Regenerative Effect of Growth Hormone (GH) in the Retina after Kainic Acid Excitotoxic Damage



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Administration of Intraocular Injections
2.3. Quantification of Gene Expression by Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.4. Western Blot Analysis
2.5. Histochemistry and Bromodeoxyuridine (BrdU) Assay
2.6. Statistical Analysis
3. Results
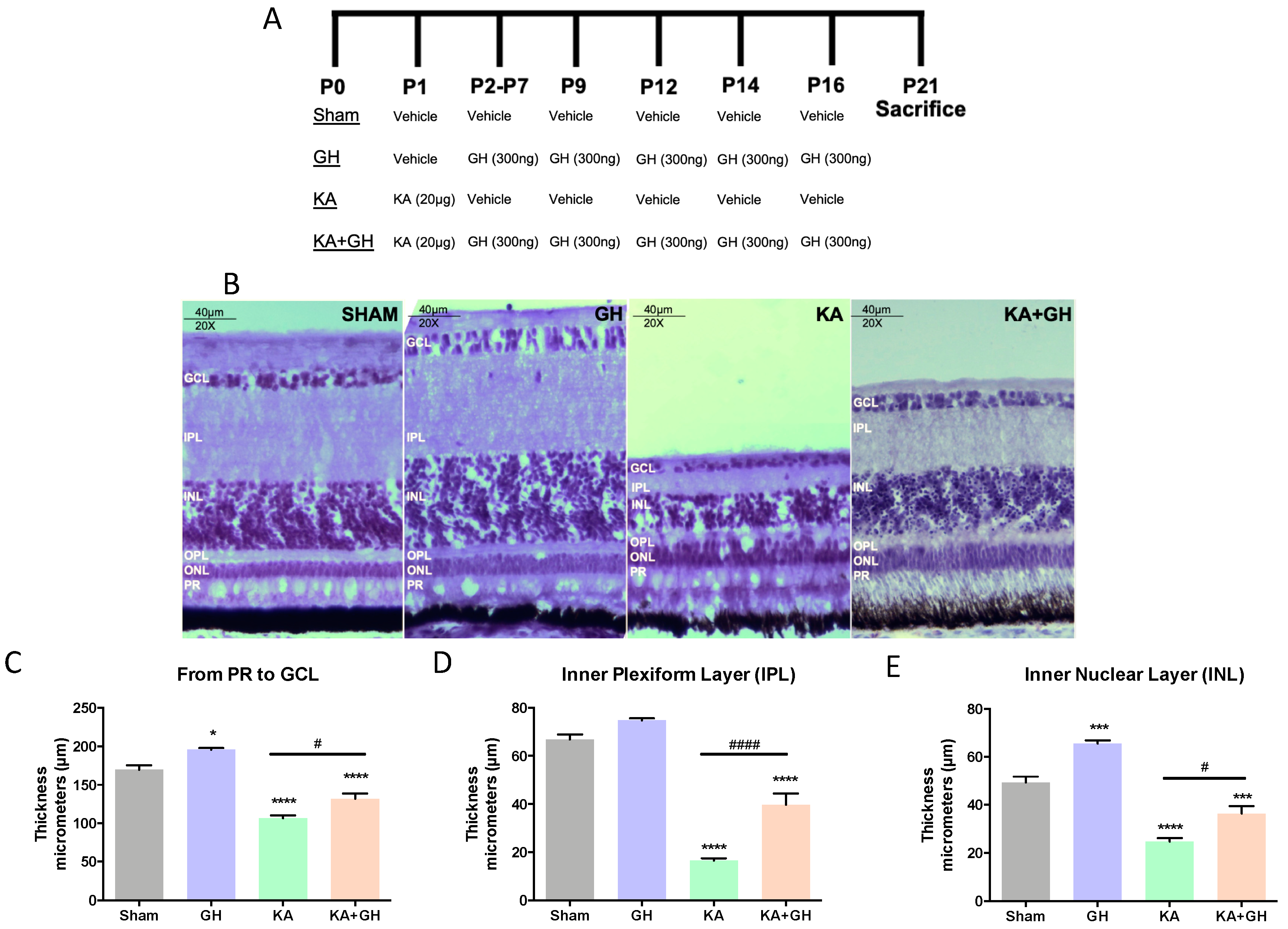
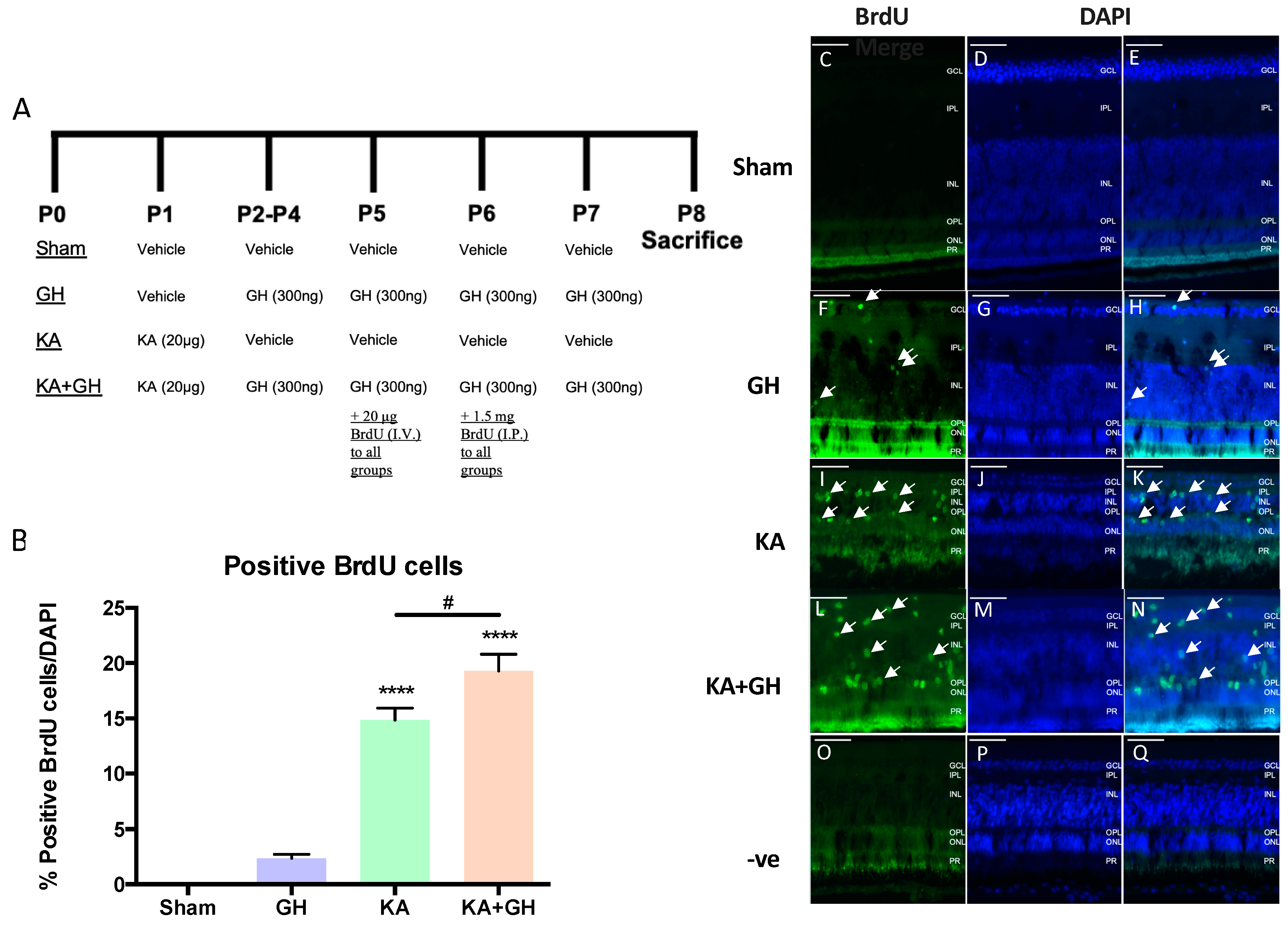
3.1. Effect of Growth Hormone (GH) on Neuroregeneration in KA Damaged Retinas
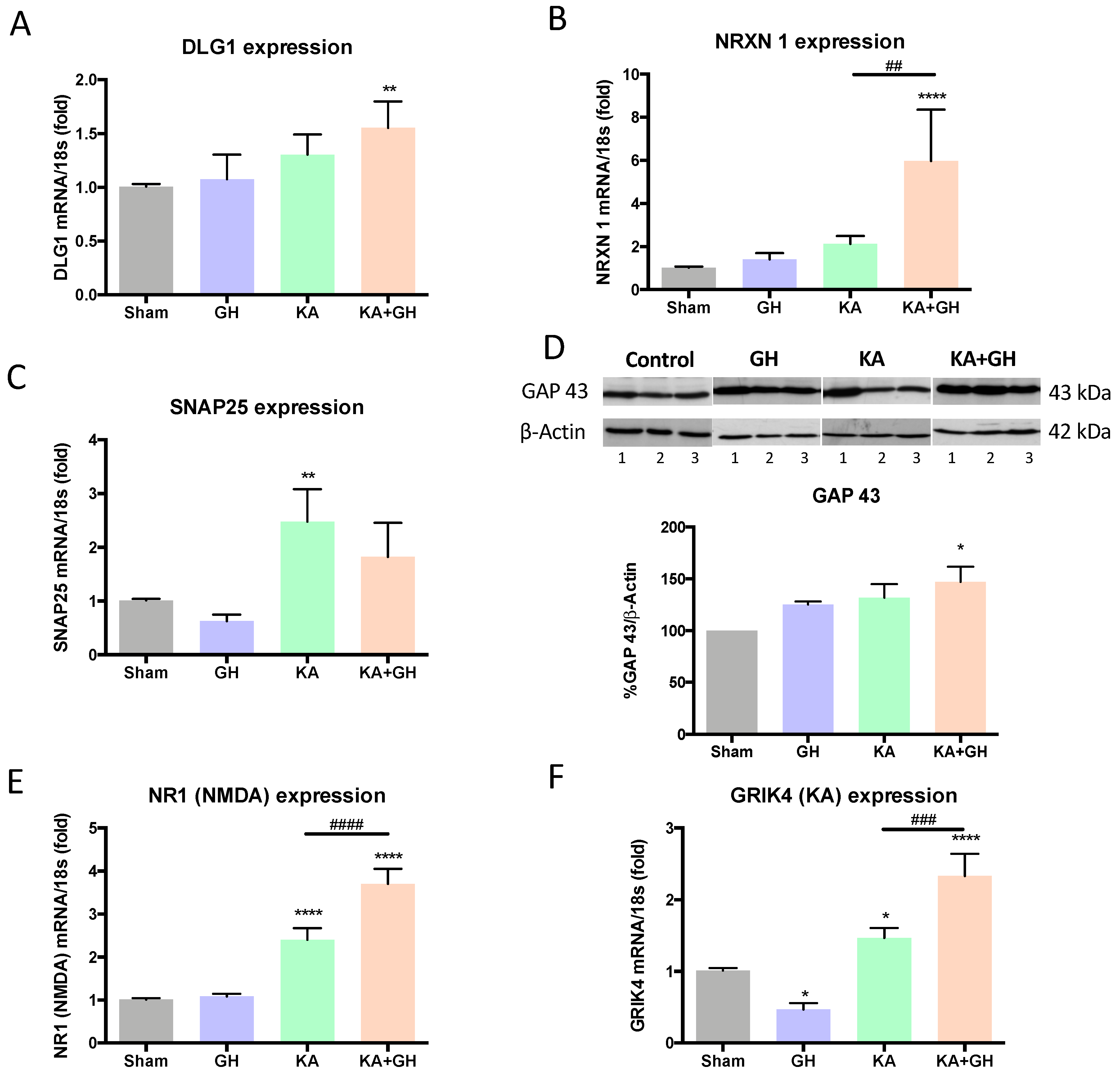
3.2. Growth Hormone (GH) increases Synaptogenic Markers and Glutamate Receptor Expression in Response to Injury
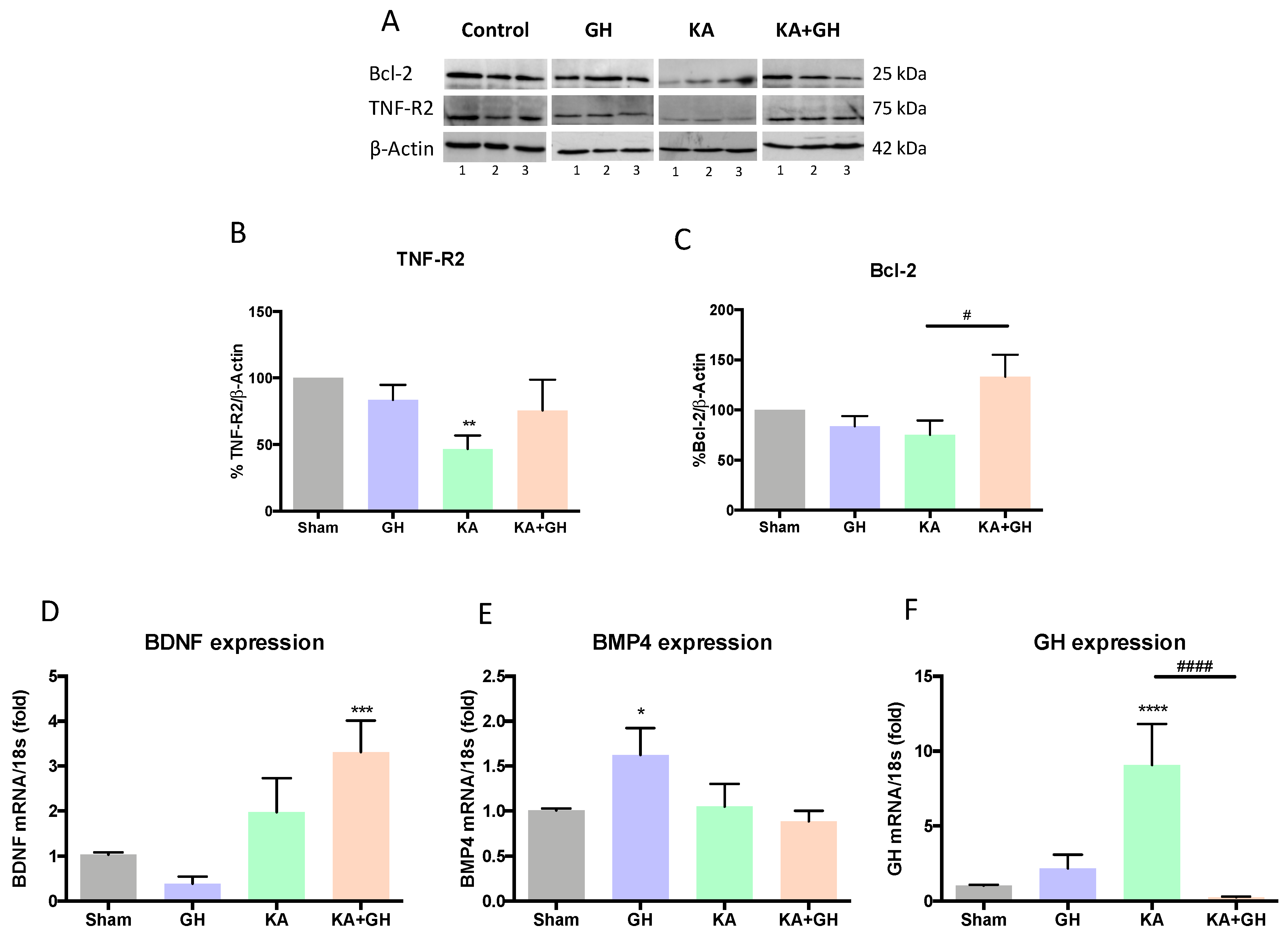
3.3. Pro-survival Effects of Growth Hormone (GH) in the Damaged Retina
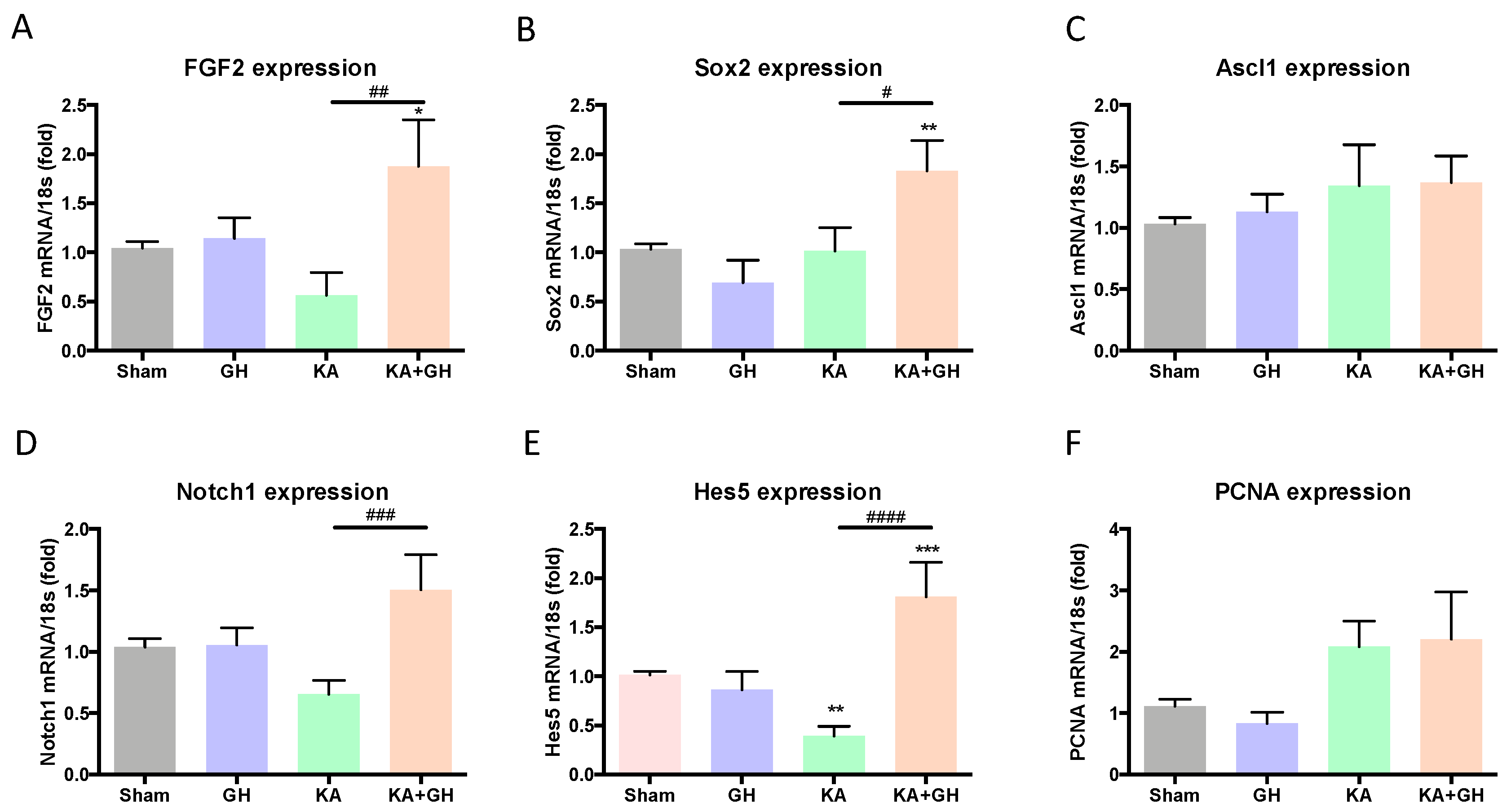
3.4. Effect of Growth Hormone (GH) Treatment on Cell Transdifferentiation and Notch Signaling Pathway
3.5. Proliferative Effect of Growth Hormone (GH) in the Damaged Retina
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Engler, A.; Zhang, R.; Taylor, V. Notch and Neurogenesis. In Molecular Mechanisms of Notch Signaling; Borggrefe, T., Giaimo, B.D., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 223–234. ISBN 978–3-319–89512. [Google Scholar]
- Katsimpardi, L.; Lledo, P.M. Regulation of neurogenesis in the adult and aging brain. Curr. Opin. Neurobiol. 2018, 53, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Zelinka, C.P.; Scott, M.A.; Volkov, L.; Fischer, A.J. The reactivity, distribution and abundance of non-astrocytic inner retinal glial (NIRG) cells are regulated by microglia, acute damage, and IGF1. PLoS ONE 2012, 7, e44477. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.J.; Zelinka, C.; Gallina, D.; Scott, M.A.; Todd, L. Reactive microglia and macrophage facilitate the formation of Müller glia-derived retinal progenitors. Glia 2014, 62, 1608–1628. [Google Scholar] [CrossRef] [PubMed]
- Ramon-Cañellas, P.; Peterson, H.P.; Morante, J. From early to late neurogenesis: Neural progenitors and the glial niche from a fly’s point of view. Neuroscience 2019, 399, 39–52. [Google Scholar] [CrossRef]
- Bianchi, V.E.; Locatelli, V.; Rizzi, L. Neurotrophic and neuroregenerative effects of GH/IGF1. Int. J. Mol. Sci. 2017, 18, 2441. [Google Scholar] [CrossRef]
- Martinez-Moreno, C.G.; Fleming, T.; Carranza, M.; Ávila-Mendoza, J.; Luna, M.; Harvey, S.; Arámburo, C. Growth hormone protects against kainate excitotoxicity and induces BDNF and NT3 expression in chicken neuroretinal cells. Exp. Eye Res. 2018, 166, 1–12. [Google Scholar] [CrossRef]
- Cuatrecasas, G.; Kumru, H.; Coves, M.J.; Vidal, J. GH deficiency in patients with spinal cord injury: Efficacy/safety of GH replacement, a pilot study. Endocr. Connect. 2018, 7, 1031–1039. [Google Scholar] [CrossRef]
- Butler, T.; Harvey, P.; Cardozo, L.; Zhu, Y.S.; Mosa, A.; Tanzi, E.; Pervez, F. Epilepsy, depression, and growth hormone. Epilepsy Behav. 2019, 94, 297–300. [Google Scholar] [CrossRef]
