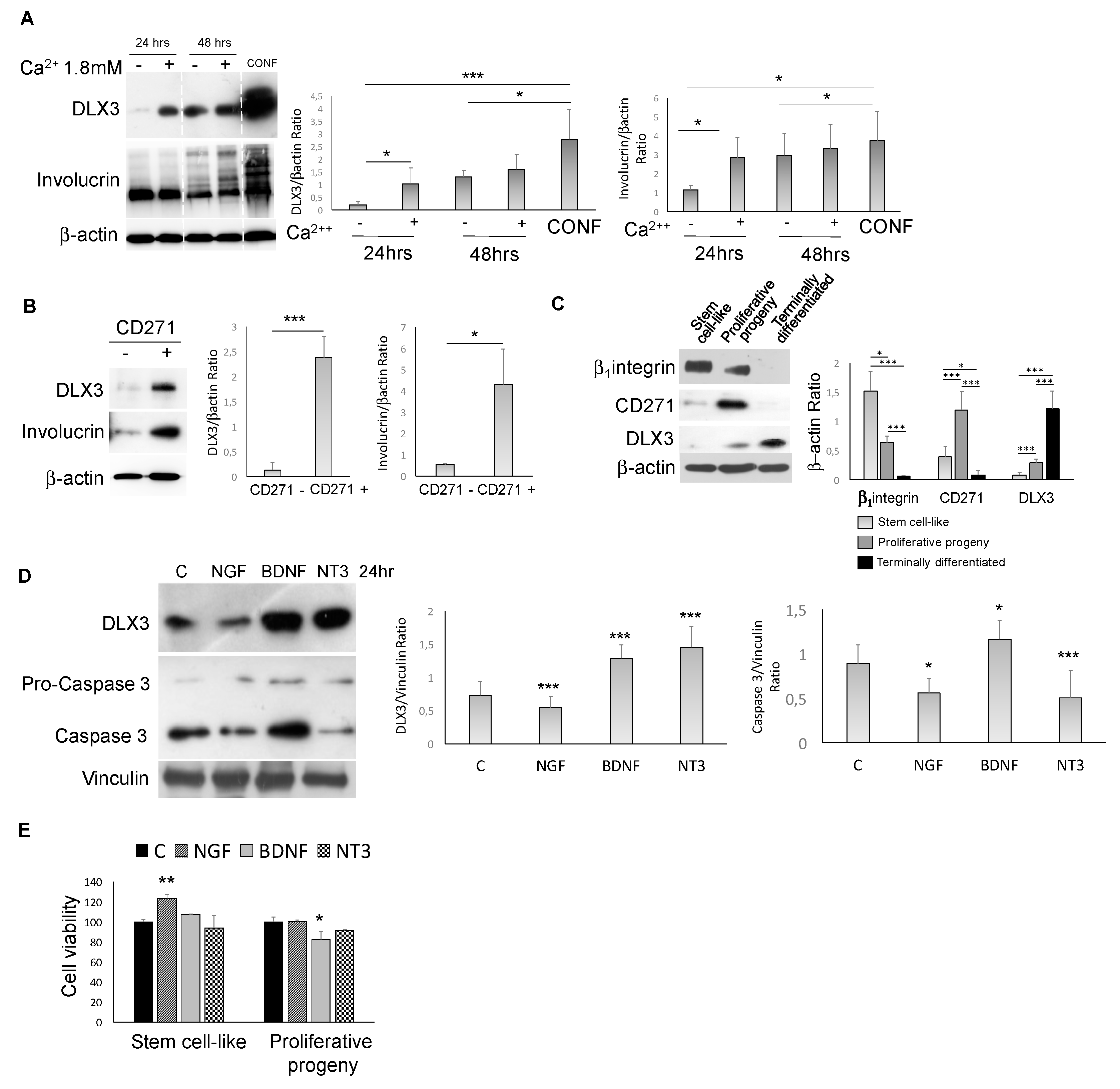
2.2. DLX3-CD271 Feedback Loop in Human Healthy Keratinocytes
3. Conclusions
4. Materials and Methods
4.1. Isolation of Primary Keratinocytes from cSCC Tissues
4.2. Isolation of Primary Keratinocytes from Healthy Skin and SCC Cell Line Cultures
4.3. Keratinocyte Treatment with Neurotrophins and Transduction
4.4. Multicellular Spheroids Culture
4.5. Picture Analysis
4.6. Western Blotting
4.7. Cell Sorting
4.8. FACS Analysis
4.9. MTT Assay
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fuchs, E.; Raghavan, S. Getting under the skin of epidermal morphogenesis. Nat. Rev. Genet. 2002, 3, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Que, S.K.; Zwald, F.O.; Schmults, C.D. Cutaneous squamous cell carcinoma: Incidence, risk factors, diagnosis, and staging. J. Am. Acad. Dermatol. 2018, 78, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Radoja, N.; Gazel, A.; Banno, T.; Yano, S.; Blumenberg, M. Transcriptional profiling of epidermal differentiation. Physiol. Genom. 2006, 27, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Dellambra, E. Oncogenic Ras: A double-edged sword for human epidermal stem and transient amplifying cells. Small GTPases 2016, 7, 147–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Liu, Z.; Myers, J.N. TP53 Mutations in Head and Neck Squamous Cell Carcinoma and Their Impact on Disease Progression and Treatment Response. J. Cell. Biochem. 2016, 117, 2682–2692. [Google Scholar] [CrossRef] [PubMed]
- Park, G.T.; Morasso, M.I. Regulation of the Dlx3 homeobox gene upon differentiation of mouse keratinocytes. J. Biol. Chem. 1999, 274, 26599–26608. [Google Scholar] [CrossRef]
- Palazzo, E.; Kellett, M.; Cataisson, C.; Gormley, A.; Bible, P.W.; Pietroni, V.; Radoja, N.; Hwang, J.; Blumenberg, M.; Yuspa, S.H.; et al. The homeoprotein DLX3 and tumor suppressor p53 co-regulate cell cycle progression and squamous tumor growth. Oncogene 2016, 35, 3114. [Google Scholar] [CrossRef]
- Hwang, J.; Kita, R.; Kwon, H.S.; Choi, E.H.; Lee, S.H.; Udey, M.C.; Morasso, M.I. Epidermal ablation of Dlx3 is linked to IL-17-associated skin inflammation. Proc. Natl. Acad. Sci. USA 2011, 108, 11566–11571. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Kim, J.C.; Ogawa, Y.; Nakato, G.; Nagle, V.; Brooks, S.R.; Udey, M.C.; Morasso, M.I. DLX3-Dependent STAT3 Signaling in Keratinocytes Regulates Skin Immune Homeostasis. J. Investig. Dermatol. 2018, 138, 1052–1061. [Google Scholar] [CrossRef] [Green Version]
- Botchkarev, V.A.; Botchkareva, N.V.; Peters, E.M.; Paus, R. Epithelial growth control by neurotrophins: Leads and lessons from the hair follicle. Prog. Brain Res. 2004, 146, 493–513. [Google Scholar]
