Sulfasalazine as an Immunomodulator of the Inflammatory Process during HIV-1 Infection

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Effect of SSZ on Cell Viability
2.2. SSZ Decreases the Production of IL-1β in Response to Inflammasome Inducers
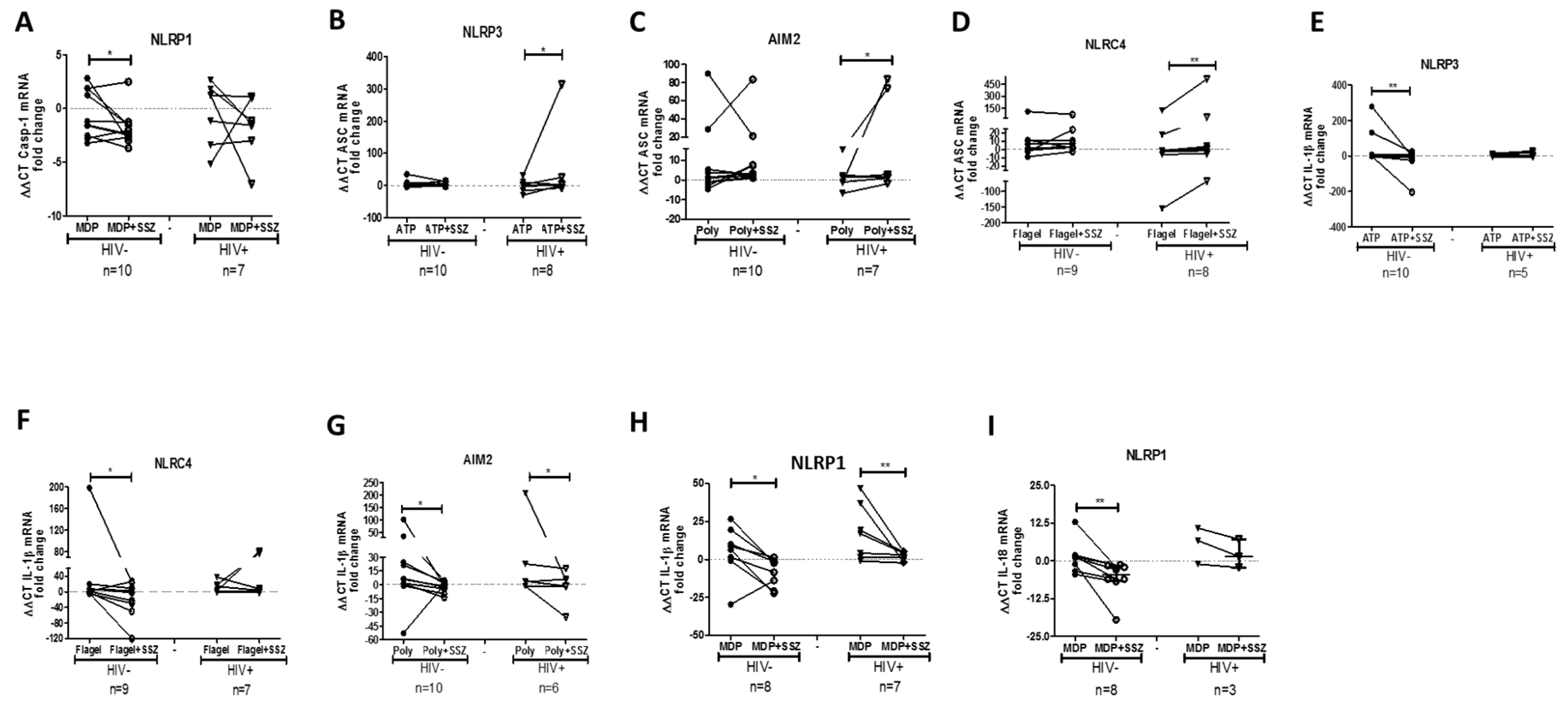
2.3. SSZ Treatment Modifies the Expression of Inflammasome-Related Genes
2.4. SSZ Modulates the TLRs Activation in PBMCs
2.5. PBMCs from People Living with HIV with High CD4+ T Cell Count and Low Viral Load Exhibit an Enhanced Response to SSZ
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Plasma Viral Load and CD4+ T-Cells Count
4.3. Reagents
4.4. Determination of the Cytotoxicity of SSZ
4.5. In Vitro Activation of Inflammasomes and SSZ Treatment
4.6. In Vitro Activation of TLR and SSZ Treatment
4.7. ELISA
4.8. mRNA Expression of Inflammasome-Related Genes
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lawrence, T.; Gilroy, D.W. Chronic inflammation: A failure of resolution? Int. J. Exp. Pathol. 2007, 88, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Paiardini, M.; Muller-Trutwin, M. HIV-associated chronic immune activation. Immunol. Rev. 2013, 254, 78–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Kanneganti, T.D.; Lamkanfi, M.; Nunez, G. Intracellular NOD-like receptors in host defense and disease. Immunity 2007, 27, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Rodriguez, E.V.; Napolitani, G.; Lanzavecchia, A.; Sallusto, F. Interleukins 1beta and 6 but not transforming growth factor-beta are essential for the differentiation of interleukin 17-producing human T helper cells. Nat. Immunol. 2007, 8, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Song, R.; Wakeland, E.K.; Pasare, C. T cell-intrinsic IL-1R signaling licenses effector cytokine production by memory CD4 T cells. Nat. Commun. 2018, 9, 3185. [Google Scholar] [CrossRef]
- Revu, S.; Wu, J.; Henkel, M.; Rittenhouse, N.; Menk, A.; Delgoffe, G.M.; Poholek, A.C.; McGeachy, M.J. IL-23 and IL-1β Drive Human Th17 Cell Differentiation and Metabolic Reprogramming in Absence of CD28 Costimulation. Cell Rep. 2018, 22, 2642–2653. [Google Scholar] [CrossRef]
- Coccia, M.; Harrison, O.J.; Schiering, C.; Asquith, M.J.; Becher, B.; Powrie, F.; Maloy, K.J. IL-1β mediates chronic intestinal inflammation by promoting the accumulation of IL-17A secreting innate lymphoid cells and CD4(+) Th17 cells. J. Exp. Med. 2012, 209, 1595–1609. [Google Scholar] [CrossRef]
- Hernández López, J.C.; Urcuqui Inchima, S. Activación y regulación del inflamasoma NLRP3 en las enfermedades infecciosas. Iatreia 2012, 25, 380–390. [Google Scholar]
- Yerramothu, P.; Vijay, A.K.; Willcox, M.D.P. Inflammasomes, the eye and anti-inflammasome therapy. Eye (Lond.) 2018, 32, 491–505. [Google Scholar] [CrossRef]