- Fischer, A.J.; Bongini, R. Turning Müller glia into neural progenitors in the retina. Mol. Neurobiol. 2010, 42, 199–209. [Google Scholar] [CrossRef]
- Hamon, A.; Roger, J.E.; Yang, X.J.; Perron, M. Müller glial cell-dependent regeneration of the neural retina: An overview across vertebrate model systems. Dev. Dyn. 2016, 245, 727–738. [Google Scholar] [CrossRef]
- Agathocleous, M.; Harris, W.A. From progenitors to differentiated cells in the vertebrate retina. Annu. Rev. Cell Dev. Biol. 2009, 25, 45–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, C.; Su, G. Cellular signaling in Müller glia: Progenitor cells for regenerative and neuroprotective responses in pharmacological models of retinal degeneration. J. Ophthalmol. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gallina, D.; Todd, L.; Fischer, A.J. A comparative analysis of Müller glia-mediated regeneration in the vertebrate retina. Exp. Eye Res. 2014, 123, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Todd, L.; Palazzo, I.; Squires, N.; Mendonca, N.; Fischer, A.J. BMP- and TGFβ-signaling regulate the formation of Müller glia-derived progenitor cells in the avian retina. Glia 2017, 65, 1640–1655. [Google Scholar] [CrossRef] [PubMed]
- Baudet, M.L.; Rattray, D.; Martin, B.T.; Harvey, S. Growth hormone promotes axon growth in the developing nervous system. Endocrinology 2009, 150, 2758–2766. [Google Scholar] [CrossRef] [PubMed]
- Harvey, S.; Martínez-Moreno, C.G.; Ávila-Mendoza, J.; Luna, M.; Arámburo, C. Growth hormone in the eye: A comparative update. Gen. Comp. Endocrinol. 2016, 234, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Moreno, C.; Andres, A.; Giterman, D.; Karpinski, E.; Harvey, S. Growth hormone and retinal ganglion cell function: QNR/D cells as an experimental model. Gen. Comp. Endocrinol. 2014, 195, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.; Martínez-Moreno, C.G.; Mora, J.; Aizouki, M.; Luna, M.; Arámburo, C.; Harvey, S. Internalization and synaptogenic effect of GH in retinal ganglion cells (RGCs). Gen. Comp. Endocrinol. 2016, 234, 151–160. [Google Scholar] [CrossRef]
- Fleming, T.; Martinez-Moreno, C.G.; Carranza, M.; Luna, M.; Harvey, S.; Arámburo, C. Growth hormone promotes synaptogenesis and protects neuroretinal dendrites against kainic acid (KA) induced damage. Gen. Comp. Endocrinol. 2018, 265, 111–120. [Google Scholar] [CrossRef]
- Martínez-Moreno, C.G.; Ávila-Mendoza, J.; Wu, Y.; Arellanes-Licea Edel, C.; Louie, M.; Luna, M.; Arámburo, C.; Harvey, S. Neuroprotection by GH against excitotoxic-induced cell death in retinal ganglion cells. Gen. Comp. Endocrinol. 2016, 234, 68–80. [Google Scholar] [CrossRef]
- Kermer, P.; Klöcker, N.; Labes, M.; Bähr, M. Insulin-like growth factor-I protects axotomized rat retinal ganglion cells from secondary death via PI3-K-dependent Akt phosphorylation and inhibition of caspase-3 in vivo. J. Neurosci. 2018, 20, 722–728. [Google Scholar] [CrossRef]
