Characterization of Proteins Involved in Chloroplast Targeting Disturbed by Rice Stripe Virus by Novel Protoplast–Chloroplast Proteomics

 , , ,
, , ,
Abstract
:
1. Introduction
1.1. Plant Virus and Chloroplast
1.2. RSV Infection and Chloroplast Proteomics
1.3. RSV Infection of Nicotiana benthamiana Plants
2. Results
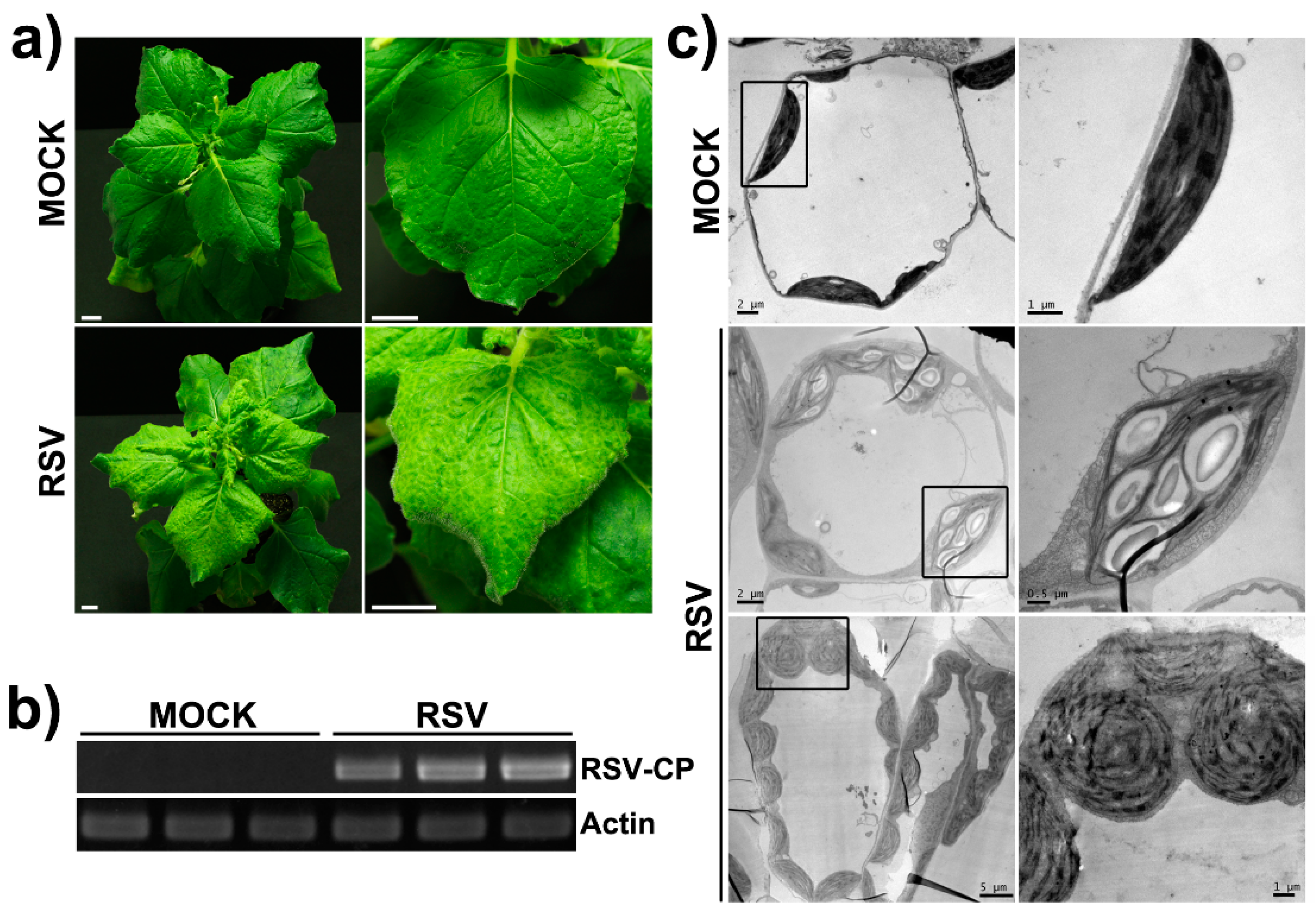
2.1. The Phenotype and the Ultrastructure Change of Chloroplasts in RSV-Infected N. benthamiana Plants
2.2. Blue Native PAGE (BN-PAGE) of Chloroplast Membrane Complexes of RSV-Infected Plants
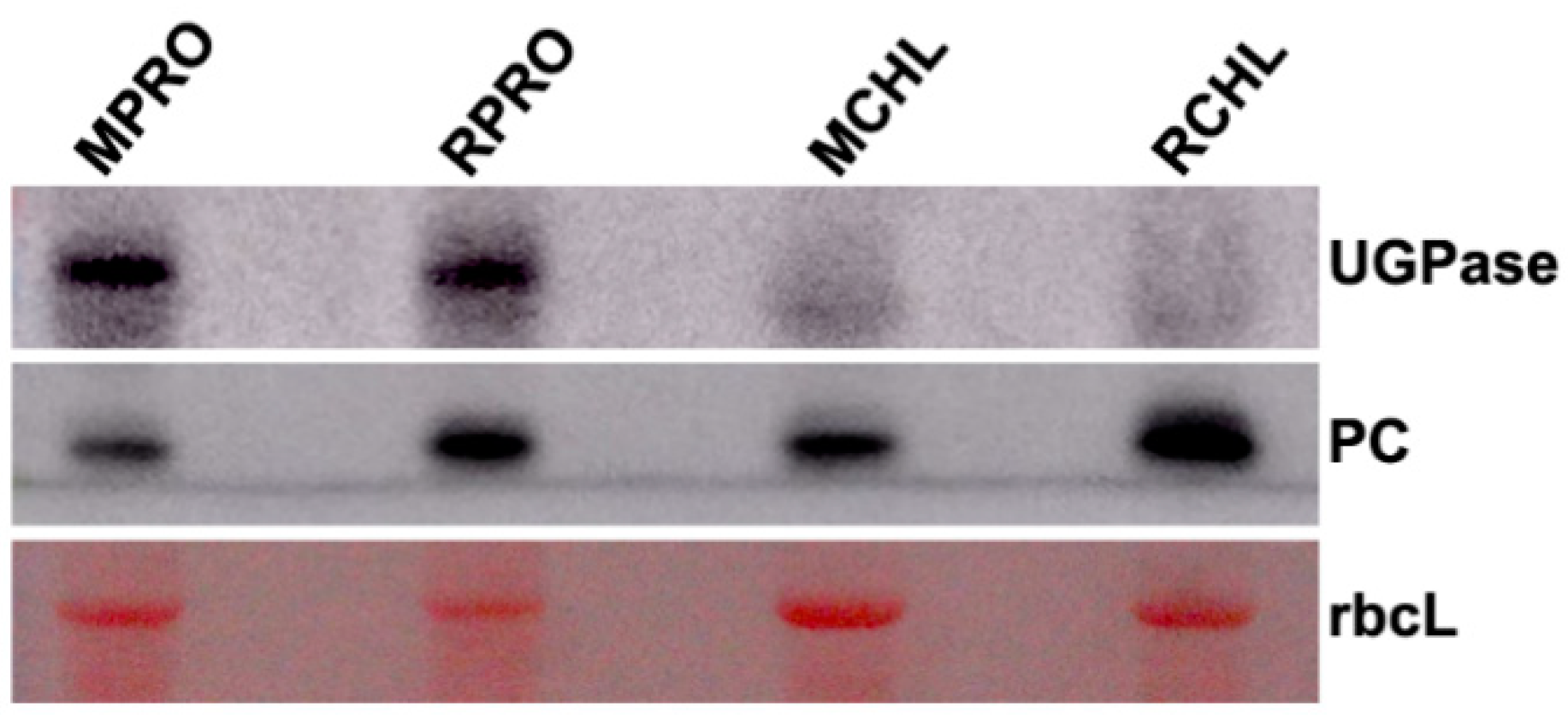
2.3. Western Blot Analysis of the Specificity of Chloroplast Protein Extracts
2.4. Analysis of Protoplast and Chloroplast Proteomics by Label-Free LC–MS/MS
2.5. GO Enrichment Analysis of the Identified Candidate Proteins
2.6. Transit Peptide Analysis of the Candidate ChRPs
3. Discussions
3.1. The Coincidence of the Chloroplast Perturbation with the RSV Symptoms
3.2. Targeting of ChRPs into Chloroplast is an Essential Layer of Virus–Chloroplast Interaction
3.3. Application of Chloroplast–Protoplast Proteomics
3.4. Candidate for Plant Virus Resistance Improvement
4. Material and methods
4.1. Plant Materials and Virus Inoculation
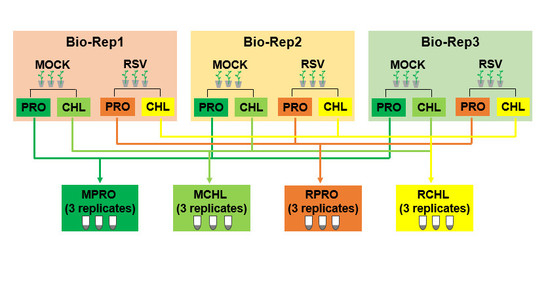
4.2. Isolation of Protoplast and Chloroplast
4.3. Blue Native PAGE Electrophoresis
4.4. Protein Extraction
4.5. Protein Digestion
4.6. Mass Spectrometric Acquisition
4.7. Mass Spectrometric Raw Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AltMV | Alternanthera mosaic virus |
| AMV | alfalfa mosaic virus |
| BaMV | bamboo mosaic virus |
| BN-PAGE | blue native PAGE |
| BsMV | barley stripe mosaic virus |
| CB | Cajal bodies |
| CF1β | chloroplast β-ATPase |
| ChRG | photosynthesis-related genes |
| ChRP | chloroplast-related protein |
| CIRV | carnation Italian ringspot virus |
| CMV | cucumber mosaic virus |
| CP | coat protein |
| CpGK | chloroplast phosphoglycerate kinase |
| cPGK | chloroplast phosphoglycerate kinase |
| cpSRP54 | chloroplast signal recognition particle 54 kDa protein |
| dpi | days post-inoculation |
| ER | endoplasmic reticulum |
| FA | formic acid |
| FC | fold change |
| Fd I | ferredoxin I |
| FDR | false discovery rate |
| GarVX | garlic virus X |
| GO | gene ontology |
| GRV | groundnut rosette virus |
| HC-Pro | helper component-proteinase |
| HR | hypersensitive reaction |
| HSP90C | HSP90 in chloroplast |
| iTRAQ | isobaric tags for relative and absolute quantification |
| MES | 2-morpholinoethanesulfonic acid |
