Comparison of Lipid and Palmitoleic Acid Induction of Tribonema minus under Heterotrophic and Phototrophic Regimes by Using High-Density Fermented Seeds

Abstract
:1. Introduction
2. Results
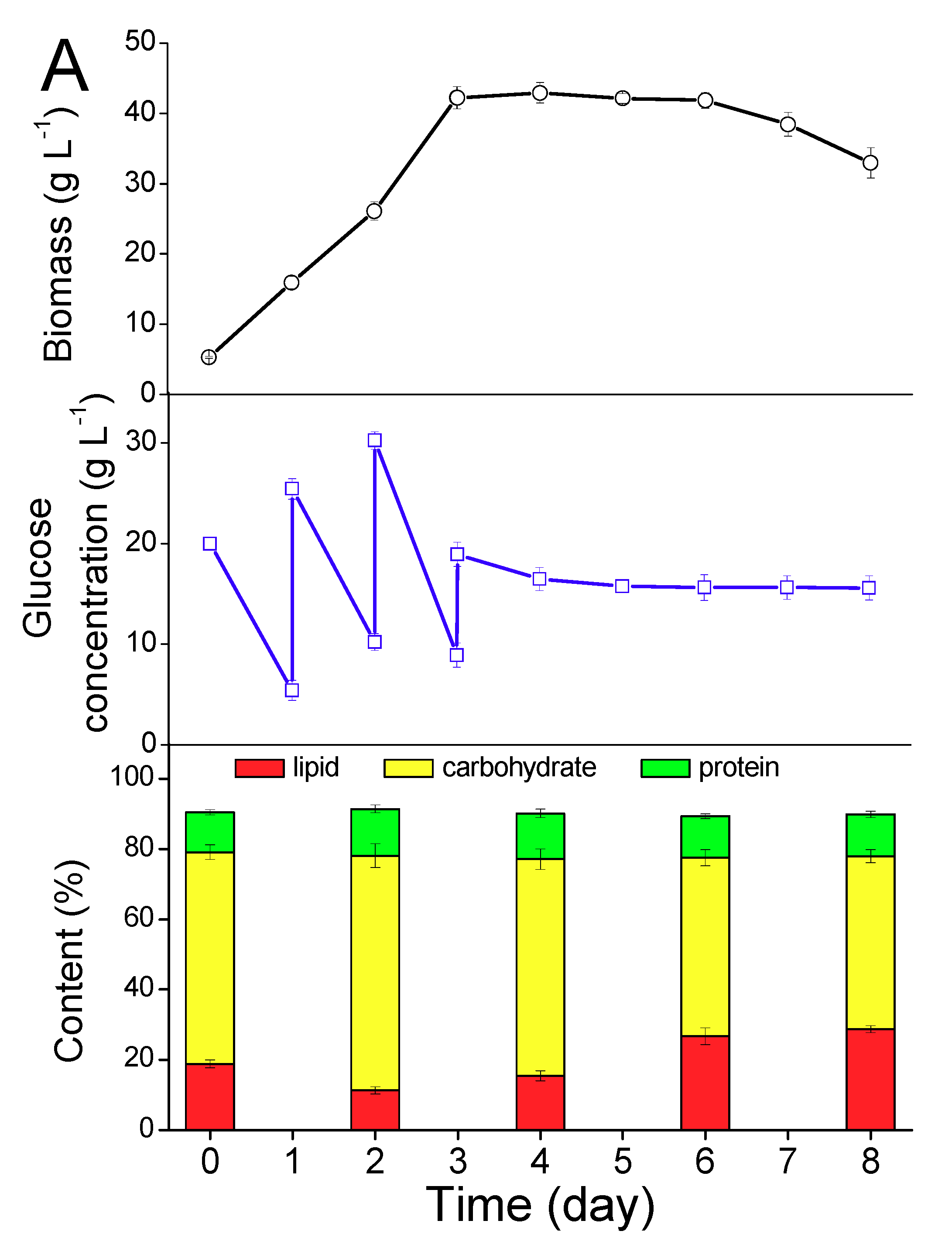
2.1. Cultivation and Lipid Accumulation of T. minus by Heterotrophy
2.1.1. Growth
2.1.2. Lipid Accumulation
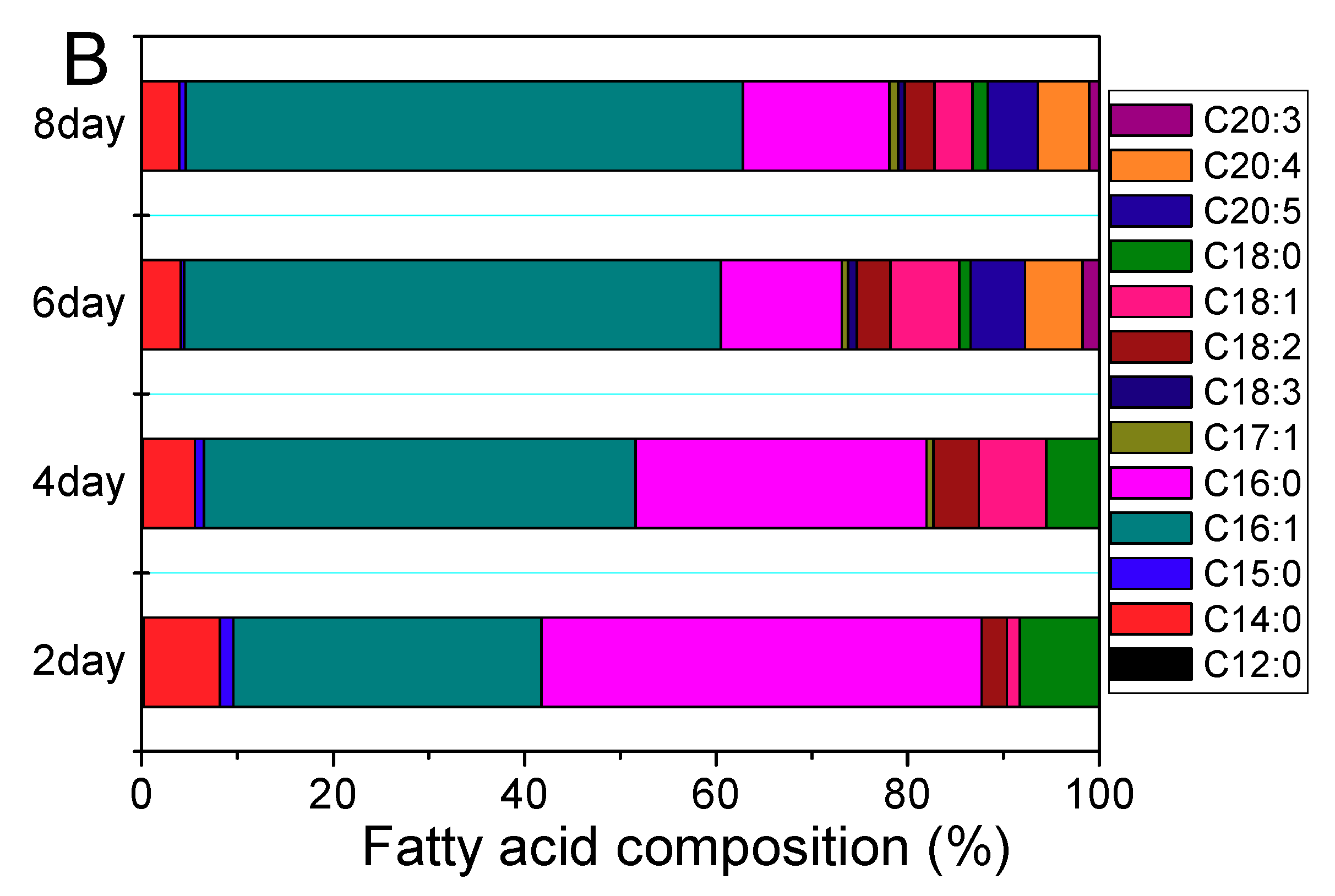
2.1.3. Palmitoleic Acid Accumulation
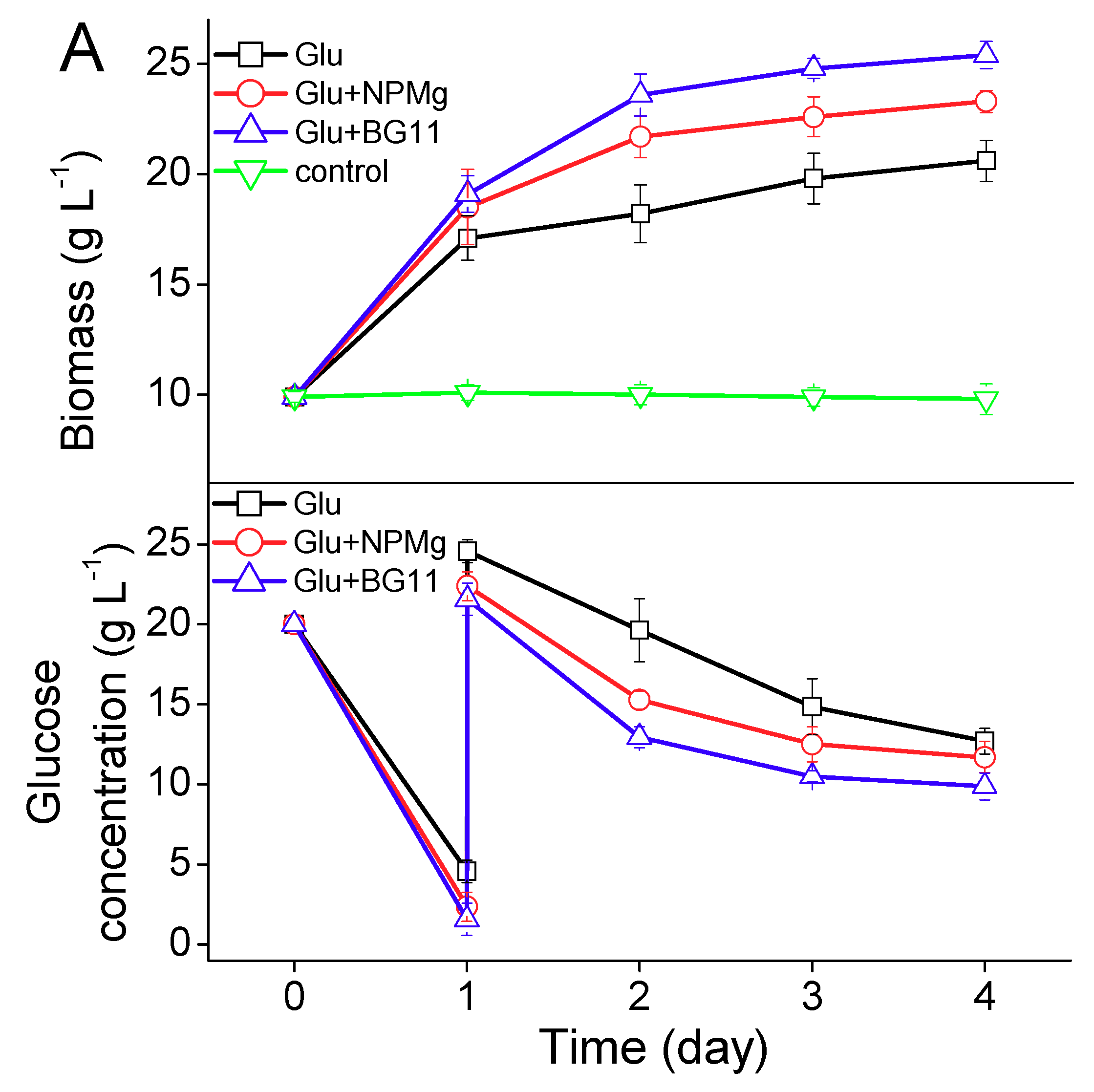
2.2. Lipid Induction under Different Regimes
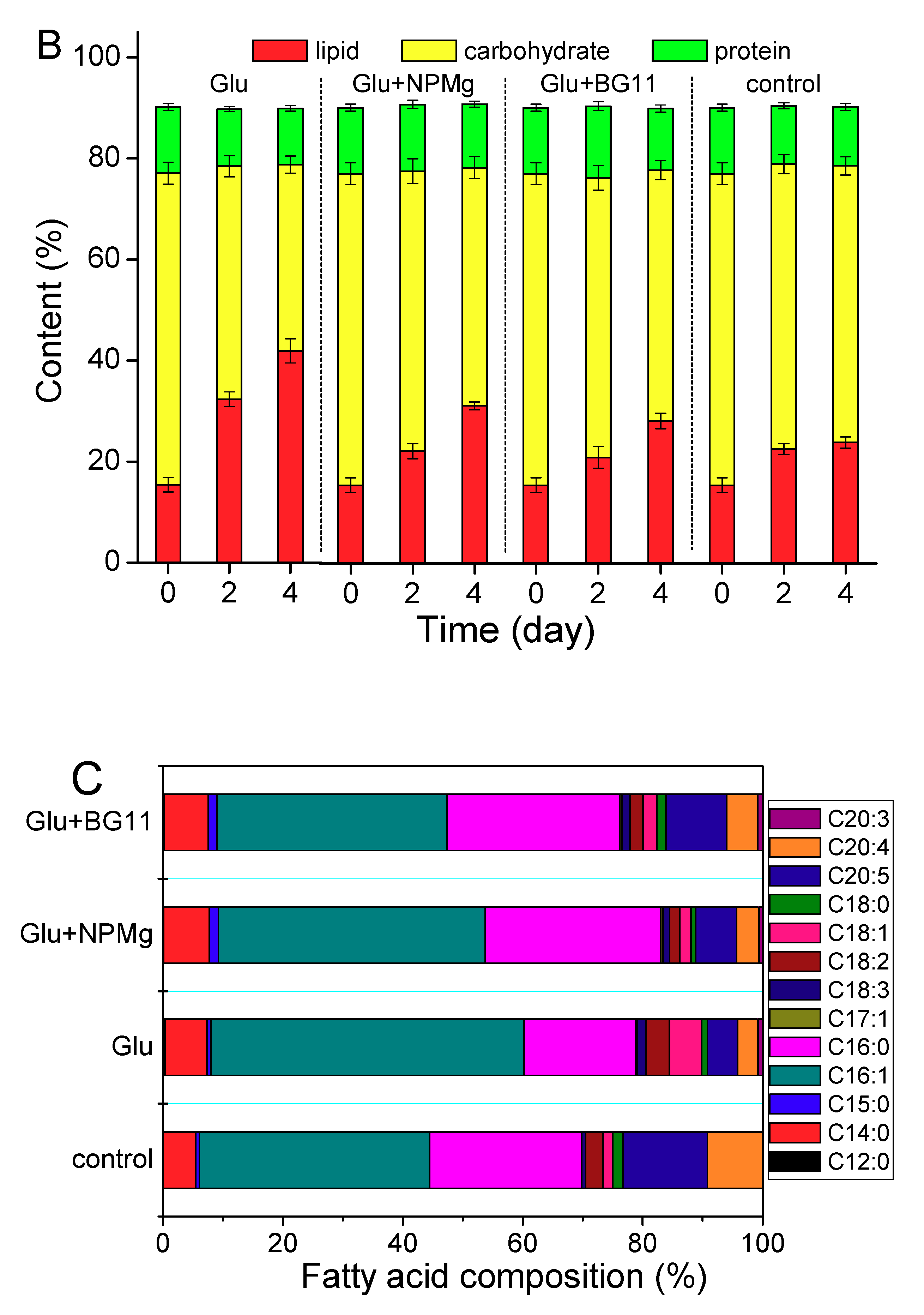
2.2.1. Lipid Induction by Heterotrophy
2.2.2. Lipid induction by Phototrophy
3. Discussion
4. Materials and Methods
4.1. Microalga and Culture Conditions
4.2. Experimental Design
4.3. Analytical Methods
4.4. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- De Souza, C.O.; Vannice, G.K.; Rosa Neto, J.C.; Calder, P.C. Is palmitoleic acid a plausible nonpharmacological strategy to prevent or control chronic metabolic and inflammatory disorders? Mol. Nutr. Food Res. 2018, 62, 1700504. [Google Scholar] [CrossRef]
- Cao, H.M.; Gerhold, K.; Mayers, J.R.; Wiest, M.M.; Watkins, S.M.; Hotamisligil, G.S. Identification of a lipokine, a lipid hormone linking adipose tissue to systemic metabolism. Cell 2008, 134, 933–944. [Google Scholar] [CrossRef]
- Souza, C.O.; Teixeira, A.A.S.; Lima, E.A.; Batatinha, H.A.P.; Gomes, L.M.; Carvalho-Silva, M.; Mota, I.T.; Streck, E.L.; Hirabara, S.M.; Rosa Neto, J.C. Palmitoleic acid (n-7) attenuates the immunometabolic disturbances caused by a high-fat diet independently of PPARα. Med. Inflamm. 2014, 2014, 582197. [Google Scholar] [CrossRef]
- Guo, X.; Li, H.G.; Xu, H.; Halim, V.; Zhang, W.Y.; Wang, H.; Ong, K.T.; Woo, S.L.; Walzem, R.L.; Mashek, D.G.; et al. Palmitoleate induces hepatic steatosis but suppresses liver inflammatory response in mice. PLoS ONE 2012, 7, e39286. [Google Scholar] [CrossRef]
