Xanthones from the Bark of Garcinia xanthochymus and the Mechanism of Induced Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells via the Mitochondrial Pathway


Abstract
:1. Introduction
2. Results
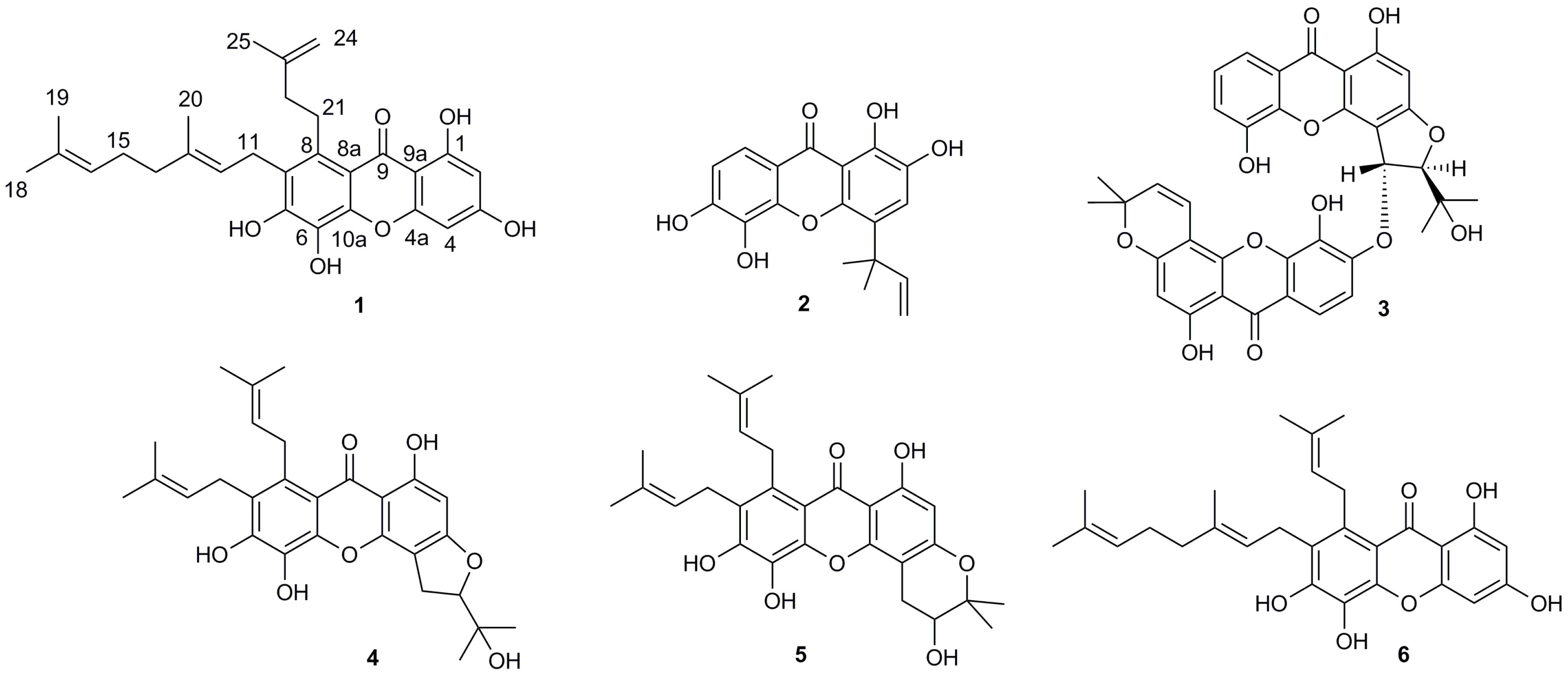
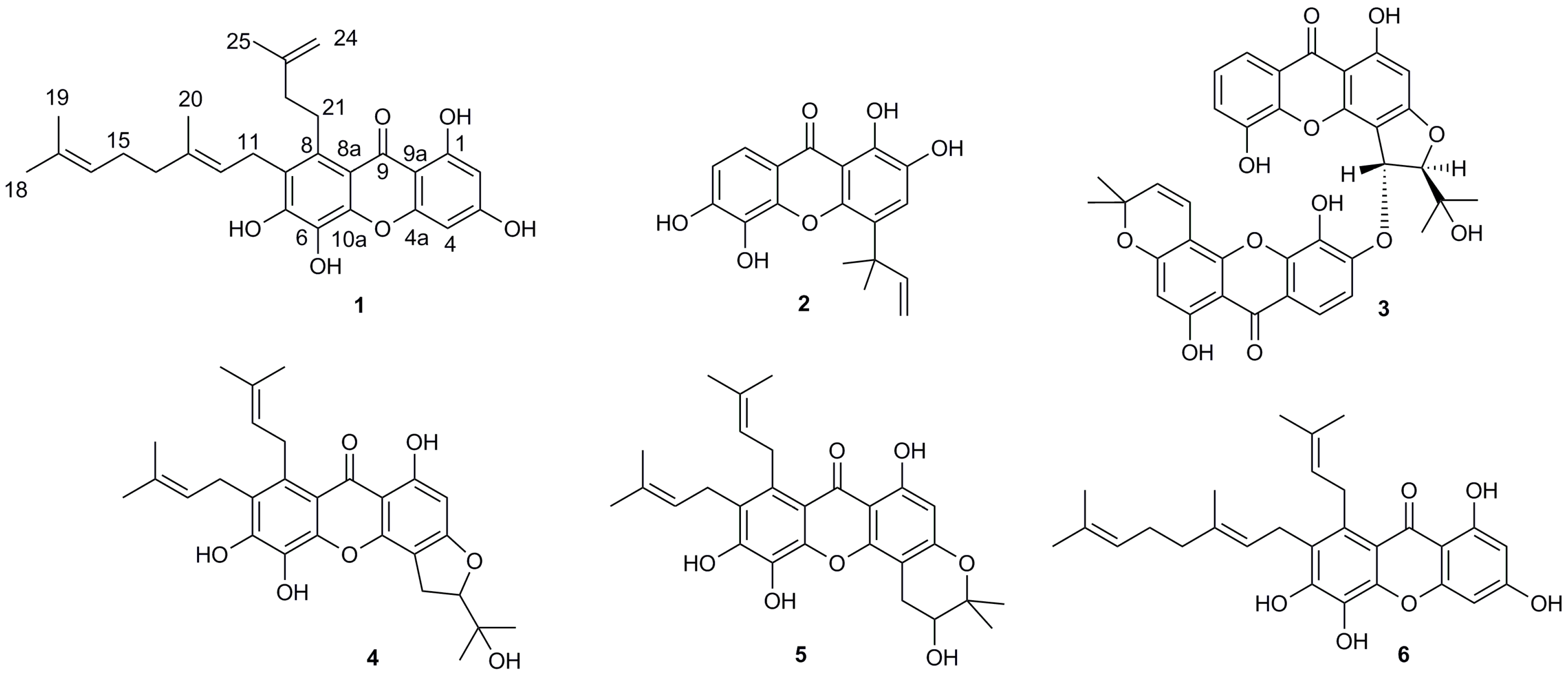