| MNSV | melon necrotic spot carmovirus |
| MVBs | multivesicular bodies |
| OEC | oxygen evolving complex |
| PC | plastocyanin |
| PD | plasmodesma |
| PDLP | plasmodesmata-located protein |
| PLMVd | peach latent mosaic viroid |
| PMSF | phenyl methyl sulfonyl fluoride |
| PSI | photosystem I |
| PSII | photosystem II |
| PVY | potato virus Y |
| RbCL | RuBisCO large subunit |
| RbCS | RuBisCO small subunit |
| RBSDV | rice black-streak dwarf virus |
| RNAi | RNA interference |
| RSV | rice stripe virus |
| RT-PCR | reverse transcription PCR |
| RuBisCO | ribulose-1,5-bisphosphate carboxylase/oxygenase |
| TBSV | tomato bushy stunt virus |
| TECP | Tris(2-carboxyethyl)phosphine |
| TMV | tobacco mosaic virus |
| ToMV | tomato mosaic virus |
| TP | transit peptide |
| TuMV | turnip mosaic virus |
| UGPase | UDP-glucose pyrophosphorylase |
| WB | western blot |
| MPRO | protoplasts from mock-inoculated healthy leaves |
| RPRO | protoplasts from RSV-infected leaves |
| MCHL | chloroplasts isolated from protoplasts from mock-inoculated healthy leaves |
| RCHL | chloroplasts isolated from protoplasts from RSV-infected leaves |
References
- Yeam, I. Current advances and prospectus of viral resistance in horticultural crops. Hortic. Environ. Biotechnol. 2016, 57, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Nelson, R.S.; Citovsky, V. Plant viruses. Invaders of cells and pirates of cellular pathways. Plant Physiol. 2005, 138, 1809–1814. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhao, J.; Chen, T.; Liu, Q.; Zhang, H.; Wang, Y.; Hong, Y.; Xiao, F.; Zhang, L.; Shen, Q.; et al. Type I J-domain NbMIP1 proteins are required for both tobacco mosaic virus infection and plant innate immunity. PLoS Pathog. 2013, 9, e1003659. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Kumar, S.; Singh, L.; Hallan, V. Movement Protein of Cucumber Mosaic Virus Associates with Apoplastic Ascorbate Oxidase. PLoS ONE 2016, 11, e0163320. [Google Scholar] [CrossRef] [PubMed]
- Dardick, C. Comparative expression profiling of Nicotiana benthamiana leaves systemically infected with three fruit tree viruses. Mol. Plant-Microbe Interact. 2007, 20, 1004–1017. [Google Scholar] [CrossRef]
- Shi, B.; Lin, L.; Wang, S.; Guo, Q.; Zhou, H.; Rong, L.; Li, J.; Peng, J.; Lu, Y.; Zheng, H.; et al. Identification and regulation of host genes related to Rice stripe virus symptom production. New Phytol. 2016, 209, 1106–1119. [Google Scholar] [CrossRef]