- Çimen, I.; Kocatürk, B.; Koyuncu, S.; Tufanlı, Ö.; Onat, U.I.; Yıldırım, A.D.; Apaydın, O.; Demirsoy, Ş.; Aykut, Z.G.; Nguyen, U.T.; et al. Prevention of atherosclerosis by bioactive palmitoleate through suppression of organelle stress and inflammasome activation. Sci. Trans. Med. 2016, 8, 358ra126. [Google Scholar] [CrossRef]
- Souza, C.O.; Teixeira, A.A.S.; Biondo, L.A.; Silveira, L.S.; Calder, P.C.; Rosa Neto, J.C. Palmitoleic acid reduces the inflammation in LPS-stimulated macrophages by inhibition of NFκB, independently of PPARs. Clin. Exp. Pharmacol. Physiol. 2017, 44, 566–575. [Google Scholar] [CrossRef]
- Chan, K.L.; Pillon, N.J.; Sivaloganathan, D.M.; Costford, S.R.; Liu, Z.; Théret, M.; Chazaud, B.; Klip, A. Palmitoleate reverses high fat-induced pro-inflammatory macrophage polarization via AMPK. J. Biol. Chem. 2015, 290, 16979–16988. [Google Scholar] [CrossRef]
- Yang, Z.H.; Miyahara, H.; Hatanaka, A. Chronic administration of palmitoleic acid reduces insulin resistance and hepatic lipid accumulation in KK-Ay Mice with genetic type 2. Lipids Health Dis. 2011, 10, 120. [Google Scholar] [CrossRef]
- Yang, Z.H.; Takeo, J.; Katayama, M. Oral administration of omega-7 palmitoleic acid induces satiety and the release of appetite-related hormones in male rats. Appetite 2013, 65, 1–7. [Google Scholar] [CrossRef]
- Knothe, G. Biodiesel derived from a model oil enriched in palmitoleic acid, macadamia nut oil. Energy Fuels 2010, 24, 2098–2103. [Google Scholar] [CrossRef]
- Cao, Y.J.; Liu, W.; Xu, X.; Zhang, H.B.; Wang, J.M.; Xian, M. Production of free monounsaturated fatty acids by metabolically engineered Escherichia coli. Biotechnol. Biofuels 2014, 7, 59–69. [Google Scholar] [CrossRef]