- Sanders, E.J.; Baudet, M.L.; Parker, E.; Harvey, S. Signaling mechanisms mediating local GH action in the neural retina of the chick embryo. Gen. Comp. Endocrinol. 2009, 163, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Le Grevès, M.; Zhou, Q.; Berg, M.; Le Grevès, P.; Fhölenhag, K.; Meyerson, B.; Nyberg, F. Growth hormone replacement in hypophysectomized rats affects spatial performance and hippocampal levels of NMDA receptor subunit and PSD-95 gene transcript levels. Exp. Brain Res. 2006, 173, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Nag, T.C.; Sharma, U.; Jagannathan, N.R.; Wadhwa, S. Differential effects of prenatal chronic high-decibel noise and music exposure on the excitatory and inhibitory synaptic components of the auditory cortex analog in developing chicks (Gallus gallus domesticus). Neuroscience 2014, 269, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Uchigashima, M.; Cheung, A.; Suh, J.; Watanabe, M.; Futai, K. Differential expression of neurexin genes in the mouse brain. J. Comp. Neurol. 2019, 1940–1965. [Google Scholar] [CrossRef] [PubMed]
- Paraoanu, L.E.; Becker-Roeck, M.; Christ, E.; Layer, P.G. Expression patterns of neurexin-1 and neuroligins in brain and retina of the chick embryo: Neuroligin-3 is absent in retina. Neurosci. Lett. 2006, 395, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, G.W.; Wang, J.; Brin, M.F.; Aoki, K.R.; Wheeler, L.; Dolly, J.O. Fusion of Golgi-derived vesicles mediated by SNAP-25 is essential for sympathetic neuron outgrowth but relatively insensitive to botulinum neurotoxins in vitro. FEBS J. 2014, 281, 3243–3260. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yang, W.; Xiao, T.; Miao, Z.; Luo, W.; You, Z.; Li, G. Possible role of miR-204 in optic nerve injury through the regulation of GAP-43. Mol. Med. Rep. 2018, 17, 3891–3897. [Google Scholar] [CrossRef] [PubMed]
- Leon, S.; Yin, Y.; Nguyen, J.; Irwin, N.; Benowitz, L.I. Lens Injury Stimulates Axon Regeneration in the Mature Rat Optic Nerve. J. Neurosci. 2000, 20, 4615–4626. [Google Scholar] [CrossRef] [Green Version]
- Ramis, M.; Sarubbo, F.; Sola, J.; Aparicio, S.; Garau, C.; Miralles, A.; Esteban, S. Cognitive improvement by acute growth hormone is mediated by NMDA and AMPA receptors and MEK pathway. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2013, 45, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Nylander, E.; Grönbladh, A.; Zelleroth, S.; Diwakarla, S.; Nyberg, F.; Hallberg, M. Growth hormone is protective against acute methadone-induced toxicity by modulating the NMDA receptor complex. Neuroscience 2016, 339, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Elias, E.; Yang, N.; Wang, P.; Tian, N. Glutamate activity regulates and dendritic development of J-RGCs. Front. Cell. Neurosci. 2018, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- McAllister, A.K. Cellular and molecular mechanisms of dendrite growth. Cereb. Cortex 2000, 10, 963–973. [Google Scholar] [CrossRef]
- Lee, J.G.; Lee, K.P.; Jeon, C.J. Synaptic pattern of KA1 and KA2 upon the direction-selective ganglion cells in developing and adult mouse retina. Acta Histochem. Cytochem. 2012, 45, 35–45. [Google Scholar] [CrossRef]