- Waliullah, S.; Mochizuki, T.; Sekine, K.T.; Atsumi, G.; Ali, M.E.; Yaeno, T.; Yamaoka, N.; Nishiguchi, M.; Kobayashi, K. Artificial induction of a plant virus protein in transgenic tobacco provides a synchronous system for analyzing the process of leaf chlorosis. Physiol. Mol. Plant Pathol. 2014, 88, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Bhor, S.A.; Tateda, C.; Mochizuki, T.; Sekine, K.T.; Yaeno, T.; Yamaoka, N.; Nishiguchi, M.; Kobayashi, K. Inducible transgenic tobacco system to study the mechanisms underlying chlorosis mediated by the silencing of chloroplast heat shock protein 90. Virusdisease 2017, 28, 1–12. [Google Scholar] [CrossRef]
- Balachandran, S.; Osmond, C.B.; Daley, P.F. Diagnosis of the earliest strain-specific interactions between tobacco mosaic virus and chloroplasts of tobacco leaves in vivo by means of chlorophyll fluorescence imaging. Plant Physiol. 1994, 104, 1059–1065. [Google Scholar] [CrossRef]
- Wang, S.; Cui, W.; Wu, X.; Yuan, Q.; Zhao, J.; Zheng, H.; Lu, Y.; Peng, J.; Lin, L.; Chen, J.; et al. Suppression of nbe-miR166h-p5 attenuates leaf yellowing symptoms of potato virus X on Nicotiana benthamiana and reduces virus accumulation. Mol. Plant Pathol. 2018, 19, 2384–2396. [Google Scholar] [CrossRef]
- Otulak, K.; Chouda, M.; Bujarski, J.; Garbaczewska, G. The evidence of tobacco rattle virus impact on host plant organelles ultrastructure. Micron 2015, 70, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.; Folimonova, S.Y.; Cole, A.B.; Ballard, K.D.; Lei, Z.; Watson, B.S.; Sumner, L.W.; Nelson, R.S. Influence of host chloroplast proteins on Tobacco mosaic virus accumulation and intercellular movement. Plant Physiol. 2013, 161, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Ogata, Y.; Hirata, Y.; Ohki, S.T. Quantitative transcriptional changes associated with chlorosis severity in mosaic leaves of tobacco plants infected with cucumber mosaic virus. Mol. Plant Pathol. 2014, 15, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Das, P.P.; Lin, Q.; Wong, S.-M. Comparative proteomics of Tobacco mosaic virus-infected Nicotiana tabacum plants identified major host proteins involved in photosystems and plant defence. J. Proteom. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mwaba, I.; Rey, M.E.C. Nitric oxide associated protein 1 is associated with chloroplast perturbation and disease symptoms in Nicotiana benthamiana infected with South African cassava mosaic virus. Virus Res. 2017, 238, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, X.; Hong, Y.; Liu, Y. Chloroplast in Plant-Virus Interaction. Front. Microbiol. 2016, 7, 1565. [Google Scholar] [CrossRef]
