Membrane-Free Stem Cell Components Inhibit Interleukin-1α-Stimulated Inflammation and Cartilage Degradation In Vitro and In Vivo: A Rat Model of Osteoarthritis

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Cytotoxic Effect of Membrane-Free Stem Cells Components (MFSCC) on Chondrocytes
2.2. Effect of MFSCC on IL-1α-Induced Production of NO and PGE2
2.3. Effects of MFSCC on iNOS and COX-2 Expression in Rat Primary Chondrocytes
2.4. MFSCC Inhibited MMPs and Produced Cartilage Matrix Components in Il-1α-Stimulated Rat Chondrocytes
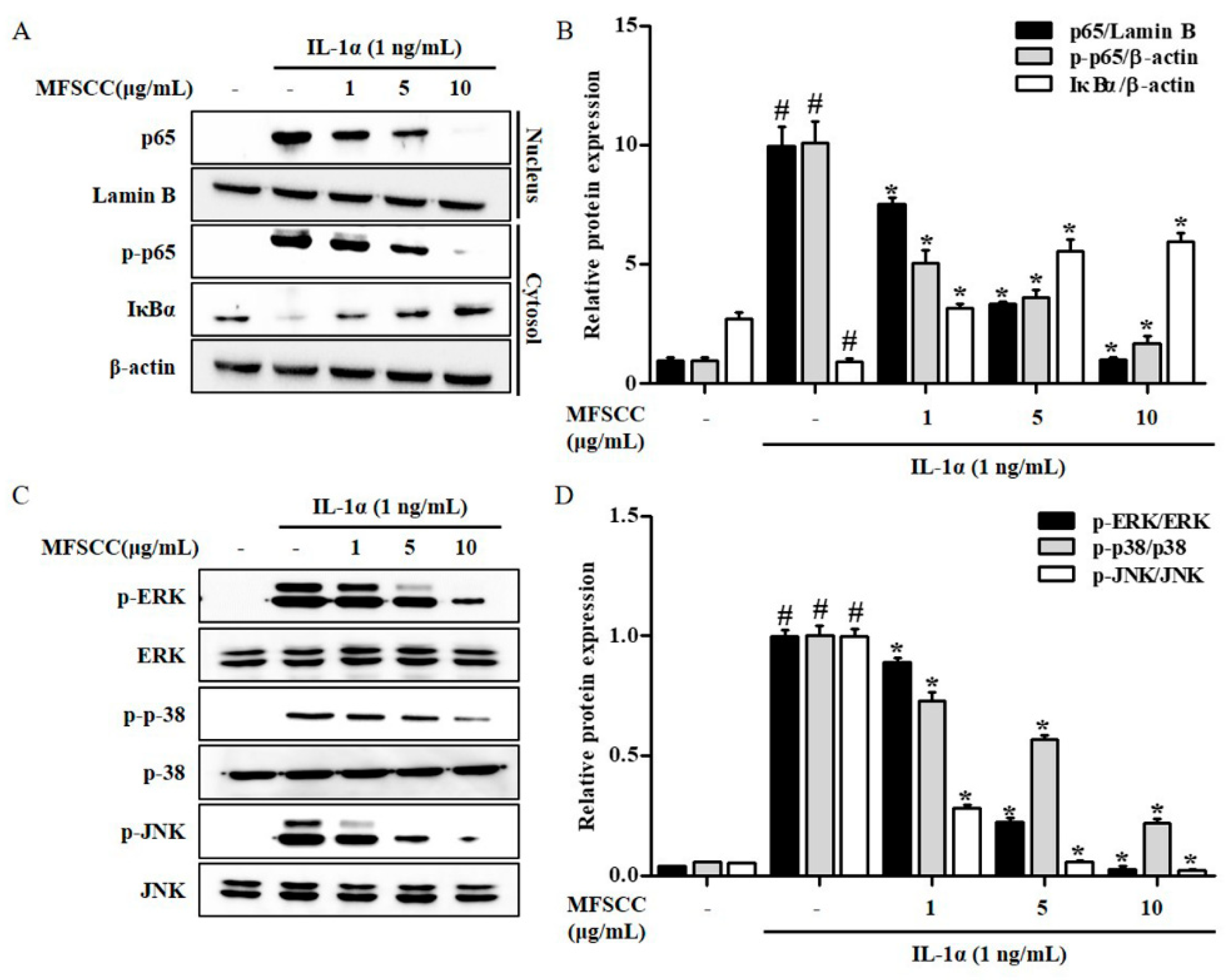
2.5. MFSCC Regulates the Nuclear Translocation of Nf-κB p65 the Expression of MAPKs in the Primary Chondrocytes Stimulated by IL-1α
2.6. Inhibition of Cytokines Expression by MFSCC
2.7. MFSCC Reduced Biomarkers of Chondrocyte in OA Rat Model
3. Discussion
4. Material and Methods
4.1. Chemicals and Reagents
4.2. Preparation of Membrane-Free Stem Cell Components (MFSCC)
4.3. Primary Chondrocyte Culture
4.4. Cell Viability Assay
4.5. NO Production Assay
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Western Blot Analysis
4.8. qRT-PCR
4.9. Animals
4.10. Induction of OA in Rats
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Neogi, T. The epidemiology and impact of pain in osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1145–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litwic, A.; Edwards, M.H.; Dennison, E.M.; Cooper, C. Epidemiology and burden of osteoarthritis. Br. Med. Bull. 2013, 105, 185–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houard, X.; Goldring, M.B.; Berenbaum, F. Homeostatic mechanisms in articular cartilage and role of inflammation in osteoarthritis. Curr. Rheumatol Rep. 2013, 15, 375. [Google Scholar] [CrossRef] [PubMed]
- Mathiessen, A.; Conaghan, P.G. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Mao, Z.; Zheng, Y.L.; Han, B.P.; Chen, L.T.; Li, J.; Li, F. Elevation of inducible nitric oxide synthase and cyclooxygenase-2 expression in the mouse brain after chronic nonylphenol exposure. Int. J. Mol. Sci. 2008, 9, 1977–1988. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [PubMed]
- Florian, A.M. IL-1 and its role in osteoarthritis. Open J. Med. 2011, 1, 3. [Google Scholar]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, C.; Huang, X.; Xiao, Y.; Song, M.; Ma, Y.; Yan, J.; You, H.; Wu, H. Schisandrin A Inhibits the IL-1beta-Induced Inflammation and Cartilage Degradation via Suppression of MAPK and NF-kappaB Signal Pathways in Rat Chondrocytes. Front. Pharm. 2019, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pizzute, T.; Pei, M. A review of crosstalk between MAPK and Wnt signals and its impact on cartilage regeneration. Cell Tissue Res. 2014, 358, 633–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Girolamo, L.; Lopa, S.; Arrigoni, E.; Sartori, M.F.; Baruffaldi Preis, F.W.; Brini, A.T. Human adipose-derived stem cells isolated from young and elderly women: Their differentiation potential and scaffold interaction during in vitro osteoblastic differentiation. Cytotherapy 2009, 11, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Dubey, N.K.; Wei, H.J.; Yu, S.H.; Williams, D.F.; Wang, J.R.; Deng, Y.H.; Tsai, F.C.; Wang, P.D.; Deng, W.P. Adipose-derived Stem Cells Attenuates Diabetic Osteoarthritis via Inhibition of Glycation-mediated Inflammatory Cascade. Aging Dis. 2019, 10, 483–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Dai, C.; Zhang, Z.; Du, H.; Li, S.; Ye, P.; Fu, Q.; Zhang, L.; Wu, X.; Dong, Y.; et al. Treatment of knee osteoarthritis with intra-articular injection of autologous adipose-derived mesenchymal progenitor cells: A prospective, randomized, double-blind, active-controlled, phase IIb clinical trial. Stem Cell Res. Ther 2019, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Matuska, A.; O’Shaughnessey, K.; King, W.; Woodell-May, J. Autologous solution protects bovine cartilage explants from IL-1alpha- and TNFalpha-induced cartilage degradation. J. Orthop Res. 2013, 31, 1929–1935. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol 2011, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, P.; Hou, Y.; Chen, S.; Xiao, Z.; Zhan, J.; Luo, D.; Gu, M.; Lin, D. Berberine inhibits the interleukin-1 beta-induced inflammatory response via MAPK downregulation in rat articular chondrocytes. Drug Dev. Res. 2019, 80, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Ye, G.; Huang, B. Kaempferol Alleviates the Interleukin-1beta-Induced Inflammation in Rat Osteoarthritis Chondrocytes via Suppression of NF-kappaB. Med. Sci Monit. 2017, 23, 3925–3931. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Markovic, B.S.; Fellabaum, C.; Arsenijevic, A.; Volarevic, V. Mesenchymal stem cell-based therapy of osteoarthritis: Current knowledge and future perspectives. Biomed. Pharm. 2019, 109, 2318–2326. [Google Scholar] [CrossRef]
- Boeth, H.; Raffalt, P.C.; MacMahon, A.; Poole, A.R.; Eckstein, F.; Wirth, W.; Buttgereit, F.; Onnerfjord, P.; Lorenzo, P.; Klint, C.; et al. Association between changes in molecular biomarkers of cartilage matrix turnover and changes in knee articular cartilage: A longitudinal pilot study. J. Exp. Orthop. 2019, 6, 19. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Sharma, A.R.; Chakraborty, C.; Saibaba, B.; Ahn, M.E.; Lee, S.S. Review of Prospects of Biological Fluid Biomarkers in Osteoarthritis. Int. J. Mol. Sci. 2017, 18, 601. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Schmitt-Talbot, E.; DiMattia, D.A.; Dullea, R.G. The differential effects of IL-1 and TNF-alpha on proinflammatory cytokine and matrix metalloproteinase expression in human chondrosarcoma cells. Inflamm. Res. 2004, 53, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Masuko-Hongo, K.; Okamoto, M.; Nishioka, K. Induction of vascular endothelial growth factor and matrix metalloproteinase-3 (stromelysin) by interleukin-1 in human articular chondrocytes and synoviocytes. Rheumatol. Int. 2005, 26, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Heinegard, D.; Saxne, T. The role of the cartilage matrix in osteoarthritis. Nat. Rev. Rheumatol 2011, 7, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Conde, J.; Scotece, M.; Abella, V.; Lois, A.; Lopez, V.; Pino, J.; Gomez, R.; Gomez-Reino, J.J.; Gualillo, O. SERPINE2 Inhibits IL-1alpha-Induced MMP-13 Expression in Human Chondrocytes: Involvement of ERK/NF-kappaB/AP-1 Pathways. PLoS ONE 2015, 10, e0135979. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, J.C.; Delmas, P.D. Biological markers in osteoarthritis. Nat. Rev. Rheumatol. 2007, 3, 346–356. [Google Scholar] [CrossRef]