- Wajant, H.; Siegmund, D. TNFR1 and TNFR2 in the control of the life and death balance of macrophages. Front. Cell Dev. Biol. 2019, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, V.; Mohand-Said, S.; Hanoteau, N.; Fuchs, C.; Pfizenmaier, K.; Eisel, U. Neurodegenerative and neuroprotective effects of tumor necrosis factor (TNF) in retinal ischemia: Opposite roles of TNF receptor 1 and TNF receptor 2. J. Neurosci. 2002, 22, RC216. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, T.; Nakazawa, C.; Matsubara, A.; Noda, K.; Hisatomi, T.; She, H.; Michaud, N.; Hafezi-Moghadam, A.; Miller, J.W.; Benowitz, L.I. Tumor necrosis factor-alpha mediates oligodendrocyte death and delayed retinal ganglion cell loss in a mouse model of glaucoma. J. Neurosci. 2006, 26, 12633–12641. [Google Scholar] [CrossRef] [PubMed]
- Alba-Betancourt, C.; Luna-Acosta, J.L.; Ramírez-Martínez, C.E.; Ávila-González, D.; Granados-Ávalos, E.; Carranza, M.; Martínez-Coria, H.; Arámburo, C.; Luna, M. Neuro-protective effects of growth hormone (GH) after hypoxia-ischemia injury in embryonic chicken cerebellum. Gen. Comp. Endocrinol. 2013, 183, 17–31. [Google Scholar] [CrossRef]
- Luna-Acosta, J.L.; Alba-Betancourt, C.; Martínez-Moreno, C.G.; Ramírez, C.; Carranza, M.; Luna, M.; Arámburo, C. Direct antiapoptotic effects of growth hormone are mediated by PI3K/Akt pathway in the chicken bursa of Fabricius. Gen. Comp. Endocrinol. 2015, 224, 148–159. [Google Scholar] [CrossRef]
- Mysona, B.A.; Zhao, J.; Bollinger, K.E. Role of BDNF/TrkB pathway in the visual system: Therapeutic implications for glaucoma. Expert Rev. Ophthalmol. 2017, 12, 69–81. [Google Scholar] [CrossRef]
- Fischer, A.J.; Schmidt, M.; Omar, G.; Reh, T.A. BMP4 and CNTF are neuroprotective and suppress damage-induced proliferation of Müller glia in the retina. Mol. Cell. Neurosci. 2004, 27, 531–542. [Google Scholar] [CrossRef]
- Todd, L.; Squires, N.; Suarez, L.; Fischer, A.J. Jak/Stat signaling regulates the proliferation and neurogenic potential of Müller glia-derived progenitor cells in the avian retina. Sci. Rep. 2016, 6, 35703. [Google Scholar] [CrossRef] [PubMed]
- Seydewitz, V.; Rothermel, A.; Fuhrmann, S.; Schneider, A.; DeGrip, W.J.; Layer, P.G.; Hofmann, H.D. Expression of CNTF receptor-alpha in chick violet-sensitive cones with unique morphologic properties. Investig. Ophthalmol. Vis. Sci. 2004, 45, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.J.; McGuire, C.R.; Dierks, B.D.; Reh, T.A. Insulin and fibroblast growth factor 2 activate a neurogenic program in Müller glia of the chicken retina. J. Neurosci. 2002, 22, 9387–9398. [Google Scholar] [CrossRef] [PubMed]
- Zelinka, C.P.; Volkov, L.; Goodman, Z.A.; Todd, L.; Palazzo, I.; Bishop, W.A.; Fischer, A.J. mTor signaling is required for the formation of proliferating Müller glia-derived progenitor cells in the chick retina. Development 2016, 143, 1859–1873. [Google Scholar] [CrossRef] [PubMed]