- Place, D.E.; Kanneganti, T.D. Recent advances in inflammasome biology. Curr. Opin. Immunol. 2017, 50, 32–38. [Google Scholar] [CrossRef]
- Hernandez, J.; Sirois, C.; Latz, E. Activation and Regulation of the NLRP3 Inflammasome. Progress in Inflammation Research; Springer: Basel, Switzerland, 2011; pp. 197–208. [Google Scholar]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.C.; Latz, E.; Urcuqui-Inchima, S. HIV-1 induces the first signal to activate the NLRP3 inflammasome in monocyte-derived macrophages. Intervirology 2014, 57, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Feria, M.G.; Taborda, N.A.; Hernandez, J.C.; Rugeles, M.T. HIV replication is associated to inflammasomes activation, IL-1beta, IL-18 and caspase-1 expression in GALT and peripheral blood. PLoS ONE 2018, 13, e0192845. [Google Scholar] [CrossRef] [PubMed]
- Plosker, G.L.; Croom, K.F. Sulfasalazine: A review of its use in the management of rheumatoid arthritis. Drugs 2015, 65, 1825–1849. [Google Scholar] [CrossRef]
- Mijiyawa, M.; David, M. Beneficial effect of sulfasalazine in spondylarthropathy in 2 patients with HIV infection. Rev. Rhum. Ed. Fr. 1993, 60, 527–529. [Google Scholar]
- Youssef, P.P.; Bertouch, J.V.; Jones, P.D. Successful treatment of human immunodeficiency virus-associated Reiter’s syndrome with sulfasalazine. Arthritis Rheum. 1992, 35, 723–724. [Google Scholar] [CrossRef]
- Hunt, P.W. HIV and inflammation: Mechanisms and consequences. Curr. Hiv/Aids Rep. 2012, 9, 139–147. [Google Scholar] [CrossRef]
- Wahl, C.; Liptay, S.; Adler, G.; Schmid, R.M. Sulfasalazine: A potent and specific inhibitor of nuclear factor kappa B. J. Clin. Invest. 1998, 101, 1163–1174. [Google Scholar] [CrossRef]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef]
- Rodenburg, R.J.; Ganga, A.; van Lent, P.L.; van de Putte, L.B.; van Venrooij, W.J. The antiinflammatory drug sulfasalazine inhibits tumor necrosis factor alpha expression in macrophages by inducing apoptosis. Arthritis Rheum. 2000, 43, 1941–1950. [Google Scholar] [CrossRef]
- Volin, M.V.; Campbell, P.L.; Connors, M.A.; Woodruff, D.C.; Koch, A.E. The effect of sulfasalazine on rheumatoid arthritic synovial tissue chemokine production. Exp Mol. Pathol. 2002, 73, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Perdomo-Celis, F.; Feria, M.G.; Taborda, N.A.; Rugeles, M.T. A Low Frequency of IL-17-Producing CD8(+) T-Cells Is Associated With Persistent Immune Activation in People Living With HIV Despite HAART-Induced Viral Suppression. Front. Immunol. 2018, 9, 2502. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.J. Of Inflammasomes and Alarmins: IL-1beta and IL-1alpha in Kidney Disease. J. Am. Soc. Nephrol. 2016, 27, 2564–2575. [Google Scholar] [CrossRef]