- Truzzi, F.; Saltari, A.; Palazzo, E.; Lotti, R.; Petrachi, T.; Dallaglio, K.; Gemelli, C.; Grisendi, G.; Dominici, M.; Pincelli, C.; et al. CD271 mediates stem cells to early progeny transition in human epidermis. J. Investig. Dermatol. 2015, 135, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Truzzi, F.; Marconi, A.; Atzei, P.; Panza, M.C.; Lotti, R.; Dallaglio, K.; Tiberio, R.; Palazzo, E.; Vaschieri, C.; Pincelli, C. p75 neurotrophin receptor mediates apoptosis in transit-amplifying cells and its overexpression restores cell death in psoriatic keratinocytes. Cell Death Differ. 2011, 18, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D. The neurotrophin family of neurotrophic factors: An overview. Methods Mol. Biol. 2012, 846, 1–12. [Google Scholar] [PubMed]
- Gentry, J.J.; Barker, P.A.; Carter, B.D. The p75 neurotrophin receptor: Multiple interactors and numerous functions. Prog. Brain Res. 2004, 146, 25–39. [Google Scholar] [PubMed]
- Khotskaya, Y.B.; Holla, V.R.; Farago, A.F.; Mills Shaw, K.R.; Meric-Bernstam, F.; Hong, D.S. Targeting TRK family proteins in cancer. Pharmacol. Ther. 2017, 173, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Doebele, R.C.; Davis, L.E.; Vaishnavi, A.; Le, A.T.; Estrada-Bernal, A.; Keysar, S.; Jimeno, A.; Varella-Garcia, M.; Aisner, D.L.; Li, Y.; et al. An Oncogenic NTRK Fusion in a Patient with Soft-Tissue Sarcoma with Response to the Tropomyosin-Related Kinase Inhibitor LOXO-101. Cancer Discov. 2015, 5, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Rolfo, C.; Ruiz, R.; Giovannetti, E.; Gil-Bazo, I.; Russo, A.; Passiglia, F.; Giallombardo, M.; Peeters, M.; Raez, L. Entrectinib: A potent new TRK, ROS1, and ALK inhibitor. Expert Opin. Investig. Drugs 2015, 24, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jiffar, T.; Kupferman, M.E. A novel role for BDNF-TrkB in the regulation of chemotherapy resistance in head and neck squamous cell carcinoma. PLoS ONE 2012, 7, e30246. [Google Scholar] [CrossRef] [PubMed]
- Dallaglio, K.; Petrachi, T.; Marconi, A.; Truzzi, F.; Lotti, R.; Saltari, A.; Morandi, P.; Puviani, M.; Maiorana, A.; Pincelli, C. Expression of nuclear survivin in normal skin and squamous cell carcinoma: A possible role in tumour invasion. Br. J. Cancer 2014, 110, 199–207. [Google Scholar] [CrossRef]
- Huang, S.D.; Yuan, Y.; Liu, X.H.; Gong, D.J.; Bai, C.G.; Wang, F.; Luo, J.H.; Xu, Z.Y. Self-renewal and chemotherapy resistance of p75NTR positive cells in esophageal squamous cell carcinomas. BMC Cancer 2009, 9, 9. [Google Scholar] [CrossRef]
- Kiyosue, T.; Kawano, S.; Matsubara, R.; Goto, Y.; Hirano, M.; Jinno, T.; Toyoshima, T.; Kitamura, R.; Oobu, K.; Nakamura, S. Immunohistochemical location of the p75 neurotrophin receptor (p75NTR) in oral leukoplakia and oral squamous cell carcinoma. Int. J. Clin. Oncol. 2013, 18, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Sauca, O.; Chung, M.K.; Shin, J.H.; Karamboulas, C.; Kwok, S.; Jung, Y.H.; Oakley, R.; Tysome, J.R.; Farnebo, L.O.; Kaplan, M.J.; et al. CD271 is a functional and targetable marker of tumor-initiating cells in head and neck squamous cell carcinoma. Oncotarget 2014, 5, 6854–6866. [Google Scholar] [CrossRef] [PubMed]
- Rheinwald, J.G.; Beckett, M.A. Tumorigenic keratinocyte lines requiring anchorage and fibroblast support cultured from human squamous cell carcinomas. Cancer Res. 1981, 41, 1657–1663. [Google Scholar] [PubMed]