- Rybak, A.; Fokou, P.A.; Meier, M.A.R. Metathesis as a versatile tool in oleochemistry. Eur. J. Lipid Sci. Technol. 2008, 110, 797–804. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Mishra, G.; Whittle, E.; Pidowich, M.S.; Bevan, S.A.; Merlo, A.O.; Walsh, T.A.; Shanklin, J. Metabolic engineering of seeds can achieve levels of omega-7 fatty acids comparable to the highest levels found in natural plant sources. Plant Physiol. 2010, 154, 1897–1904. [Google Scholar] [CrossRef]
- Wu, Y.M.; Li, R.Z.; Hildebrand, D.F. Biosynthesis and metabolic engineering of palmitoleate production, an important contributor to human health and sustainable industry. Prog. Lipid Res. 2012, 51, 340–349. [Google Scholar] [CrossRef]
- Yang, B.R.; Kallio, H.P. Fatty acid composition of lipids in sea buckthorn (Hippophae rhamnoides L.) berries of different origins. J. Agric. Food Chem. 2001, 49, 1939–1947. [Google Scholar] [CrossRef]
- Salas, J.J.; Martinez-Force, E.; Garces, R. Biochemcial characterization of a high palmitoleic acid Helianthus annuus mutant. Plant Physiol. Biochem. 2004, 42, 373–381. [Google Scholar] [CrossRef]
- Okullo, A.A.; Temu, A.K.; Ogwok, P.; Ntalikwa, J.W. Physico-chemical properties of biodiesel from jatropha and castor oils. Int. J. Renew. Energy Res. 2012, 2, 47–52. [Google Scholar]
- De Swaaf, M.; Sijtsma, L.; Pronk, J. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing marine alga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef]
- Xiong, W.; Li, X.F.; Xiang, J.Y.; Wu, Q.Y. High-density fermentation of microalga Chlorella protothecoides in bioreactor for microbio-diesel production. Appl. Microbiol. Biotechnol. 2008, 78, 29–36. [Google Scholar] [CrossRef]