- Elsaeidi, F.; Macpherson, P.; Mills, E.A.; Jui, J.; Flannery, J.G.; Goldman, D. Notch suppression collaborates with Ascl1 and Lin28 to unleash a regenerative response in fish retina, but not in mice. J. Neurosci. 2018, 38, 2246–2261. [Google Scholar] [CrossRef] [PubMed]
- Wilken, M.S.; Reh, T.A. Retinal regeneration in birds and mice. Curr. Opin. Genet. Dev. 2016, 40, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Ghai, K.; Zelinka, C.; Fischer, A.J. Notch signaling influences neuroprotective and proliferative properties of mature Müller glia. J. Neurosci. 2010, 30, 3101–3112. [Google Scholar] [CrossRef]
- Hayes, S.; Nelson, B.R.; Buckingham, B.; Reh, T.A. Notch signaling regulates regeneration in the avian retina. Dev. Biol. 2007, 312, 300–311. [Google Scholar] [CrossRef] [Green Version]
- Mills, E.A.; Goldman, D. The regulation of Notch signaling in retinal development and regeneration. Curr. Pathobiol. Rep. 2017, 5, 323–331. [Google Scholar] [CrossRef]
- Li, L.; Chen, L.; Liu, Q. Effect of the Notch signaling pathway on retinal ganglion cells and its neuroprotection in rats with acute ocular hypertension. Int. J. Ophthalmol. 2018, 11, 208–215. [Google Scholar] [PubMed]
- Fleming, T.; Balderas-Márquez, J.E.; Epardo, D.; Ávila-Mendoza, J.; Carranza, M.; Luna, M.; Harvey, S.; Arámburo, C.; Martínez-Moreno, C.G. GH neuroprotection against kainate (KA) excitotoxicity in the retina is mediated by Notch/PTEN/Akt signaling. Investig. Ophthalmol. Vis. Sci. 2019. submitted for publication. [Google Scholar]
- Fischer, A.J.; Reh, T.A. Identification of a proliferating marginal zone of retinal progenitors in postnatal chickens. Dev. Biol. 2000, 220, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Boehm, E.M.; Gildenberg, M.S.; Washington, M.T. The Many Roles of PCNA in Eukaryotic DNA Replication. Enzymes 2016, 39, 231–254. [Google Scholar] [PubMed] [Green Version]
- Fischer, A.J.; Reh, T.A. Exogenous growth factors stimulate the regeneration of ganglion cells in the chicken retina. Dev. Biol. 2002, 251, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.J.; Reh, T.A. Growth factors induce neurogenesis in the ciliary body. Dev. Biol. 2003, 259, 225–240. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method. Methods 2001, 408, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Kuchipudi, S.V.; Tellabati, M.; Nelli, R.K.; White, G.A.; Perez, B.B.; Sebastian, S.; Slomka, M.J.; Brookes, S.M.; Brown, I.H.; Dunham, S.P.; et al. 18S rRNA is a reliable normalisation gene for real time PCR based on influenza virus infected cells. Virol. J. 2012, 9, 1–7. [Google Scholar] [CrossRef]
- Bas, A.; Forsberg, G.; Hammarström, S.; Hammarström, M.L. Utility of the housekeeping genes 18S rRNA, β-actin and glyceraldehyde-3-phosphate-dehydrogenase for normalization in real-time quantitative reverse transcriptase-polymerase chain reaction analysis of gene expression in human T lymphocytes. Scand. J. Immunol. 2004, 59, 566–573. [Google Scholar] [CrossRef]