- Xu, K.; Nagy, P.D. Dissecting virus-plant interactions through proteomics approaches. Curr. Proteom. 2010, 7, 316–327. [Google Scholar] [CrossRef]
- Manfre, A.; Glenn, M.; Nuñez, A.; Moreau, R.A.; Dardick, C. Light quantity and photosystem function mediate host susceptibility to Turnip mosaic virus via a salicylic acid-independent mechanism. Mol. Plant-Microbe Interact. 2011, 24, 315–327. [Google Scholar] [CrossRef]
- Li, Y.; Cui, H.; Cui, X.; Wang, A. The altered photosynthetic machinery during compatible virus infection. Curr. Opin. Virol. 2016, 17, 19–24. [Google Scholar] [CrossRef]
- Revers, F.; García, J.A. Chapter Three—Molecular Biology of Potyviruses. In Advances in Virus Research; Maramorosch, K., Mettenleiter, T.C., Eds.; Academic Press, Elsevier Inc.: Amsterdam, The Netherlands, 2015; Volume 92, pp. 101–199. [Google Scholar]
- Cheng, Y.-Q.; Liu, Z.-M.; Xu, J.; Zhou, T.; Wang, M.; Chen, Y.-T.; Li, H.-F.; Fan, Z.-F. HC-Pro protein of sugar cane mosaic virus interacts specifically with maize ferredoxin-5 in vitro and in planta. J. Gen. Virol. 2008, 89, 2046–2054. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, D.; Gnanasekaran, P.; Kumar, R.K.; Kushwaha, N.K.; Sharma, V.K.; Yusuf, M.A.; Chakraborty, S. A geminivirus betasatellite damages the structural and functional integrity of chloroplasts leading to symptom formation and inhibition of photosynthesis. J. Exp. Bot. 2015, 66, 5881–5895. [Google Scholar] [CrossRef] [Green Version]
- Zarzyńska-Nowak, A.; Jeżewska, M.; Hasiów-Jaroszewska, B.; Zielińska, L. A comparison of ultrastructural changes of barley cells infected with mild and aggressive isolates of barley stripe mosaic virus. J. Plant Dis. Prot. 2015, 122, 153–160. [Google Scholar] [CrossRef]
- Tu, Y.; Jin, Y.; Ma, D.; Li, H.; Zhang, Z.; Dong, J.; Wang, T. Interaction between PVY HC-Pro and the NtCF1β-subunit reduces the amount of chloroplast ATP synthase in virus-infected tobacco. Sci. Rep. 2015, 5, 15605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Li, Y.; Shi, M.; Zhang, N.; Wu, G.; Li, T.; Qing, L.; Zhou, C. In vitro binding and bimolecular fluorescence complementation assays suggest an interaction between tomato mosaic virus coat protein and tobacco chloroplast ferredoxin I. Arch. Virol. 2013, 158, 2611–2615. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhou, T.; Hong, Y.; Fan, Z.; Li, H. Decreased level of ferredoxin I in tobacco mosaic virus-infected tobacco is associated with development of the mosaic symptom. Physiol. Mol. Plant Pathol. 2008, 72, 39–45. [Google Scholar] [CrossRef]