- Wei, A.L.; Zhang, X.W.; Wei, D.; Chen, G.; Wu, Q.Y.; Yang, S.T. Effects of cassava starch hydrolysate on cell growth and lipid accumulation of the heterotrophic microalgae Chlorella protothecoides. J. Ind. Microbiol. Biotechnol. 2009, 36, 1383–1389. [Google Scholar] [CrossRef]
- Han, F.F.; Huang, J.K.; Li, Y.G.; Wang, W.L.; Wang, J.; Fan, J.H.; Shen, G.M. Enhancement of microalgal biomass and lipid productivities by a model of photoautotrophic culture with heterotrophic cells as seed. Bioresour. Technol. 2012, 118, 431–437. [Google Scholar] [CrossRef]
- Lang, I.K.; Hodac, L.; Friedl, T.; Feussner, I. Fatty acid profiles and their distribution patterns in microalgae: A comprehensive analysis of more than 2000 strains from the SAG culture collection. BMC Plant Biol. 2011, 11, 124–140. [Google Scholar] [CrossRef]
- Klok, A.J.; Lamers, P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. Edible oils from microalgae: Insights in TAG accumulation. Trends Biotechnol. 2014, 32, 521–528. [Google Scholar] [CrossRef]
- Wang, H.; Gao, L.L.; Chen, L.; Guo, F.J.; Liu, T.Z. Integration process of biodiesel production from filamentous oleaginous microalgae Tribonema minus. Bioresour. Technol. 2013, 142, 39–44. [Google Scholar] [CrossRef]
- Guo, F.J.; Wang, H.; Wang, J.F.; Zhou, W.J.; Gao, L.L.; Chen, L.; Dong, Q.Z.; Zhang, W.; Liu, T.Z. Special biochemical responses to nitrogen deprivation of filamentous oleaginous microalgae Tribonema sp. Bioresour. Technol. 2014, 158, 19–24. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, W.J.; Cheng, W.T.; Gao, L.L.; Liu, T.Z. Strategy study on enhancing lipid productivity of filamentous oleaginous microalgae Tribonema. Bioresour. Technol. 2016, 218, 161–166. [Google Scholar]