2.1. Structural Elucidation of Isolated Compounds
2.2. Anti-Proliferation Activity on Human Tumor Cell Lines
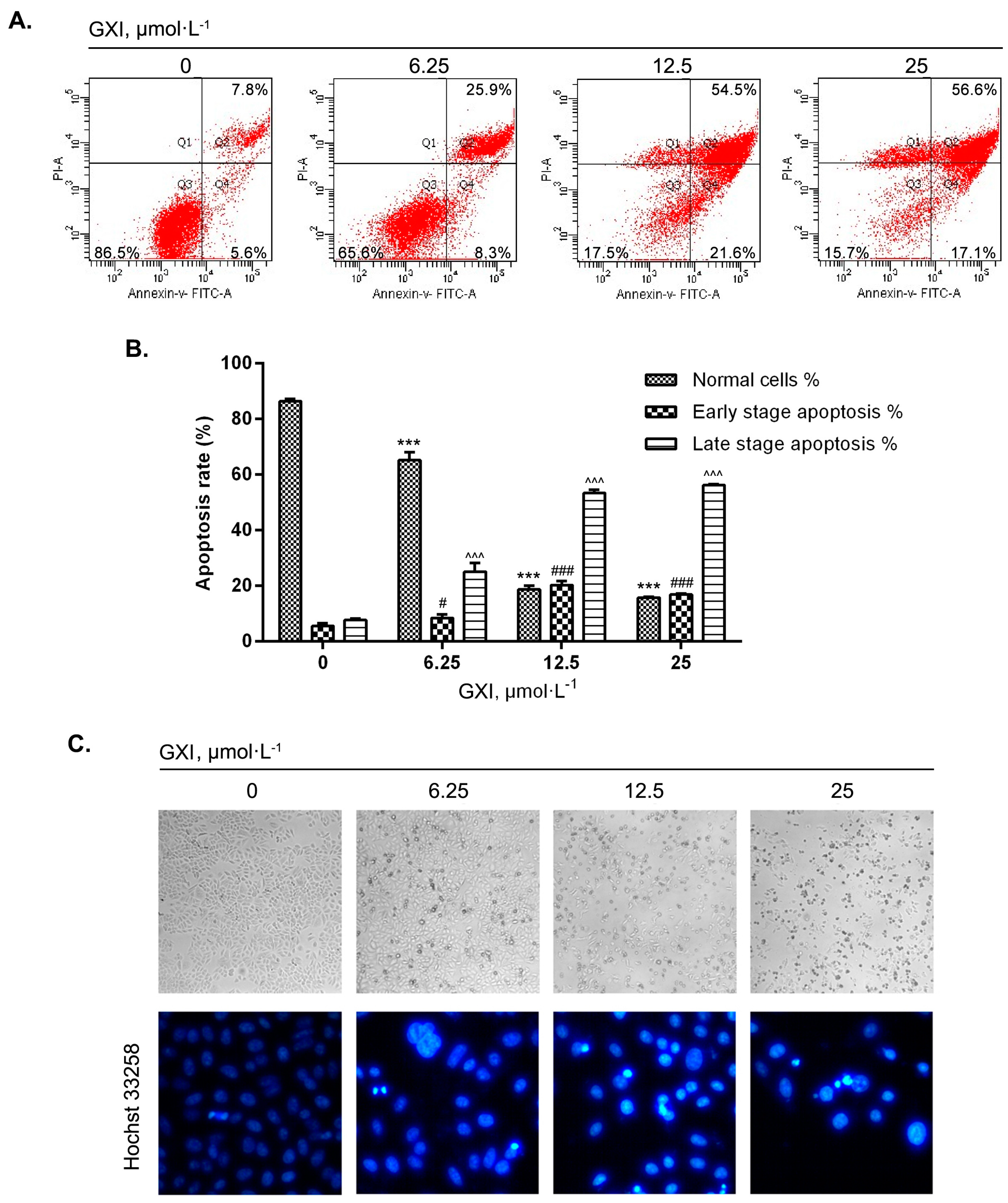
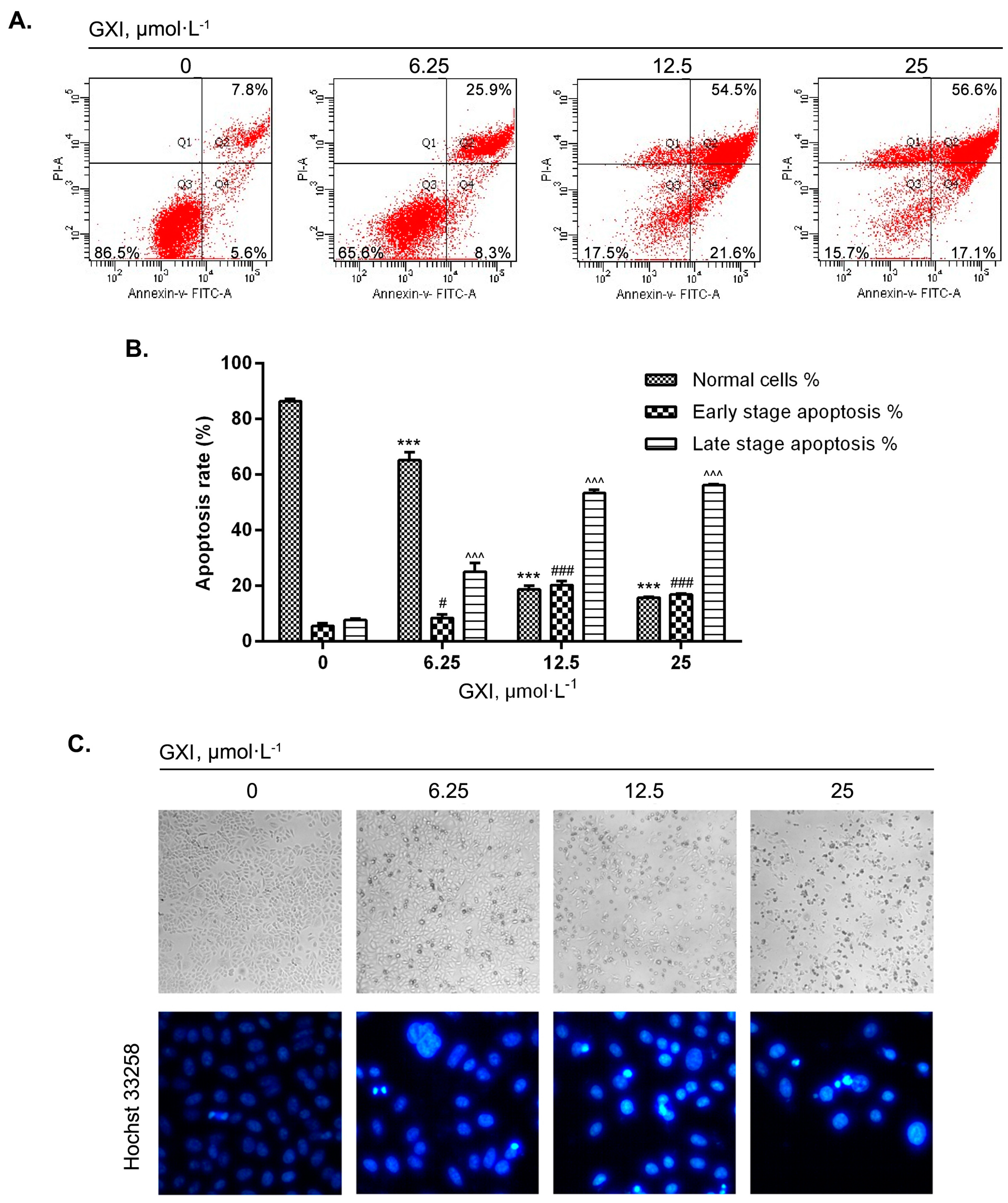
2.3. GXI Induced HepG2 Apoptosis
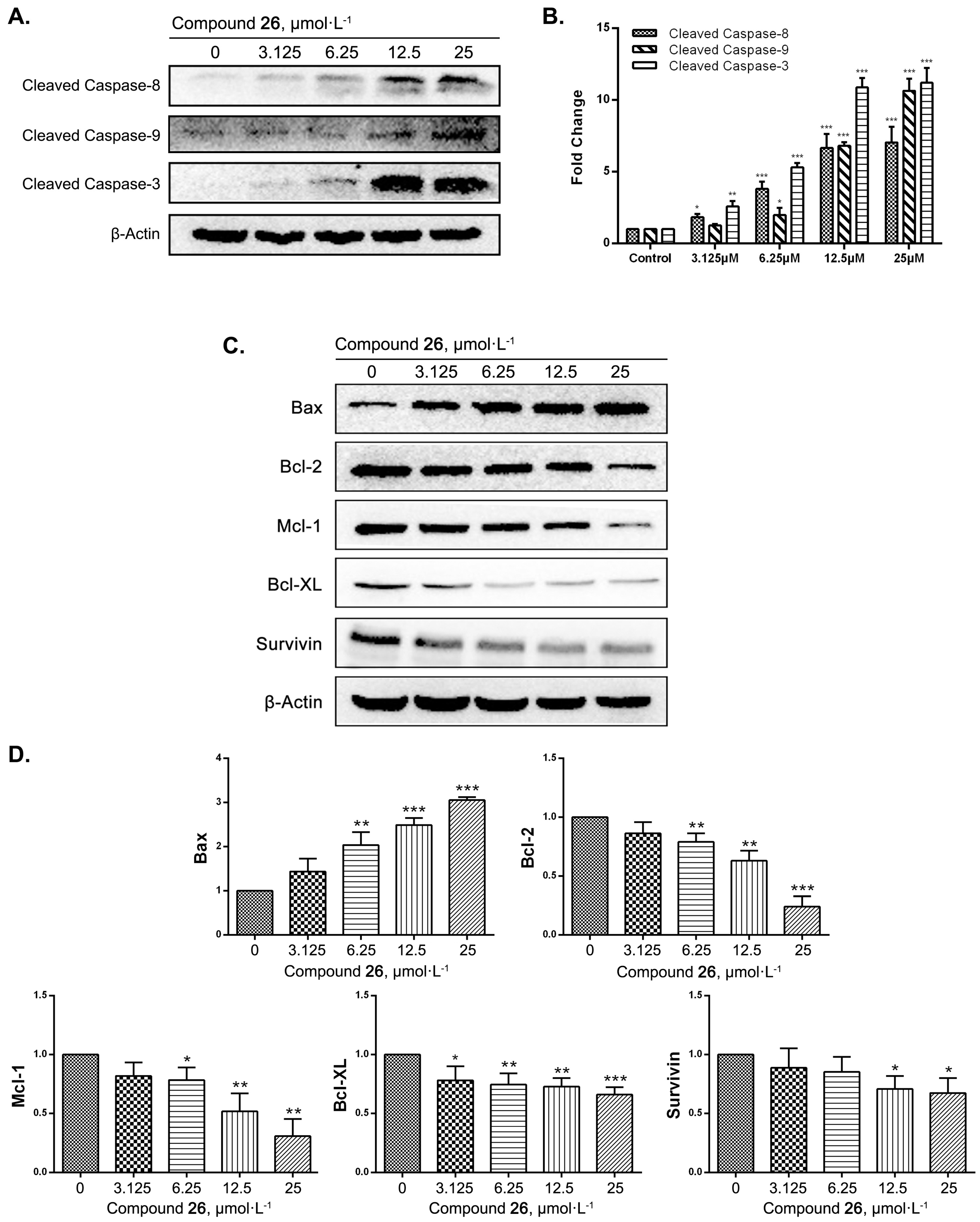