- Bobik, K.; Burch-Smith, T.M. Chloroplast signaling within, between and beyond cells. Front. Plant Sci. 2015, 6, 781. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, M.; Kim, B.S.; Hutchenswilliams, H.M.; Loeschfries, L.S. The photosystem II oxygen-evolving complex protein PsbP interacts with the coat protein of Alfalfa mosaic virus and inhibits virus replication. Mol. Plant-Microbe Interact. 2014, 27, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Burch-Smith, T.M.; Zambryski, P.C. Regulation of Plasmodesmal Transport and Modification of Plasmodesmata During Development and Following Infection by Viruses and Viral Proteins. In Plant-Virus Interactions: Molecular Biology, Intra- and Intercellular Transport; Kleinow, T., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 87–122. [Google Scholar]
- Zhao, J.; Liu, Q.; Zhang, H.; Jia, Q.; Hong, Y.; Liu, Y. The RubisCO small subunit is involved in Tobamovirus movement and Tm-22-mediated extreme resistance. Plant Physiol. 2013, 161, 374–383. [Google Scholar] [CrossRef]
- Cheng, S.-F.; Huang, Y.-P.; Chen, L.-H.; Hsu, Y.-H.; Tsai, C.-H. Chloroplast phosphoglycerate kinase is involved in the targeting of bamboo mosaic virus to chloroplasts in Nicotiana benthamiana plants. Plant Physiol. 2013, 163, 1598–1608. [Google Scholar] [CrossRef]
- Seo, E.Y.; Nam, J.; Kim, H.S.; Park, Y.H.; Hong, S.M.; Lakshman, D.; Bae, H.; Hammond, J.; Lim, H.S. Selective Interaction Between Chloroplast β-ATPase and TGB1L88 Retards Severe Symptoms Caused by Alternanthera mosaic virus Infection. Plant Pathol. J. 2014, 30, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Cai, Z.; Hu, T.; Yao, H.; Wang, L.; Dong, N.; Wang, B.; Ru, Z.; Zhai, W. Genetic Analysis and Molecular Mapping of a Novel Gene Conferring Resistance to Rice Stripe Virus. Plant Mol. Biol. Report. 2010, 28, 512–518. [Google Scholar] [CrossRef]
- Cho, W.K.; Lian, S.; Kim, S.M.; Park, S.H.; Kim, K.H. Current Insights into Research on Rice stripe virus. Plant Pathol. J. 2013, 29, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toriyama, S. Rice stripe virus: Prototype of a new group of viruses that replicate in plants and insects. Microbiol. Sci. 1986, 3, 347–351. [Google Scholar] [PubMed]
- Lee, S.B.; Hur, Y.J.; Lee, J.H.; Kwon, T.; Shin, D.; Kim, T.H.; Han, S.I.; Cho, J.H.; Yoon, Y.N.; Kiswara, G. Molecular mapping of a quantitative trait locus qSTV11Z harbouring rice stripe virus resistance gene, Stv-b. Plant Breed. 2017, 136, 61–66. [Google Scholar] [CrossRef]