- Wang, H.; Gao, L.L.; Zhou, W.J.; Liu, T.Z. Growth and palmitoleic acid accumulation of filamentous oleaginous microalgae Tribonema minus at varying temperatures and light regimes. Bioprocess Biosyst. Eng. 2016, 39, 1589–1595. [Google Scholar] [CrossRef]
- Belcher, J.H.; Fogg, G.E. Studies on the growth of Xanthophyceae in pure culture. Arch. Microbiol. 1958, 30, 17–22. [Google Scholar] [CrossRef]
- Zhou, W.J.; Wang, H.; Chen, L.; Cheng, W.T.; Liu, T.Z. Heterotrophy of filamentous oleaginous microalgae Tribonema minus for potential production of lipid and palmitoleic acid. Bioresour. Technol. 2017, 239, 250–257. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, W.J.; Shao, H.M.; Liu, T.Z. A comparative analysis of biomass and lipid content in five Tribonema sp. strains at autotrophic, heterotrophic and mixotrophic cultivation. Algal Res. 2017, 24, 284–289. [Google Scholar] [CrossRef]
- Li, Y.Q.; Mu, J.X.; Chen, D.; Han, F.X.; Xu, H.; Kong, F.; Xie, F.; Feng, B. Production of biomass and lipid by the microalgae Chlorella protothecoides with heterotrophic-Cu(II) stressed (HCuS) coupling cultivation. Bioresour. Technol. 2013, 148, 283–292. [Google Scholar] [CrossRef]
- Li, Y.Q.; Mu, J.X.; Chen, D.; Xu, H.; Han, F.X. Enhanced lipid accumulation and biodiesel production by oleaginous Chlorella protothecoides under a structured heterotrophic-iron (II) induction strategy. World J. Microbiol. Biotechnol. 2015, 31, 773–783. [Google Scholar] [CrossRef]
- Shen, X.F.; Liu, J.J.; Chauhan, A.S.; Hu, H.; Ma, L.L.; Lam, P.K.S.; Zeng, R.J. Combining nitrogen starvation with sufficient phosphorus supply for enhanced biodiesel productivity of Chlorella vulgaris fed on acetate. Algal Res. 2016, 17, 261–267. [Google Scholar] [CrossRef]