- Saltari, A.; Truzzi, F.; Quadri, M.; Lotti, R.; Palazzo, E.; Grisendi, G.; Tiso, N.; Marconi, A.; Pincelli, C. CD271 Down-Regulation Promotes Melanoma Progression and Invasion in Three-Dimensional Models and in Zebrafish. J. Investig. Dermatol. 2016, 136, 2049–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marconi, A.; Terracina, M.; Fila, C.; Franchi, J.; Bonté, F.; Romagnoli, G.; Maurelli, R.; Failla, C.M.; Dumas, M.; Pincelli, C. Expression and function of neurotrophins and their receptors in cultured human keratinocytes. J. Investig. Dermatol. 2003, 121, 1515–1521, Erratum in 2004, 123, 803. [Google Scholar]
- Lotti, R.; Palazzo, E.; Petrachi, T.; Dallaglio, K.; Saltari, A.; Truzzi, F.; Quadri, M.; Puviani, M.; Maiorana, A.; Marconi, A.; et al. Survivin Modulates Squamous Cell Carcinoma-Derived Stem-Like Cell Proliferation, Viability and Tumor Formation in Vivo. Int. J. Mol. Sci. 2016, 17, 89. [Google Scholar] [CrossRef] [PubMed]
- Gandarillas, A.; Watt, F.M. c-Myc promotes differentiation of human epidermal stem cells. Genes Dev. 1997, 11, 2869–2882. [Google Scholar] [CrossRef] [Green Version]
- Pincelli, C.; Marconi, A. Autocrine nerve growth factor in human keratinocytes. J. Dermatol. Sci. 2000, 22, 71–79. [Google Scholar] [CrossRef]
- Carlsson, J.; Yuhas, J.M. Liquid-overlay culture of cellular spheroids. In Recent Results in Cancer Research: Spheroids in Cancer Research; Springer: Berlin/Heidelberg, Germany, 1984; Volume 95, pp. 1–23. [Google Scholar]
- De Wever, O.; Hendrix, A.; De Boeck, A.; Westbroek, W.; Braems, G.; Emami, S.; Sabbah, M.; Gespach, C.; Bracke, M. Modeling and quantification of cancer cell invasion through collagen type I matrices. Int. J. Dev. Biol. 2010, 54, 887–896. [Google Scholar] [CrossRef]
- Hwang, J.; Mehrani, T.; Millar, S.E.; Morasso, M.I. Dlx3 is a crucial regulator of hair follicle differentiation and cycling. Development 2008, 135, 3149–3159. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NT Receptors | Fold-Change |
|---|---|
| CD271 (NGFR) | 3.19 |
| TrkA (NTRK1) | 1.19 |
| TrkB (NTRK2) | −1.07 |
| TrkC (NTRK3) | −1.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palazzo, E.; Marconi, A.; Pincelli, C.; Morasso, M.I. Do DLX3 and CD271 Protect Human Keratinocytes from Squamous Tumor Development? Int. J. Mol. Sci. 2019, 20, 3541. https://doi.org/10.3390/ijms20143541
Palazzo E, Marconi A, Pincelli C, Morasso MI. Do DLX3 and CD271 Protect Human Keratinocytes from Squamous Tumor Development? International Journal of Molecular Sciences. 2019; 20(14):3541. https://doi.org/10.3390/ijms20143541
Chicago/Turabian StylePalazzo, Elisabetta, Alessandra Marconi, Carlo Pincelli, and Maria I. Morasso. 2019. "Do DLX3 and CD271 Protect Human Keratinocytes from Squamous Tumor Development?" International Journal of Molecular Sciences 20, no. 14: 3541. https://doi.org/10.3390/ijms20143541
APA StylePalazzo, E., Marconi, A., Pincelli, C., & Morasso, M. I. (2019). Do DLX3 and CD271 Protect Human Keratinocytes from Squamous Tumor Development? International Journal of Molecular Sciences, 20(14), 3541. https://doi.org/10.3390/ijms20143541