- Wang, B.; Hajano, J.-U.-D.; Ren, Y.; Lu, C.; Wang, X. iTRAQ-based quantitative proteomics analysis of rice leaves infected by Rice stripe virus reveals several proteins involved in symptom formation. Virol. J. 2015, 12, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Smith, J.W.; Huang, C.M. Mass spectrometry-based label-free quantitative proteomics. J. Biomed. Biotechnol. 2010, 2010, 840518. [Google Scholar] [CrossRef]
- Bond, N.J.; Shliaha, P.V.; Lilley, K.S.; Gatto, L. Improving qualitative and quantitative performance for MS(E)-based label-free proteomics. J. Proteome Res. 2013, 12, 2340–2353. [Google Scholar] [CrossRef]
- Falk, B.W.; Tsai, J.H. Biology and molecular biology of viruses in the genus Tenuivirus. Annu. Rev. Phytopathol. 1998, 36, 139–163. [Google Scholar] [CrossRef]
- Lian, S.; Jonson, M.G.; Cho, W.K.; Choi, H.S.; Je, Y.H.; Kim, K.H. Generation of Antibodies Against Rice stripe virus Proteins Based on Recombinant Proteins and Synthetic Polypeptides. Plant Pathol. J. 2011, 27, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Hibino, H. Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 2003, 34, 249–274. [Google Scholar] [CrossRef]
- Cho, W.K.; Lian, S.; Kim, S.-M.; Seo, B.Y.; Jung, J.K.; Kim, K.-H. Time-Course RNA-Seq Analysis Reveals Transcriptional Changes in Rice Plants Triggered by Rice stripe virus Infection. PLoS ONE 2015, 10, e0136736. [Google Scholar] [CrossRef] [PubMed]
- Xiong, R.; Wu, J.; Zhou, Y.; Zhou, X. Identification of a Movement Protein of the Tenuivirus Rice Stripe Virus. J. Virol. 2008, 82, 12304–12311. [Google Scholar] [CrossRef] [PubMed]
- Peltier, J.-B.; Emanuelsson, O.; Kalume, D.E.; Ytterberg, J.; Friso, G.; Rudella, A.; Liberles, D.A.; Söderberg, L.; Roepstorff, P.; von Heijne, G.; et al. Central Functions of the Lumenal and Peripheral Thylakoid Proteome of Arabidopsis Determined by Experimentation and Genome-Wide Prediction. Plant Cell 2002, 14, 211–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, A.J.K.; Ohlrogge, J.B. The Predicted Candidates of Arabidopsis Plastid Inner Envelope Membrane Proteins and Their Expression Profiles. Plant Physiol. 2002, 130, 823–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, L.J.; Imre, K.M.; Hall, D.A.; Last, R.L. Analysis of Essential Arabidopsis Nuclear Genes Encoding Plastid-Targeted Proteins. PLoS ONE 2013, 8, e73291. [Google Scholar] [CrossRef]