- Fu, L.; Cui, X.C.; Li, Y.B.; Xu, L.; Zhang, C.F.; Xiong, R.H.; Zhou, D.D.; Crittenden, J.C. Excessive phosphorus enhances Chlorella regularis lipid production under nitrogen starvation stress during glucose heterotrophic cultivation. Chem. Eng. J. 2017, 330, 566–572. [Google Scholar] [CrossRef]
- Fan, J.H.; Zheng, L.H. Acclimation to NaCl and light stress of heterotrophic Chlamydomonas reinhardtii for lipid accumulation. J. Biosci. Bioeng. 2017, 124, 302–308. [Google Scholar] [CrossRef]
- Li, Y.Q.; Xu, H.; Han, F.X.; Mu, J.X.; Chen, D.; Feng, B.; Zeng, H.Y. Regulation of lipid metabolism in the green microalga Chlorella protothecoides by heterotrophy-photoinduction cultivation regime. Bioresour. Technol. 2015, 192, 781–791. [Google Scholar] [CrossRef]
- Zheng, Y.B.; Chi, Z.Y.; Lucker, B.; Chen, S.L. Two-stage heterotrophic and phototrophic culture strategy for algal biomass and lipid production. Bioresour. Technol. 2012, 103, 484–488. [Google Scholar] [CrossRef]
- Shi, X.M.; Liu, H.J.; Zhang, X.W.; Chen, F. Production of biomass and lutein by Chlorella protothecoides at various glucose concentrations in heterotrophic cultures. Process Biochem. 1999, 34, 341–347. [Google Scholar] [CrossRef]
- Jiang, Y.; Yoshida, T.; Ouigg, A. Photosynthetic performance, lipid production and biomass composition in response to nitrogen limitation in marine microalgae. Plant Physiol. Biochem. 2012, 54, 70–77. [Google Scholar] [CrossRef]
- Li, Y.; Han, D.P.; Hu, Q.; Sommerfeld, M. Inhibition of starch synthesis results in overproduction of lipids in Chlamydomonas reinhardtii. Biotechnol. Bioeng. 2010, 107, 258–268. [Google Scholar] [CrossRef]
- Vince, O.; Wendy, A.; Peter, B.; Johanes van, S.; Csaba, L. Changes in lipid, protein and pigment concentrations in nitrogen-stressed Chlorella minutissima cultures. J. Appl. Phycol. 2012, 24, 907–914. [Google Scholar]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar]