- Laslett, L.L.; Quinn, S.J.; Darian-Smith, E.; Kwok, M.; Fedorova, T.; Korner, H.; Steels, E.; March, L.; Jones, G. Treatment with 4Jointz reduces knee pain over 12 weeks of treatment in patients with clinical knee osteoarthritis: A randomised controlled trial. Osteoarthr. Cartil. 2012, 20, 1209–1216. [Google Scholar] [CrossRef]
- Pan, T.; Chen, R.; Wu, D.; Cai, N.; Shi, X.; Li, B.; Pan, J. Alpha-Mangostin suppresses interleukin-1beta-induced apoptosis in rat chondrocytes by inhibiting the NF-kappaB signaling pathway and delays the progression of osteoarthritis in a rat model. Int. Immunopharmacol 2017, 52, 156–162. [Google Scholar] [CrossRef]
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef]
- Thalhamer, T.; McGrath, M.A.; Harnett, M.M. MAPKs and their relevance to arthritis and inflammation. Rheumatology (Oxford) 2008, 47, 409–414. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| iNOS | TGGATGCAACCCCATTGTC | CCCGCTGCCCCAGTTT |
| COX-2 | CAAATCCTTGCTGTTCCCACCCAT | GTGCACTGTGTTTGGAGTGGGTTT |
| MMP-3 | CTCTGGAGTAATGTCACACCTCT | TGTTGGTCCACCTTTCATCTTC |
| MMP-13 | ACGTTCAAGGAATCCAGTCTCTCT | GGATAGGGCTGGGTCACACTT |
| SOX-9 | CATCAAGACGGAGCAACTGAG | GTGGTCGGTGTAGTCATACTGC |
| Collagen II | TCAACAATGGGAAGGCGTGAG | GTTCACGTACACTGCCCTGAAG |
| Aggrecan | GGGCGTCAGAACTGTCTACC | ACTGACACACCTCGGAAGC |
| Collagen I | AGCGCTGGTTTCGACTTCAGCTTCC | CATCGGCAGGGTCGGAGCCCT |
| IL-1β | TGATGGCTTATTACAGTGGCAATG | GTAGTGGTGGTCGGAGATTCG |
| TNF-α | ATCTTCTCGAACCCCGAGTG | GGGTTTGCTACAACATGGGC |
| IL-6 | GTGTTGCCTGCTGCCTTC | AGTGCCTCTTTGCTGCTTTC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.J.; Lee, S.M.; Moon, Y.G.; Jung, Y.S.; Lee, J.H.; Venkatarame Gowda Saralamma, V.; Kim, Y.S.; Pak, J.E.; Lee, H.J.; Kim, G.S.; et al. Membrane-Free Stem Cell Components Inhibit Interleukin-1α-Stimulated Inflammation and Cartilage Degradation In Vitro and In Vivo: A Rat Model of Osteoarthritis. Int. J. Mol. Sci. 2019, 20, 4869. https://doi.org/10.3390/ijms20194869
Lee HJ, Lee SM, Moon YG, Jung YS, Lee JH, Venkatarame Gowda Saralamma V, Kim YS, Pak JE, Lee HJ, Kim GS, et al. Membrane-Free Stem Cell Components Inhibit Interleukin-1α-Stimulated Inflammation and Cartilage Degradation In Vitro and In Vivo: A Rat Model of Osteoarthritis. International Journal of Molecular Sciences. 2019; 20(19):4869. https://doi.org/10.3390/ijms20194869
Chicago/Turabian StyleLee, Ho Jeong, Seon Min Lee, Yeon Gyu Moon, Yeon Seop Jung, Ju Hong Lee, Venu Venkatarame Gowda Saralamma, Young Sil Kim, Jung Eun Pak, Hye Jin Lee, Gon Sup Kim, and et al. 2019. "Membrane-Free Stem Cell Components Inhibit Interleukin-1α-Stimulated Inflammation and Cartilage Degradation In Vitro and In Vivo: A Rat Model of Osteoarthritis" International Journal of Molecular Sciences 20, no. 19: 4869. https://doi.org/10.3390/ijms20194869