- Ajjawi, I.; Lu, Y.; Savage, L.J.; Bell, S.M.; Last, R.L. Large-Scale Reverse Genetics in Arabidopsis: Case Studies from the Chloroplast 2010 Project. Plant Physiol. 2010, 152, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Myouga, F.; Akiyama, K.; Tomonaga, Y.; Kato, A.; Sato, Y.; Kobayashi, M.; Nagata, N.; Sakurai, T.; Shinozaki, K. The Chloroplast Function Database II: A Comprehensive Collection of Homozygous Mutants and Their Phenotypic/Genotypic Traits for Nuclear-Encoded Chloroplast Proteins. Plant Cell Physiol. 2013, 54, e2. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting Subcellular Localization of Proteins Based on their N-terminal Amino Acid Sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef]
- Jin, X.; Jiang, Z.; Zhang, K.; Wang, P.; Cao, X.; Yue, N.; Wang, X.; Zhang, X.; Li, Y.; Li, D.; et al. Three-Dimensional Analysis of Chloroplast Structures Associated with Virus Infection. Plant Physiol. 2018, 176, 282–294. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Chakraborty, S. Chloroplast: The Trojan horse in plant–virus interaction. Mol. Plant Pathol. 2017, 19, 504–518. [Google Scholar] [CrossRef]
- Roberts, P.L.; Wood, K.R. Effects of a severe (P6) and a mild (W) strain of cucumber mosaic virus on tobacco leaf chlorophyll, starch and cell ultrastructure. Physiol. Plant Pathol. 1982, 21, 31IN3333–32IN3537. [Google Scholar] [CrossRef]
- Lei, R.; Jiang, H.; Hu, F.; Yan, J.; Zhu, S. Chlorophyll fluorescence lifetime imaging provides new insight into the chlorosis induced by plant virus infection. Plant Cell Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lehto, K.; Tikkanen, M.; Hiriart, J.-B.; Paakkarinen, V.; Aro, E.-M. Depletion of the photosystem II core complex in mature tobacco leaves infected by the flavum strain of tobacco mosaic virus. Mol. Plant-Microbe Interact. 2003, 16, 1135–1144. [Google Scholar] [CrossRef]
- Reinero, A.; Beachy, R.N. Association of TMV coat protein with chloroplast membranes in virus-infected leaves. Plant Mol. Biol. 1986, 6, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, Y.; Sun, X.; Qian, W.; Zhang, D.; Qiu, B. Characterization of a specific interaction between IP-L, a tobacco protein localized in the thylakoid membranes, and Tomato mosaic virus coat protein. Biochem. Biophys. Res. Commun. 2008, 374, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Wu, J.; Lu, L.; Xu, Y.; Zhou, X. Interaction between rice stripe virus disease-specific protein and host PsbP enhances virus symptoms. Mol. Plant 2014, 7, 691–708. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Bueno, M.L.; Rahoutei, J.; Sajnani, C.; García-Luque, I.; Barón, M. Proteomic analysis of the oxygen-evolving complex of photosystem II under biotec stress: Studies on Nicotiana benthamiana infected with tobamoviruses. Proteomics 2004, 4, 418–425. [Google Scholar] [CrossRef]