- Jo, E.K.; Kim, J.K.; Shin, D.M.; Sasakawa, C. Molecular mechanisms regulating NLRP3 inflammasome activation. Cell Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Groslambert, M.; Py, B.F. Spotlight on the NLRP3 inflammasome pathway. J. Inflamm Res. 2018, 11, 359–374. [Google Scholar] [CrossRef]
- Chang, J.J.; Altfeld, M. Immune activation and the role of TLRs and TLR agonists in the pathogenesis of HIV-1 infection in the humanized mouse model. J. Infect. Dis. 2013, 208 (Suppl. 2), S145–S149. [Google Scholar] [CrossRef]
- Meier, A.; Altfeld, M. Toll-like receptor signaling in HIV-1 infection: A potential target for therapy? Expert Rev. Anti-Infect Ther. 2007, 5, 323–326. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Xiao, T.S. Post-translational regulation of inflammasomes. Cell Mol. Immunol. 2017, 14, 65–79. [Google Scholar] [CrossRef]
- Zhong, Z.; Umemura, A.; Sanchez-Lopez, E.; Liang, S.; Shalapour, S.; Wong, J.; He, F.; Boassa, D.; Perkins, G.; Ali, S.R.; et al. NF-kappaB Restricts Inflammasome Activation via Elimination of Damaged Mitochondria. Cell 2016, 164, 896–910. [Google Scholar] [CrossRef]
- Nakahira, K.; Haspel, J.A.; Rathinam, V.A.; Lee, S.J.; Dolinay, T.; Lam, H.C.; Englert, J.A.; Rabinovitch, M.; Cernadas, M.; Kim, H.P.; et al. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nat. Immunol. 2011, 12, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, M.; Kageyama, S.; Ichimura, Y. p62/SQSTM1/A170: Physiology and pathology. Pharm. Res. 2012, 66, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Funderburg, N.T.; Jiang, Y.; Debanne, S.M.; Labbato, D.; Juchnowski, S.; Ferrari, B.; Clagett, B.; Robinson, J.; Lederman, M.M.; McComsey, G.A. Rosuvastatin reduces vascular inflammation and T-cell and monocyte activation in HIV-infected subjects on antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2015, 68, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Funderburg, N.T.; Jiang, Y.; Debanne, S.M.; Storer, N.; Labbato, D.; Clagett, B.; Robinson, J.; Lederman, M.M.; McComsey, G.A. Rosuvastatin treatment reduces markers of monocyte activation in HIV-infected subjects on antiretroviral therapy. Clin. Infect. Dis. 2014, 58, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Piconi, S.; Parisotto, S.; Rizzardini, G.; Passerini, S.; Terzi, R.; Argenteri, B.; Meraviglia, P.; Capetti, A.; Biasin, M.; Trabattoni, D.; et al. Hydroxychloroquine drastically reduces immune activation in HIV-infected, antiretroviral therapy-treated immunologic nonresponders. Blood 2011, 118, 3263–3272. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.; Montenont, E.; Hu, L.; Nardi, M.A.; Valdes, V.; Merolla, M.; Gettenberg, G.; Cavanagh, K.; Aberg, J.A.; Bhardwaj, N.; et al. Aspirin attenuates platelet activation and immune activation in HIV-1-infected subjects on antiretroviral therapy: A pilot study. J. Acquir. Immune Defic. Syndr. 2013, 63, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, E.; Boada, J.; Perales, J.C.; Roig, T.; Bermudez, J. Aspirin inhibits NF-kappaB activation in a glycolysis-depleted lung epithelial cell line. Eur. J. Pharm. 2005, 517, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, J.; Ge, X.; Du, J.; Deb, D.K.; Li, Y.C. Vitamin D receptor inhibits nuclear factor kappaB activation by interacting with IkappaB kinase beta protein. J. Biol. Chem. 2013, 288, 19450–19458. [Google Scholar] [CrossRef] [PubMed]