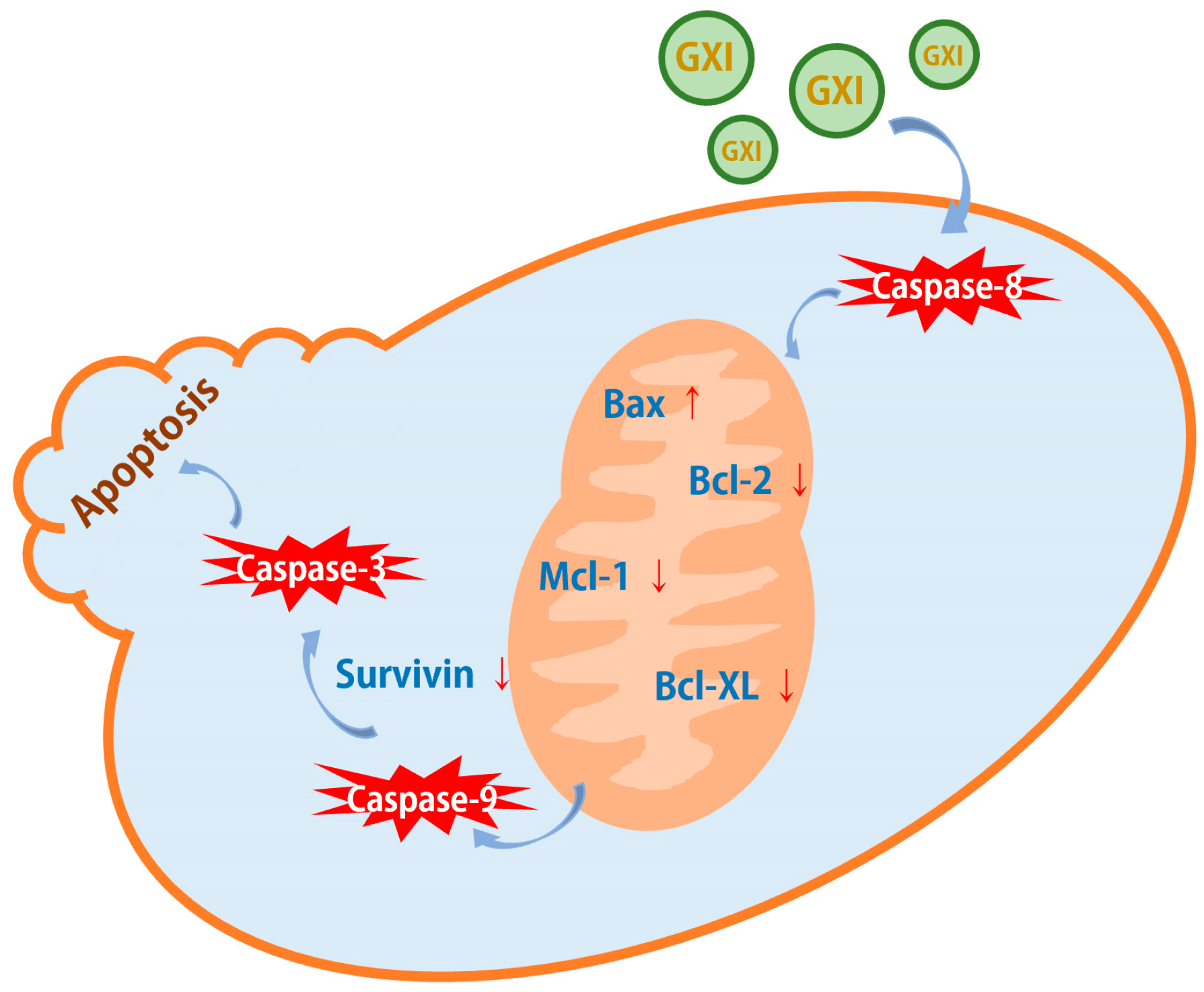
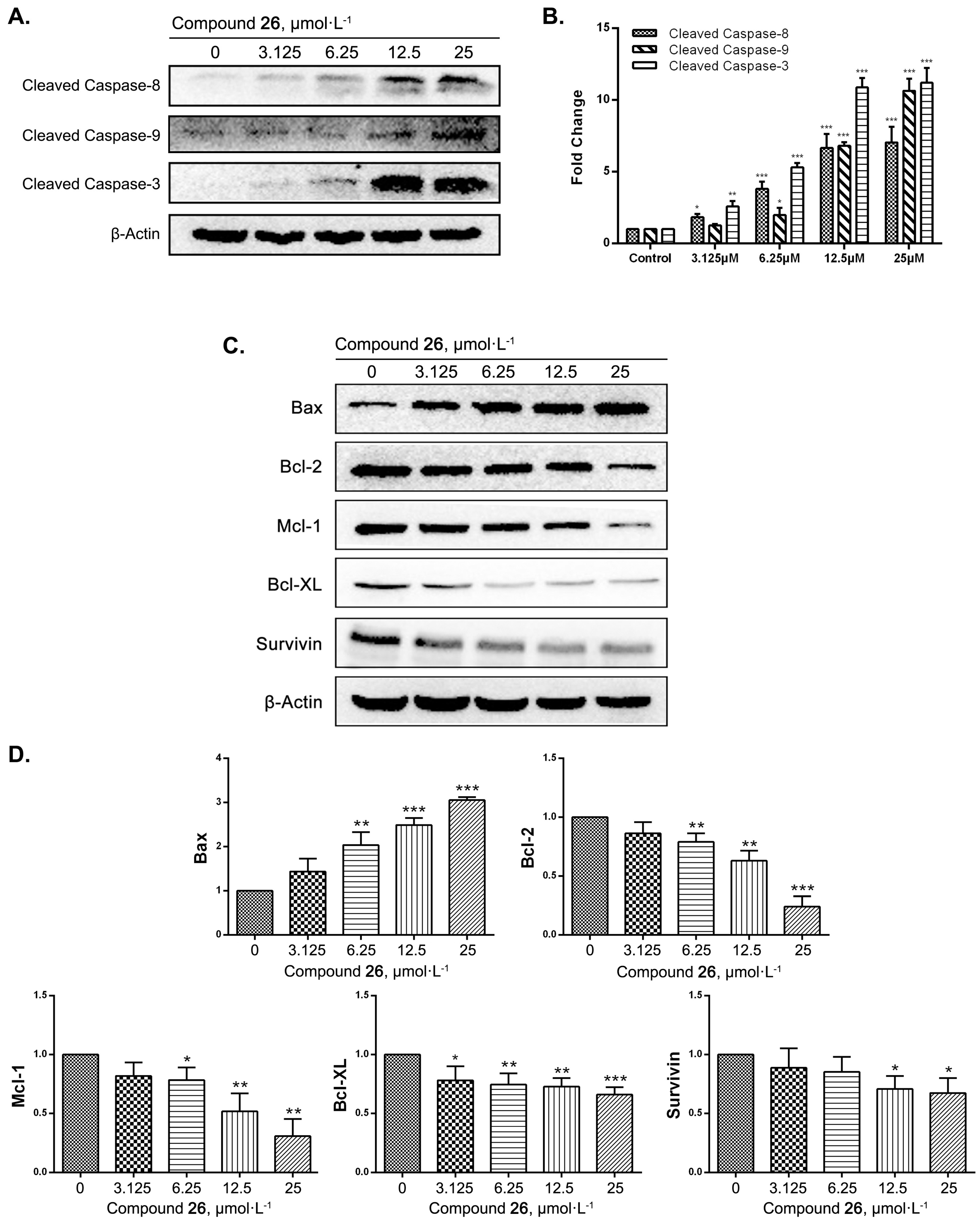
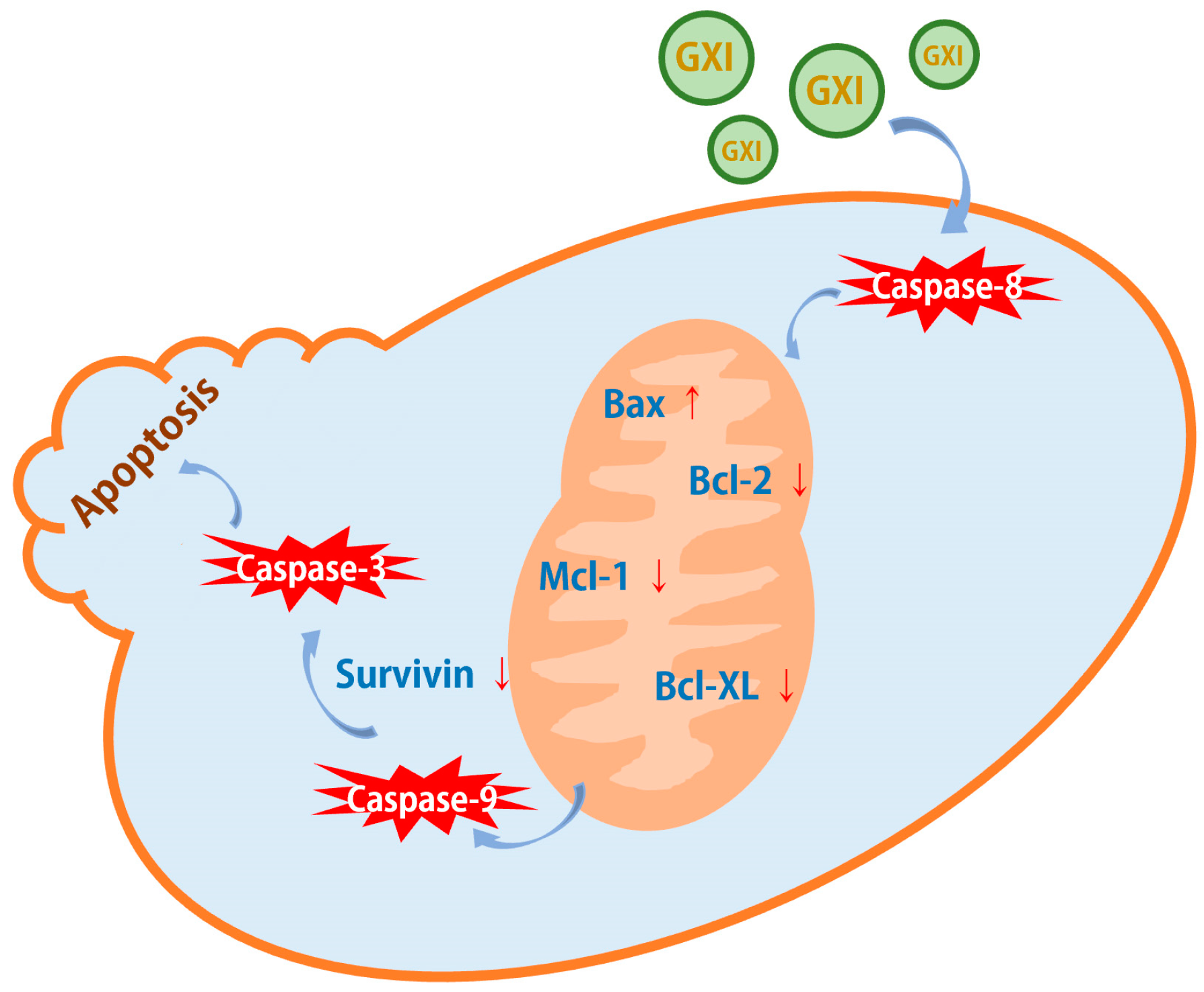
2.4. GXI-Induced HepG2 Apoptosis via the Mitochondrial Pathway
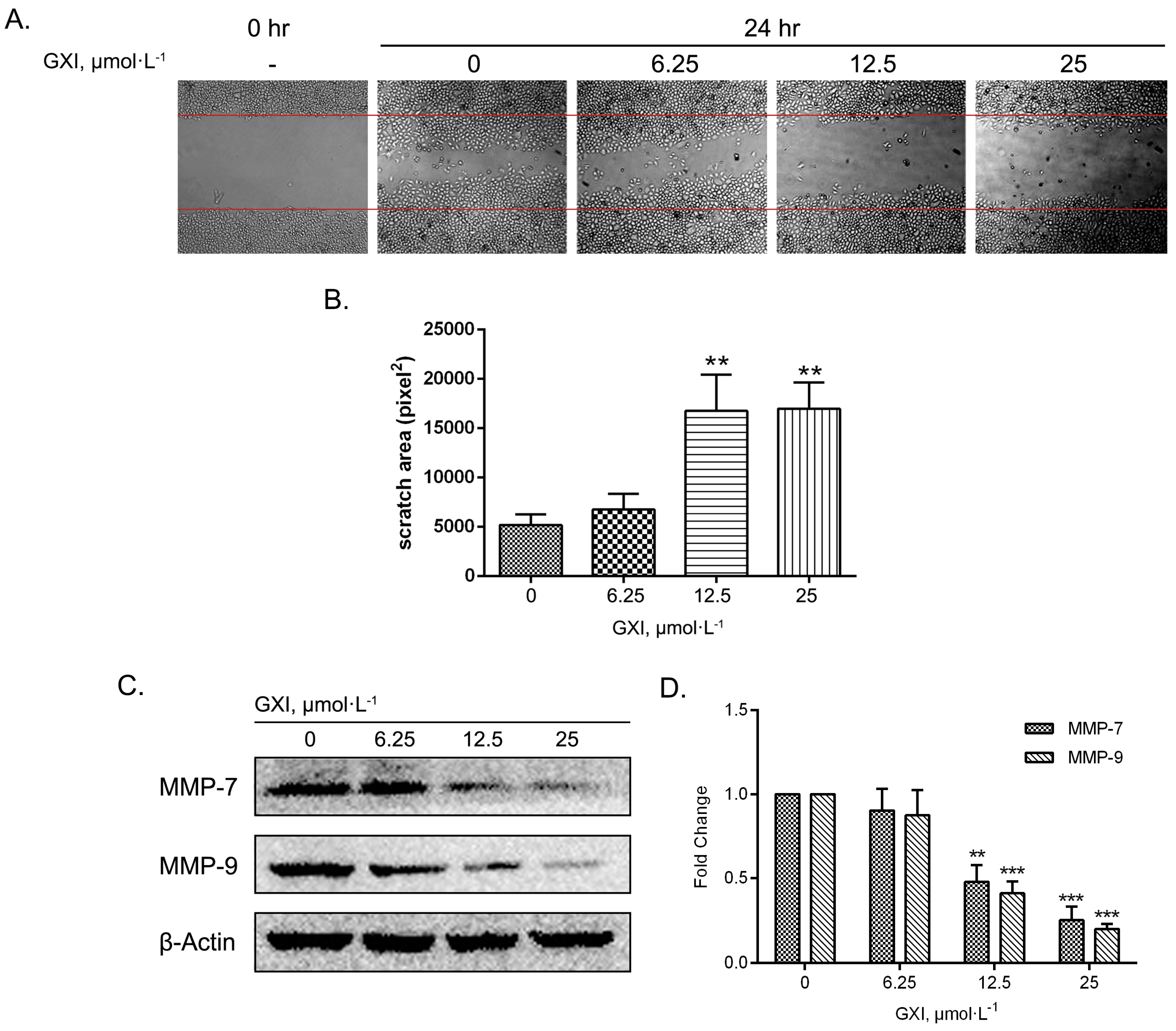
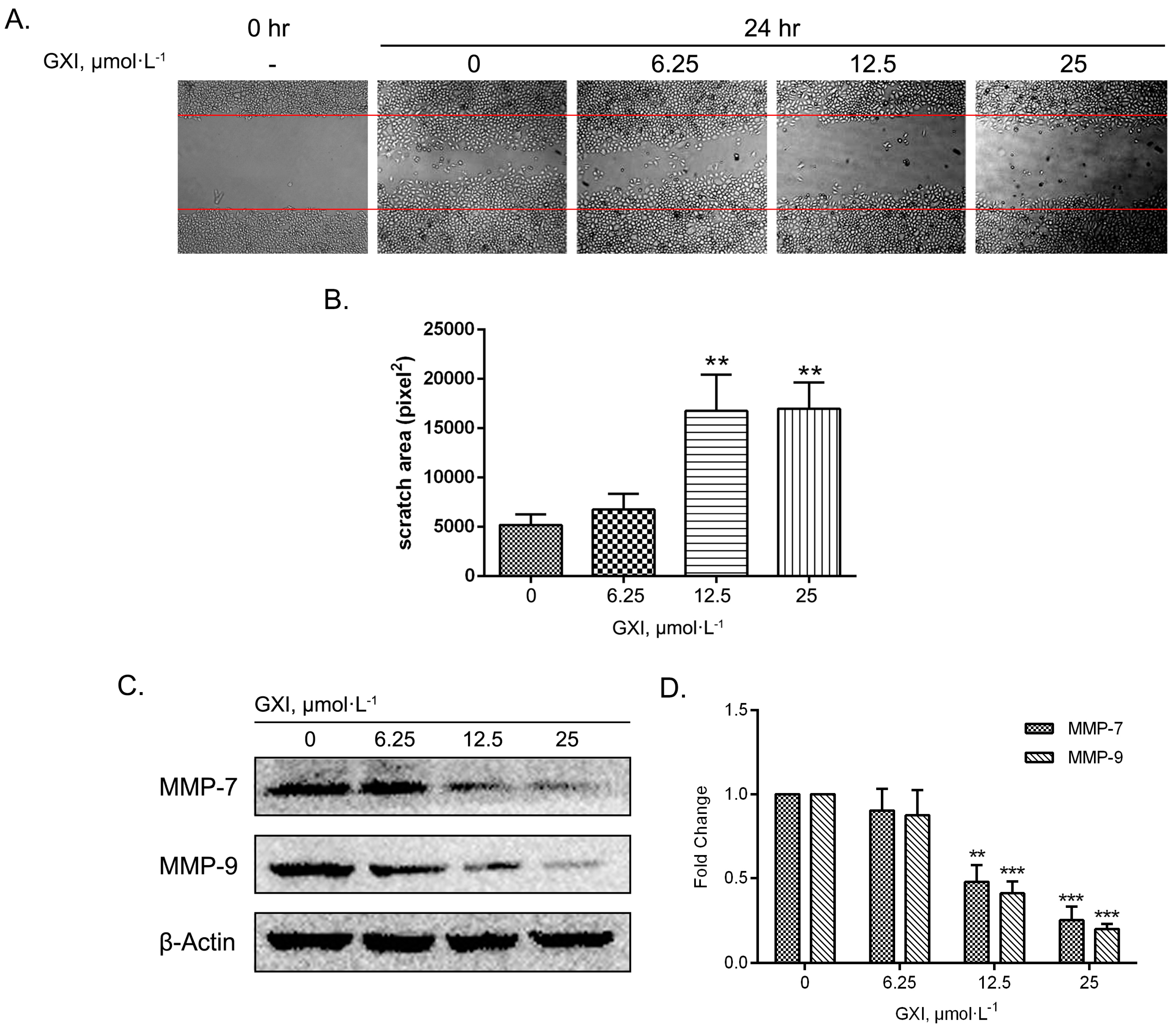
2.5. GXI Induced HepG2 Migration
3. Discussion
4. Materials and Methods
4.1. General
4.2. Plant Material
4.3. Extraction and Isolation
4.4. Cell Culture
4.5. Cytotoxicity Assay
4.6. Fluorescence Staining Assay
4.7. FACS Analysis
4.8. Western Blotting
4.9. Wound Healing Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HPLC | High-performance liquid chromatography |
| HMBC | Heteronuclear multiple bond correlation |
| SAR | Structure–activity relationship |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| FCM | Flow cytometry |
| Bcl-2 | B-cell lymphoma 2 |
| MMP | Matrix metalloproteinase |
| OD | Optical density |
| FITC | Fluorescein isothiocyanate |
| PI | Propidium iodide |
| DMSO | Dimethyl sulfoxide |
References
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 12 September 2018).