- Fischer, A.J.; Reh, T.A. Müller glia are a potential source of neural regeneration in the postnatal chicken retina. Nat. Neurosci. 2001, 4, 247–252. [Google Scholar] [CrossRef]
- Martínez-Moreno, C.G.; Calderón-Vallejo, D.; Harvey, S.; Arámburo, C.; Quintanar, J.L. Growth hormone (GH) and gonadotropin-releasing hormone (GnRH) in the central nervous system: A potential neurological combinatory therapy? Int. J. Mol. Sci. 2018, 19, 375. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Gan, L. Development of retinal amacrine cells and their dendritic stratification. Curr. Ophthalmol. Rep. 2014, 2, 100–106. [Google Scholar] [CrossRef]
- Grimbly, C.; Martin, B.; Karpinski, E.; Harvey, S. Growth hormone production and action in N1E-115 neuroblastoma cells. J. Mol. Neurosci. 2009, 39, 117–124. [Google Scholar] [CrossRef]
- Devesa, J.; Núñez, I.; Agra, C.; Bejarano, A.; Devesa, P. Treatment with growth hormone (GH) increased the metabolic activity of the brain in an elder patient, not GH-deficient, who suffered mild cognitive alterations and had an ApoE 4/3 genotype. Int. J. Mol. Sci. 2018, 19, 2294. [Google Scholar] [CrossRef]
- Nyberg, F.; Hallberg, M. Growth hormone and cognitive function. Nat. Rev. Endocrinol. 2013, 9, 357. [Google Scholar] [CrossRef]
- Martín-Rodríguez, J.F.; Ramos-Herrero, V.D.; Parras, G.G.; Flores-Martínez, Á.; Madrazo-Atutxa, A.; Cano, D.A.; Gruart, A.; Delgado-García, J.M.; Leal-Cerro, A.; Leal-Campanario, R. Chronic adult-onset of growth hormone/IGF-I hypersecretion improves cognitive functions and LTP and promotes neuronal differentiation in adult rats. Acta Physiol. 2019, e13293. [Google Scholar] [CrossRef]
- Hull, K.; Harvey, S. Growth hormone therapy and quality of life: Possibilities, pitfalls and mechanisms. J. Endocrinol. 2004, 179, 311–333. [Google Scholar] [CrossRef]
- Daly, C.; Ward, R.; Reynolds, A.L.; Galvin, O.; Collery, R.F.; Kennedy, B.N. Brain-derived neurotrophic factor as a treatment option for retinal degeneration. In Proceedings of the Retinal Degenerative Diseases; Ash, J.D., Anderson, R.E., LaVail, M.M., Bowes Rickman, C., Hollyfield, J.G., Grimm, C., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 465–471. ISBN 978–3-319–75402-4. [Google Scholar]
- Li, G.; Berven, S.; Simpson, H.; Triffitt, J.T. Expression of BMP-4 mRNA during distraction osteogenesis in rabbits. Acta Orthop. Scand. 1998, 69, 420–425. [Google Scholar] [CrossRef]
- Haase, H.R.; Ivanovski, S.; Waters, M.J.; Bartold, P.M. Growth hormone regulates osteogenic marker mRNA expression in human periodontal fibroblasts and alveolar bone-derived cells. J. Periodontal Res. 2003, 38, 366–374. [Google Scholar] [CrossRef]
- McLenachan, S.; Lum, M.G.; Waters, M.J.; Turnley, A.M. Growth hormone promotes proliferation of adult neurosphere cultures. Growth Horm. IGF Res. 2009, 19, 212–218. [Google Scholar] [CrossRef]
- Blackmore, D.G.; Reynolds, B.A.; Golmohammadi, M.G.; Large, B.; Aguilar, R.M.; Haro, L.; Waters, M.J.; Rietze, R.L. Growth hormone responsive neural precursor cells reside within the adult mammalian brain. Sci. Rep. 2012, 2, 1–10. [Google Scholar] [CrossRef]