- Opravil, M.; Ledergerber, B.; Furrer, H.; Hirschel, B.; Imhof, A.; Gallant, S.; Wagels, T.; Bernasconi, E.; Meienberg, F.; Rickenbach, M.; et al. Clinical efficacy of early initiation of HAART in patients with asymptomatic HIV infection and CD4 cell count > 350 × 10(6)/l. AIDS 2002, 16, 1371–1381. [Google Scholar] [CrossRef]
- Arcia, D.; Ochoa, R.; Hernandez, J.C.; Álvarez, C.M.; Díaz, F.J.; Velilla, P.A.; Acevedo-Sáenz, L. Potential immune escape mutations under inferred selection pressure in HIV-1 strains circulating in Medellín, Colombia. Infect. Genet. Evol. 2019, 69, 267–278. [Google Scholar] [CrossRef]
- Shoji, K.F.; Debure, L. Fluorometric Methods for Detection of Mitochondrial Membrane Depolarization Induced by CD95 Activation. Methods Mol. Biol. 2017, 1557, 49–62. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| People Living with HIV (n = 15) | Healthy Donors (n = 15) | p-Value | |
|---|---|---|---|
| Age in years, median (min-max) | 28 (18–51) | 29 (19–55) | 0.49117 |
| Sex, Male:Female | 11:4 | 8:7 | 0.1498 |
| Time of diagnosis in months median (min-max) | 15 (1–85) | - | - |
| CD4+ T cell count in the blood median (min-max) | 438 (316–840) | - | - |
| Plasma HIV viral load in RNA copies/mL median (min-max) | 16,660 (2500–140,400) | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feria-Garzón, M.G.; Rugeles, M.T.; Hernandez, J.C.; Lujan, J.A.; Taborda, N.A. Sulfasalazine as an Immunomodulator of the Inflammatory Process during HIV-1 Infection. Int. J. Mol. Sci. 2019, 20, 4476. https://doi.org/10.3390/ijms20184476
Feria-Garzón MG, Rugeles MT, Hernandez JC, Lujan JA, Taborda NA. Sulfasalazine as an Immunomodulator of the Inflammatory Process during HIV-1 Infection. International Journal of Molecular Sciences. 2019; 20(18):4476. https://doi.org/10.3390/ijms20184476
Chicago/Turabian StyleFeria-Garzón, Manuel G., María T. Rugeles, Juan C. Hernandez, Jorge A. Lujan, and Natalia A. Taborda. 2019. "Sulfasalazine as an Immunomodulator of the Inflammatory Process during HIV-1 Infection" International Journal of Molecular Sciences 20, no. 18: 4476. https://doi.org/10.3390/ijms20184476
APA StyleFeria-Garzón, M. G., Rugeles, M. T., Hernandez, J. C., Lujan, J. A., & Taborda, N. A. (2019). Sulfasalazine as an Immunomodulator of the Inflammatory Process during HIV-1 Infection. International Journal of Molecular Sciences, 20(18), 4476. https://doi.org/10.3390/ijms20184476