- Burke, P.J. Mitochondria, Bioenergetics and Apoptosis in Cancer. Trends Cancer 2017, 3, 857–870. [Google Scholar] [CrossRef] [PubMed]
- Che Hassan, N.K.N.; Taher, M.; Susanti, D. Phytochemical constituents and pharmacological properties of Garcinia xanthochymus- a review. Biomed Pharm. 2018, 106, 1378–1389. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; Huang, A.M.; Hour, T.C.; Yang, S.C.; Pu, Y.S.; Lin, C.N. Antioxidant xanthone derivatives induce cell cycle arrest and apoptosis and enhance cell death induced by cisplatin in NTUB1 cells associated with ROS. Eur. J. Med. Chem. 2011, 46, 1222–1231. [Google Scholar] [CrossRef] [PubMed]
- Cossu, F.; Milani, M.; Mastrangelo, E.; Lecis, D. Targeting the BIR Domains of Inhibitor of Apoptosis (IAP) Proteins in Cancer Treatment. Comput. Struct. Biotechnol. J. 2019, 17, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Charalambous, G.K.; Gomatos, I.P.; Konstadoulakis, M.M.; Messaris, E.G.; Manouras, A.J.; Apostolou, A.E.; Leandros, E.A.; Karayannis, M.K.; Androulakis, G.A. Protein expression of bax, bcl-2, and p53 in patients with non-Hodgkin’s gastric lymphoma: Prognostic significance. World J. Surg. 2000, 24, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Marschitz, I.; Tinhofer, I.; Hittmair, A.; Egle, A.; Kos, M.; Greil, R. Analysis of Bcl-2 protein expression in chronic lymphocytic leukemia. A comparison of three semiquantitation techniques. Am. J. Clin. Pathol. 2000, 113, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Hussin, Y.; Aziz, M.N.M.; Che Rahim, N.F.; Yeap, S.K.; Mohamad, N.E.; Masarudin, M.J.; Nordin, N.; Abd Rahman, N.M.A.; Yong, C.Y.; Akhtar, M.N.; et al. DK1 Induces Apoptosis via Mitochondria-Dependent Signaling Pathway in Human Colon Carcinoma Cell Lines In Vitro. Int. J. Mol. Sci. 2018, 19, 1151. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Akao, Y.; Ohguchi, K.; Ito, T.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Xanthones induce cell-cycle arrest and apoptosis in human colon cancer DLD-1 cells. Bioorg. Med. Chem. 2005, 13, 6064–6069. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.K.; Anu Appaiah, K.A. Application of native yeast from Garcinia (Garcinia xanthochumus) for the preparation of fermented beverage: Changes in biochemical and antioxidant properties. Food Biosci. 2014, 5, 101–107. [Google Scholar] [CrossRef]
- Sadeghi, S.; Davoodvandi, A.; Pourhanifeh, M.H.; Sharifi, N.; ArefNezhad, R.; Sahebnasagh, R.; Moghadam, S.A.; Sahebkar, A.; Mirzaei, H. Anti-cancer effects of cinnamon: Insights into its apoptosis effects. Eur. J. Med. Chem. 2019, 178, 131–140. [Google Scholar] [CrossRef]
- Wu, J.J.; Ma, T.; Wang, Z.M.; Xu, W.J.; Yang, X.L.; Luo, J.G.; Kong, L.Y.; Wang, X.B. Polycyclic xanthones via pH-switched biotransformation of α-mangostin catalysed by horseradish peroxidase exhibited cytotoxicity against hepatoblastoma cells in vitro. J. Funct. Foods 2017, 28, 205–214. [Google Scholar] [CrossRef]
- Li, Y.F.; Zhao, P.; Chen, Y.; Fu, Y.Z.; Shi, K.; Liu, L.; Xiong, M.R.; Liu, Q.H.; Yang, G.Z.; Xiao, Y.X. Depsidone and xanthones from Garcinia xanthochymus with hypoglycemic activity and the mechanism of promoting glucose uptake in L6 myotubes. Bioorgan. Med. Chem. 2017, 25, 6605–6613. [Google Scholar] [CrossRef] [PubMed]
- Hiroyuki, M.; Emi, T.; Mitsuaki, K.; Yoshiyasu, F. Three xanthones from Garcinia subelliptica. Phytochemistry 1996, 41, 629–633. [Google Scholar]
- Zhong, F.F.; Chen, Y.; Wang, P.; Feng, H.J.; Yang, G.Z. Xanthones from the Bark of Garcinia xanthochymus and Their 1,1-Diphenyl-2-picrylhydrazyl Radical-Scavenging Activity. Chin. J. Chem. 2009, 27, 74–80. [Google Scholar] [CrossRef]
- Wu, Q.L.; Wang, S.P.; Du, L.J.; Yang, J.S.; Xiao, P.G. Xanthones from hypericum japonicum and H. henryi. Phytochemistry 1998, 49, 1395–1402. [Google Scholar] [CrossRef]
- Nuangnaowarat, W.; Phupong, W.; Isaka, M. New xanthones from the barks of Cratoxylum sumatranum ssp. Neriifolium. Heterocycles 2010, 81, 2335–2341. [Google Scholar]
- Ishiguro, K.; Nagata, S.; Oku, H.; Yamaki, M. Bisxanthones from Hypericum japonicum: Inhibitors of PAF-induced hypotension. Planta Med. 2002, 68, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Lia, Z.; Liu, G.; Niu, S.; Zhao, N.; Liu, X.; Hua, H. Xanthones with antiproliferative effects on prostate cancer cells from the stem bark of Garcinia xanthochymus. Nat. Prod. Commun. 2012, 7, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, G.Z.; Zhong, F.F.; He, H.W. Two New Prenylated Xanthones from the Bark of Garcinia xanthochymus. B. Kor. Chem. Soc. 2010, 31, 3418–3420. [Google Scholar] [CrossRef]
- Han, Q.B.; Qiao, C.F.; Song, J.Z.; Yang, N.Y.; Cao, X.W.; Peng, Y.; Yang, D.J.; Chen, S.L.; Xu, H.X. Cytotoxic Prenylated Phenolic Compounds from the Twig Bark of Garcinia xanthochymus. Chem. Biodiver. 2007, 4, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Edlich, F. BCL-2 proteins and apoptosis: Recent insights and unknowns. Biochem. Biophys. Res. Commun. 2018, 500, 26–34. [Google Scholar] [CrossRef]
- Fathi, N.; Rashidi, G.; Khodadadi, A.; Shahi, S. and Sharifi, S. STAT3 and apoptosis challenges in cancer. Int. J. Biol. Macromol. 2018, 117, 993–1001. [Google Scholar] [CrossRef]
- Han, C.; Li, Z.; Hou, J.; Wang, Z.; Xu, D.; Xue, G.; Kong, L. Bioactivity evaluation of natural product alpha-mangostin as a novel xanthone-based lysine-specific demethylase 1 inhibitor to against tumor metastasis. Bioorg. Chem. 2018, 76, 415–419. [Google Scholar] [CrossRef]
- Jaeschke, H.; Duan, L.; Akakpo, J.Y.; Farhood, A.; Ramachandran, A. The role of apoptosis in acetaminophen hepatotoxicity. Food Chem. Toxicol. 2018, 118, 709–718. [Google Scholar] [CrossRef]