- Liu, T.Z.; Wang, J.F.; Cheng, P.F.; Ji, B.; Liu, J.L.; Chen, Y.; Zhang, W.; Chen, X.L.; Chen, L.; Gao, L.L.; et al. Attached cultivation technology of microalgae for efficient biomass feedstock production. Bioresour. Technol. 2013, 127, 216–222. [Google Scholar] [CrossRef]
- Chen, L.; Liu, T.Z.; Zhang, W.; Chen, X.L.; Wang, J.F. Biodiesel production from algae oil high in free fatty acids by two-step catalytic conversion. Bioresour. Technol. 2012, 111, 208–214. [Google Scholar] [CrossRef]
- Cooksey, K.E.; Guckert, J.B.; Williams, S.A.; Callis, P.R. Fluorometric determination of the neutral lipid content of microalgal cells using Nile Red. J. Microbiol. Methods 1987, 6, 333–345. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
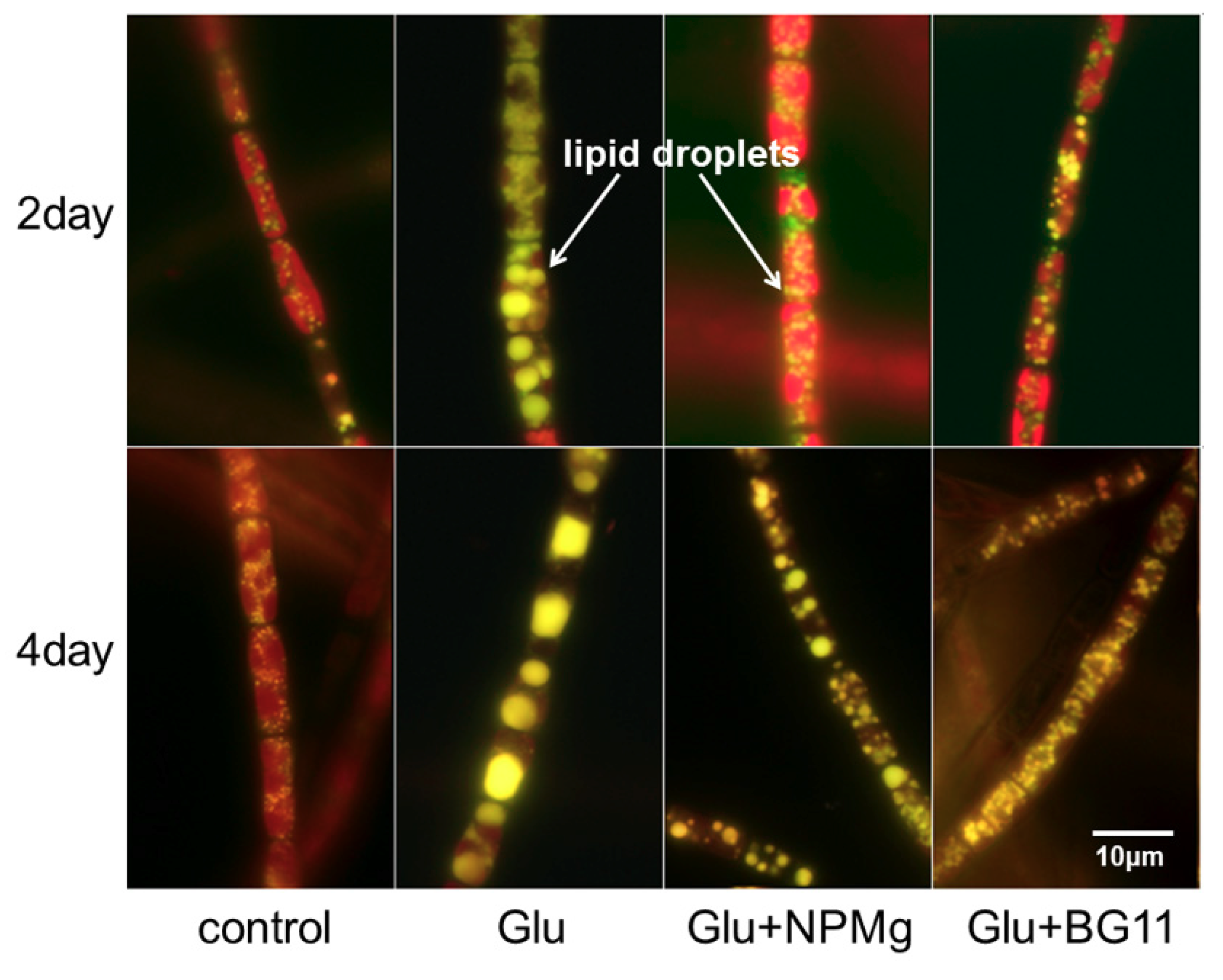{kind=link}
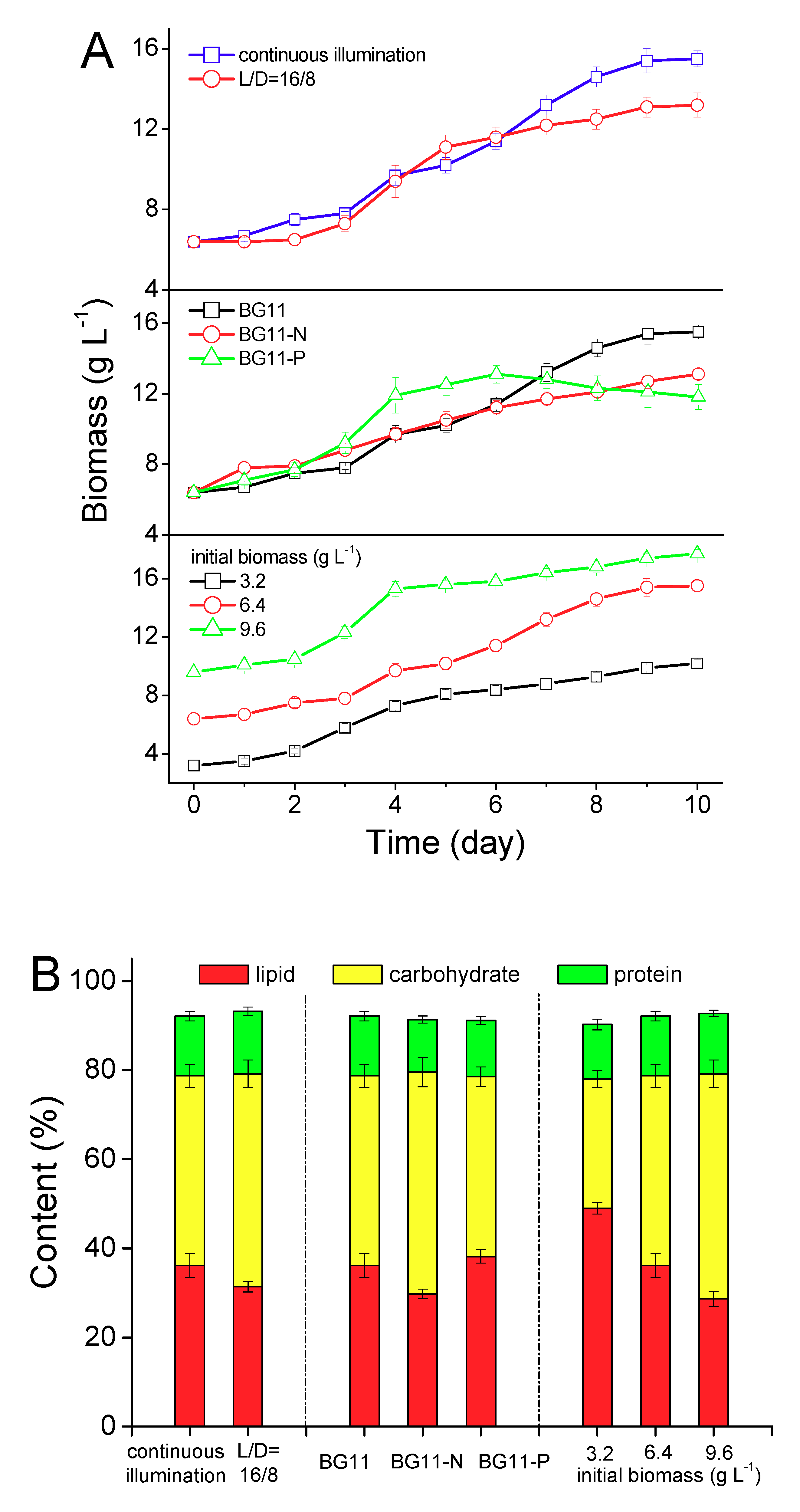{kind=link}
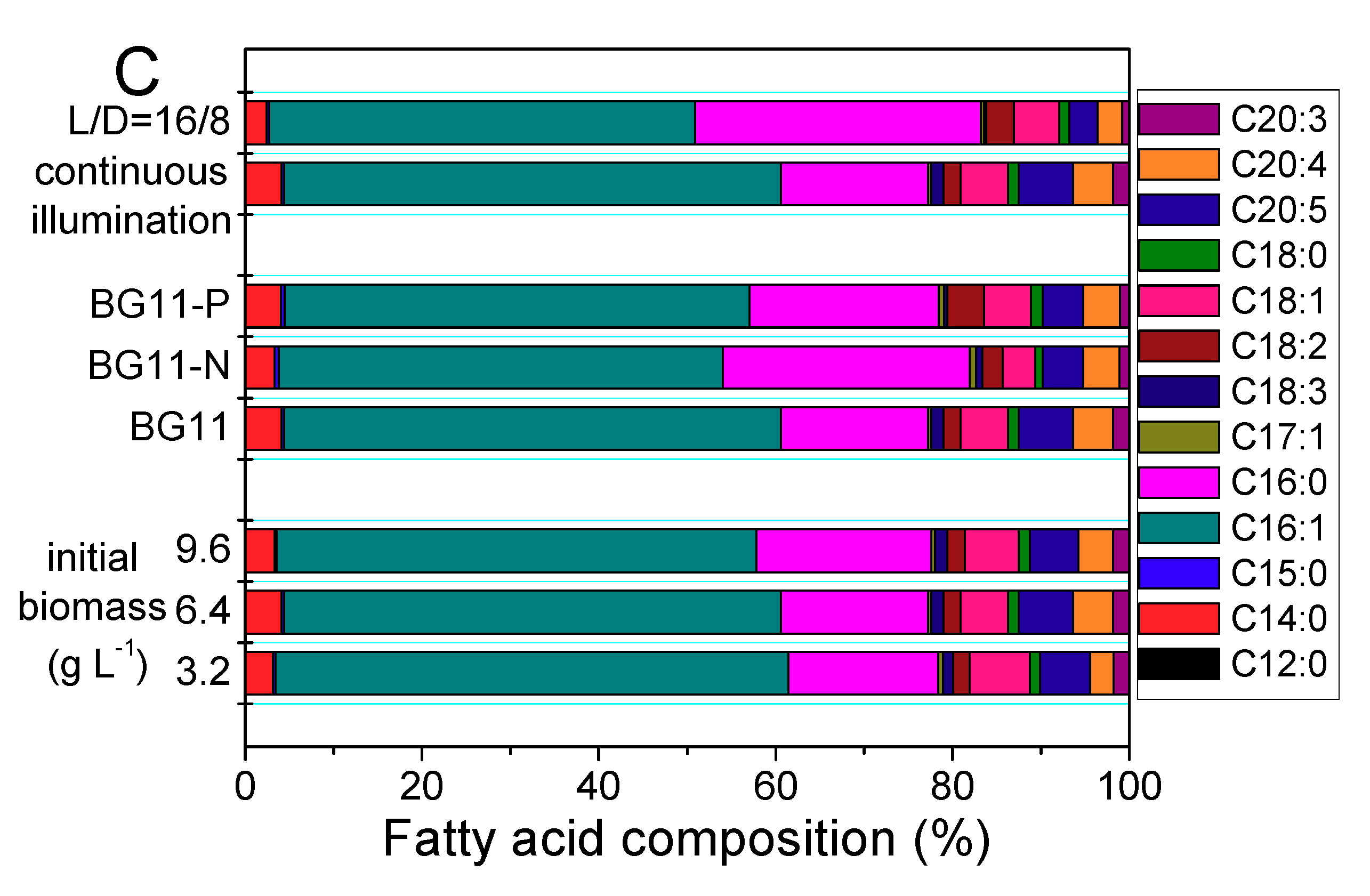{kind=link}
| Induction Methods | Phototrophy | Heterotrophy |
|---|---|---|
| Initial lipid content (% DW) | 15.5 ± 1.4 | 15.5 ± 1.4 |
| Final lipid content (% DW) | 49.0 ± 1.3 | 41.9 ± 2.4 |
| Maximum lipid productivity (g L−1 d−1) | 0.45 | 1.77 |
| Maximum palmitoleic acid content (% fatty acid profile) | 58.0 | 52.2 |
| Maximum palmitoleic acid productivity (mg L−1 d−1) | 261 | 924 |
| Modified BG11 Medium | |||
|---|---|---|---|
| Glucose | 20 g L−1 | ZnSO4·7H2O | 0.222 mg L−1 |
| Urea | 2 g L−1 | CuSO4·5H2O | 0.079 mg L−1 |
| K2HPO4 | 400 mg L−1 | MnCl2·4H2O | 1.81 mg L−1 |
| MgSO4·7H2O | 375 mg L−1 | Na2MoO4·2H2O | 0.39 mg L−1 |
| CaCl2·2H2O | 180 mg L−1 | Co(NO3)4·6H2O | 0.0494 mg L−1 |
| Na2CO3 | 100 mg L−1 | H3BO3 | 2.86 mg L−1 |
| Citric acid | 30 mg L−1 | Na2EDTA | 5 mg L−1 |
| Ammonium ferric citrate | 24 mg L−1 |
| Regimes | Specific Conditions | |