- Regalado-Santiago, C.; López-Meraz, M.L.; Santiago-García, J.; Fernández-Pomares, C.; Juárez-Aguilar, E. Growth hormone (GH) is a survival rather than a proliferative factor for embryonic striatal neural precursor cells. Growth Horm. IGF Res. 2013, 23, 179–186. [Google Scholar] [CrossRef]
- Devesa, P.; Agasse, F.; Xapelli, S.; Almengló, C.; Devesa, J.; Malva, J.O.; Arce, V.M. Growth hormone pathways signaling for cell proliferation and survival in hippocampal neural precursors from postnatal mice. BMC Neurosci. 2014, 15, 100. [Google Scholar] [CrossRef]
- Ávila-Mendoza, J.; Mora, J.; Carranza, M.; Luna, M.; Arámburo, C. Growth hormone reverses excitotoxic damage induced by kainic acid in the green iguana neuroretina. Gen. Comp. Endocrinol. 2016, 234, 57–67. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Retinal Marker | References |
|---|---|---|---|
| Discs large homolog 1 (DLG1) | Post-synaptic scaffold protein (avian PSD95) | Synaptogenic | [24,25] |
| Neurexin 1 (NRXN1) | Synaptic cell-surface protein | Synaptogenic | [26,27] |
| Synaptosomal nerve-associated protein 25 (SNAP25) | Synaptic vesicle fusion protein | Synaptogenic | [20,28] |
| Growth associated protein 43 (GAP43) | Expressed during axonal growth regulation | Axogenic | [29,30] |
| NR1 | NMDA-R subunit | Synaptogenic | [31,32,33,34] |
| GRIK4 | KA-R subunit | Synaptogenic | [33,35] |
| Tumor necrosis factor receptor 2 (TNF-R2) | Pro-survival receptor | Neurotrophic | [36,37,38] |
| B-cell lymphoma 2 (Bcl-2) | Anti-apoptotic protein | Anti-apoptotic | [21,39,40] |
| Brain-derived neurotrophic factor (BDNF) | Neurotrophic factor | Neurotrophic | [7,41] |
| Bone morphogenic protein 4 (BMP4) | Pro-survival growth factor | Neurotrophic | [42,43,44] |
| Fibroblast growth factor (FGF2) | Proliferative growth factor | Proliferation and transdifferentiation | [13,45] |
| Sex determining Region Y-box 2 (Sox2) | Pluripotency transcription factor | Transdifferentiation | [14,46] |
| Achaete-scute homolog 1 (Ascl1) | Transcription factor associated to differentiation | Transdifferentiation | [47,48] |
| Notch Receptor 1 (Notch1) | Membrane receptor—Notch signaling element | Transdifferentiation | [47,49,50,51,52,53] |
| Hes5 | Responsive gene—Notch signaling pathway | Transdifferentiation | [49,50,51,52,53] |
| Proliferating cell nuclear antigen (PCNA) | Nuclear proliferative protein | Proliferation | [54,55] |
| Target | Primer | Sequence (5′-3′) | Size | Accesion # |
|---|---|---|---|---|
| cNotch1 | Fwd | GGCTGGTTATCATGGAGTTA | 155 bp | NM_001030295.1 |
| Rev | CATCCACATTGATCTCACAG | |||
| cHes5 | Fwd | GGAGAAGGAGTTCCAGAGAC | 171 bp | NM_001012695.1 |
| Rev | ATTTGCAGAGCTTCTTTGAG | |||
| cBDNF | Fwd | AGCAGTCAAGTGCCTTTGGA | 167 bp | NM_001031616 |
| Rev | TCCGCTGCTGTTACCCACTCG | |||
| cASCL | Fwd | AGGGAACCACGTTTATGCAG | 188 bp | NM_204412.1 |
| Rev | TTATACAGGGCCTGGTGAGC | |||
| cSox2 | Fwd | AGGCTATGGGATGATGCAAG | 163 bp | NM_205188.2 |
| Rev | GTAGGTAGGCGATCCGTTCA | |||
| cFGF2 | Fwd | TGCAGCTTCAAGCAGAAGAA | 173 bp | NM_205433.1 |