- Mahmoudian-Sani, M.R.; Alghasi, A.; Saeedi-Boroujeni, A.; Jalali, A.; Jamshidi, M.; Khodadadi, A. Survivin as a diagnostic and therapeutic marker for thyroid cancer. Pathol. Res. Pract. 2019, 215, 619–625. [Google Scholar] [CrossRef]
- Liu, J.; Zhan, J.; Wang, H.; Liu, Z.; Zhang, C.; Jiang, Z.; Chen, H. Synthesis of xanthone derivatives and studies on the inhibition against cancer cells growth and synergistic combinations of them. Eur. J. Med. Chem. 2017, 133, 50–61. [Google Scholar] [CrossRef]
- Murakami, M.; Sakai, H.; Kodama, A.; Yanai, T.; Mori, T.; Maruo, K.; Masegi, T. Activation of matrix metalloproteinase (MMP)-2 by membrane type 1-MMP and abnormal immunolocalization of the basement membrane components laminin and type IV collagen in canine spontaneous hemangiosarcomas. Histol. Histopathol. 2009, 24, 437–446. [Google Scholar]
- Klein, G.; Vellenga, E.; Fraaije, M.W.; Kamps, W.A.; Bont, E.S. The possible role of matrix metalloproteinase (MMP)-2 and MMP-9 in cancer, e.g., acute leukemia. Crit. Rev. Oncol. Hematol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Roeb, E.; Arndt, M.; Jansen, B.; Schumpelick, V.; Matern, S. Simultaneous determination of matrix metalloproteinase (MMP)-7, MMP-1, -3, and -13 gene expression by multiplex PCR in colorectal carcinomas. Int. J. Colorectal Dis. 2004, 19, 518–524. [Google Scholar] [CrossRef]
- Herszényi, L.; Hritz, I.; Lakatos, G.; Varga, M.Z.; Tulassay, Z. The behavior of matrix metalloproteinases and their inhibitors in colorectal cancer. Int. J. Mol. Sci. 2012, 13, 13240–13263. [Google Scholar] [CrossRef]
- Hung, S.H.; Shen, K.H.; Wu, C.H.; Liu, C.L.; Shih, Y.W. α-mangostin suppresses PC-3 human prostate carcinoma cell metastasis by inhibiting matrix metalloproteinase-2/9 and urokinase-plasminogen expression through the JNK signaling pathway. J. Agric. Food Chem. 2009, 57, 1291–1298. [Google Scholar] [CrossRef]
- Lee, Y.B.; Ko, K.C.; Shi, M.D.; Liao, Y.C.; Chiang, T.A.; Wu, P.F.; Shih, Y.X.; Shih, Y.W. α-Mangostin, a novel dietary xanthone, suppresses TPA-mediated MMP-2 and MMP-9 expressions through the ERK signaling pathway in MCF-7 human breast adenocarcinoma cells. J. Food Sci. 2010, 75, H12–H23. [Google Scholar] [CrossRef]
- Shih, Y.W.; Chien, S.T.; Chen, P.S.; Lee, J.H.; Wu, S.H.; Yin, L.T. α-Mangostin Suppresses Phorbol 12-myristate 13-acetate-Induced MMP-2/MMP-9 Expressions via αvβ3 Integrin/FAK/ERK and NF-κB Signaling Pathway in Human Lung Adenocarcinoma A549 Cells. Cell Biochem. Biophys. 2010, 58, 31–44. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH | δC | HMBC (H→C) |
|---|---|---|---|
| C-1 | 165.1 | ||
| C-2 | 6.19 (1H, s) | 98.8 | 4, 9a, 1, 3 |
| C-3 | 165.3 | ||
| C-4 | 6.36 (1H, s) | 93.8 | 2, 9a, 4a, 3 |
| C-4a | 157.6 | ||
| C-10a | 146.5 | ||
| C-5 | 130.8 | ||
| C-6 | 150.2 | ||
| C-7 | 125.8 | ||
| C-8 | 135.8 | ||
| C-8a | 111.9 | ||
| C-9 | 183.4 | ||
| C-9a | 103.9 | ||
| C-11 | 3.54 (2H, d, J = 6.0 Hz) | 25.2 | 12, 13, 8, 6, 7 |
| C-12 | 5.14 (1H, t, J = 6.0 Hz) | 124.2 | 20, 11, 14 |
| C-13 | 135.4 | ||
| C-14 | 2.00 (2H, m) | 40.5 | 20, 15, 12, 13 |
| C-15 | 2.09 (2H, m) | 27.4 | 14, 16, 17 |
| C-16 | 5.07 (1H, t, J = 7.0 Hz) | 125.1 | 18, 19, 14 |
| C-17 | 131.8 | ||
| C-18 | 1.60 (3H, s) | 25.9 | 19, 16, 17 |
| C-19 | 1.56 (3H, s) | 17.8 | 18, 16, 17 |
| C-20 | 1.81 (3H, s) | 16.6 | 14, 12, 13 |
| C-21 | 3.44 (2H, m) | 29.2 | |
| C-22 | 2.27 (2H, t, J = 8.0 Hz) | 39.9 | 25, 21, 24, 8, 23 |
| C-23 | 147.3 | ||
| C-24 | 4.84 (1H, s) 4.77 (1H, s) | 110.2 | 25, 22, 23 |
| C-25 | 1.75 (3H, s) | 22.8 | 22, 24, 23 |
| 1-OH | 13.68 (1H, s) | 1, 2, 9a |
| Compounds 2 | HepG2 | A549 | SGC7901 | MCF-7 |
|---|---|---|---|---|
| Doxorubicin 3 | 6.52 ± 0.13 | 14.03 ± 0.21 | 7.54 ± 1.11 | 4.40 ± 1.17 |
| 1 | 24.61 ± 1.89 | 50.67 ± 4.41 | 28.31 ± 3.10 | 17.81 ± 6.91 |
| 3 | 35.06 ± 2.10 | 38.14 ± 0.06 | 25.58 ± 6.99 | 38.50 ± 9.28 |
| 4 | 19.71 ± 6.03 | 22.27 ± 2.14 | 10.15 ± 1.30 | 10.67 ± 7.85 |
| 5 | 15.88 ± 6.45 | 24.99 ± 7.67 | 5.74 ± 3.02 | 7.09 ± 1.26 |
| 6 | 20.47 ± 2.00 | 5.29 ± 1.34 | 16.58 ± 2.64 | 5.25 ± 2.96 |
| 7 | 23.12 ± 1.85 | 20.46 ± 1.45 | 16.04 ± 0.29 | 47.50 ± 2.11 |
| 8 | 4.14 ± 0.39 | 10.54 ± 0.25 | 0.07 ± 0.01 | 2.37 ± 1.38 |
| 9 | 23.62 ± 2.60 | 5.61 ± 0.60 | 13.11 ± 1.88 | 13.90 ± 0.58 |
| 10 | 20.41 ± 0.92 | 9.28 ± 2.01 | 6.96 ± 0.73 | 9.31 ± 1.19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, S.; Shi, K.; Liu, L.; Chen, Y.; Yang, G. Xanthones from the Bark of Garcinia xanthochymus and the Mechanism of Induced Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells via the Mitochondrial Pathway. Int. J. Mol. Sci. 2019, 20, 4803. https://doi.org/10.3390/ijms20194803
Jin S, Shi K, Liu L, Chen Y, Yang G. Xanthones from the Bark of Garcinia xanthochymus and the Mechanism of Induced Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells via the Mitochondrial Pathway. International Journal of Molecular Sciences. 2019; 20(19):4803. https://doi.org/10.3390/ijms20194803
Chicago/Turabian StyleJin, Shan, Kuan Shi, Liu Liu, Yu Chen, and Guangzhong Yang. 2019. "Xanthones from the Bark of Garcinia xanthochymus and the Mechanism of Induced Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells via the Mitochondrial Pathway" International Journal of Molecular Sciences 20, no. 19: 4803. https://doi.org/10.3390/ijms20194803
APA StyleJin, S., Shi, K., Liu, L., Chen, Y., & Yang, G. (2019). Xanthones from the Bark of Garcinia xanthochymus and the Mechanism of Induced Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells via the Mitochondrial Pathway. International Journal of Molecular Sciences, 20(19), 4803. https://doi.org/10.3390/ijms20194803