|---|---|---|
| Heterotrophy (air flow rate of 0.5 vvm with pure air, initial biomass of 9.4 g L−1) | Glu medium (adding glucose in water) | |
| Glu+NPMg medium (adding glucose, N, P and Mg in water) | ||
| Glu+BG11 medium (adding glucose in BG11 medium) | ||
| Control (pure water) | ||
| Phototrophy (100 μmol photons m–2 s−1, air flow rate of 0.5 vvm with 1% CO2 in air) | Illumination test (BG11 medium and initial biomass of 6.4 g L−1) | Continuous |
| Alternative (light/dark = 16 h/8 h) | ||
| Nutrient test (continuous illumination and initial biomass of 6.4 g L−1) | BG11 medium | |
| BG11-N medium (N-free) | ||
| BG11-P medium (P-free) | ||
| Initial inoculation biomass test (continuous illumination and BG11 medium) | 3.2 g L−1 | |
| 6.4 g L−1 | ||
| 9.6 g L−1 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, W.; Wang, H.; Zheng, L.; Cheng, W.; Gao, L.; Liu, T. Comparison of Lipid and Palmitoleic Acid Induction of Tribonema minus under Heterotrophic and Phototrophic Regimes by Using High-Density Fermented Seeds. Int. J. Mol. Sci. 2019, 20, 4356. https://doi.org/10.3390/ijms20184356
Zhou W, Wang H, Zheng L, Cheng W, Gao L, Liu T. Comparison of Lipid and Palmitoleic Acid Induction of Tribonema minus under Heterotrophic and Phototrophic Regimes by Using High-Density Fermented Seeds. International Journal of Molecular Sciences. 2019; 20(18):4356. https://doi.org/10.3390/ijms20184356
Chicago/Turabian StyleZhou, Wenjun, Hui Wang, Li Zheng, Wentao Cheng, Lili Gao, and Tianzhong Liu. 2019. "Comparison of Lipid and Palmitoleic Acid Induction of Tribonema minus under Heterotrophic and Phototrophic Regimes by Using High-Density Fermented Seeds" International Journal of Molecular Sciences 20, no. 18: 4356. https://doi.org/10.3390/ijms20184356