| Rev | CTTCCGTGACCGGTAAGTGT | |||
| cDLG1 | Fwd | ACCAGCCAGAAGAGATCCCT | 162 bp | XM_025153616.1 |
| Rev | TGGAGTTACCTGCCGTGCTT | |||
| cBMP4 | Fwd | CGCTGGGAGACCTTTGATGT | 153 bp | NM_205237.3 |
| Rev | CCCCTGAGGTAAAGATCGGC | |||
| cSNAP25 | Fwd | GCCTGCCCGTGTGGTAGAT | 185 bp | NM_205458.1 |
| Rev | TCTGGCGATTCTGTGTGTCG | |||
| cNRXN1 | Fwd | CCACTCTGATCATTGACCGGG | 392 bp | NM_001198975.1 |
| Rev | CGCCAGACCTTCCACATAGT | |||
| c18 s | Fwd | CTCTTTCTCGATTCCGTGGGT | 100 bp | M59389 |
| Rev | TTAGCATGCCAGAGTCTCGT | |||
| cNR1 | Fwd | ACGGTCCCACCATACTCTCA | 156 bp | NM_206979.1 |
| Rev | AGCCTTGGACTCTCTCTCCT | |||
| cGH | Fwd | CGCACCTATATTCCGGAGGAC | 128 bp | NM_204359 |
| Rev | GGCAGCTCCATGTCTGACT | |||
| cGRIK4 | Fwd | AAATGTCCCAAGGAGGAGGACCAC | 215 bp | XM_015298177.2 |
| Rev | AAACTGTCCTTGCAGAGGATGATG TTGCGC |
| Target | Host/Type | Dilution | Source | Cat.No |
|---|---|---|---|---|
| Bcl-2 | mouse/polyclonal | 1:1000 | Invitrogen | 138800 |
| β-Actin | mouse/monoclonal | 1:5000 | Santa cruz | SC-47778 |
| TNF-R2 | goat/polyclonal | 1:1000 | Santa Cruz | SC-1072 |
| GAP 43 | Mouse/monoclonal | 1:2000 | Sigma-Aldrich | GAP-7B10 |
| BrdU | Rat/monoclonal | 1:100 | Abcam | AB-6326 |
| Mouse IgG | Goat/HRP conjugated | 1:6000 | Abcam | AB-20043 |
| Rat IgG | Goat/ Alexa fluor 488 | 1:1000 | Abcam | AB-150157 |
| Goat IgG | Rabbit/HRP | 1:6000 | Life tech. | 811620 |
| Response per Treatment | |||||
|---|---|---|---|---|---|
| GH vs Sham | KA vs Sham | KA + GH vs Sham | KA + GH vs KA | ||
| Synaptogenic | DLG1 | - | - | ↑↑ | - |
| NRXN1 | - | - | ↑↑↑↑ | ↑↑ | |
| SNAP25 | - | ↑↑ | - | - | |
| GAP43 | - | - | ↑ | - | |
| NR1 | - | ↑↑↑↑ | ↑↑↑↑ | ↑↑↑↑ | |
| GRIK4 | ↓ | ↑ | ↑↑↑↑ | ↑↑↑ | |
| Survival | TNF-R2 | - | ↓↓ | - | - |
| Bcl-2 | - | - | - | ↑ | |
| BDNF | - | - | ↑↑↑ | - | |
| BMP4 | ↑ | - | - | - | |
| Transdifferentiation | FGF2 | - | - | ↑ | ↑↑ |
| Sox2 | - | - | ↑↑ | ↑ | |
| Ascl1 | - | - | - | - | |
| Notch1 | - | - | - | ↑↑↑ | |
| Hes5 | - | ↓↓ | ↑↑↑ | ↑↑↑↑ | |
| PCNA | - | - | - | - | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Moreno, C.G.; Epardo, D.; Balderas-Márquez, J.E.; Fleming, T.; Carranza, M.; Luna, M.; Harvey, S.; Arámburo, C. Regenerative Effect of Growth Hormone (GH) in the Retina after Kainic Acid Excitotoxic Damage. Int. J. Mol. Sci. 2019, 20, 4433. https://doi.org/10.3390/ijms20184433
Martinez-Moreno CG, Epardo D, Balderas-Márquez JE, Fleming T, Carranza M, Luna M, Harvey S, Arámburo C. Regenerative Effect of Growth Hormone (GH) in the Retina after Kainic Acid Excitotoxic Damage. International Journal of Molecular Sciences. 2019; 20(18):4433. https://doi.org/10.3390/ijms20184433
Chicago/Turabian StyleMartinez-Moreno, Carlos G., David Epardo, Jerusa E. Balderas-Márquez, Thomas Fleming, Martha Carranza, Maricela Luna, Steve Harvey, and Carlos Arámburo. 2019. "Regenerative Effect of Growth Hormone (GH) in the Retina after Kainic Acid Excitotoxic Damage" International Journal of Molecular Sciences 20, no. 18: 4433. https://doi.org/10.3390/ijms20184433