- Seidler, A. The extrinsic polypeptides of Photosystem II. Biochim Biophys Acta Bioenerg. 1996, 35–60. [Google Scholar] [CrossRef]
- Lindbeck, A.G.C.; Dawson, W.O.; Thomson, W.W. Coat protein-related polypeptides from in vitro tobacco mosaic virus coat protein mutants do not accumulate in the chloroplasts of directly inoculated leaves. Mol. Plant-Microbe Interact. 1991, 4, 89–94. [Google Scholar] [CrossRef]
- Lindbeck, A.G.C.; Lewandowski, D.J.; Culver, J.N.; Thomson, W.W.; Dawson, W.O. Mutant coat protein of tobacco mosaic virus induces acute chlorosis in expanded and developing tobacco leaves. Mol. Plant-Microbe Interact. 1992, 5, 235–241. [Google Scholar] [CrossRef]
- Ohnishi, J.; Hirai, K.; Kanda, A.; Usugi, T.; Meshi, T.; Tsuda, S. The coat protein of tomato mosaic virus L11Y is associated with virus-induced chlorosis on infected tobacco plants. J. Gen. Plant Pathol. 2009, 75, 297–306. [Google Scholar] [CrossRef]
- Kopek, B.G.; Perkins, G.; Miller, D.J.; Ellisman, M.H.; Ahlquist, P. Three-Dimensional Analysis of a Viral RNA Replication Complex Reveals a Virus-Induced Mini-Organelle. PLoS Biol. 2007, 5, e220. [Google Scholar] [CrossRef] [PubMed]
- Grangeon, R.; Agbeci, M.; Chen, J.; Grondin, G.; Zheng, H.; Laliberté, J.-F. Impact on the Endoplasmic Reticulum and Golgi Apparatus of Turnip Mosaic Virus Infection. J. Virol. 2012, 86, 9255–9265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, P.; Brown, J. Nucleoli: Composition, Function, and Dynamics. Plant Physiol. 2012, 158, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, A.G.; Savenkov, E.I. Factors involved in the systemic transport of plant RNA viruses: The emerging role of the nucleus. J. Exp. Bot. 2014, 65, 1689–1697. [Google Scholar] [CrossRef]
- Zheng, L.; Du, Z.; Lin, C.; Mao, Q.; Wu, K.; Wu, J.; Wei, T.; Wu, Z.; Xie, L. Rice stripe tenuivirus p2 may recruit or manipulate nucleolar functions through an interaction with fibrillarin to promote virus systemic movement. Mol. Plant Pathol. 2015, 16, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Ryabov, E.V.; Kalinina, N.O.; Rakitina, D.V.; Gillespie, T.; Macfarlane, S.; Haupt, S.; Brown, J.W.; Taliansky, M. Cajal bodies and the nucleolus are required for a plant virus systemic infection. EMBO J. 2007, 26, 2169–2179. [Google Scholar] [CrossRef] [Green Version]
- McCartney, A.W.; Greenwood, J.S.; Fabian, M.R.; White, K.A.; Mullen, R.T. Localization of the Tomato Bushy Stunt Virus Replication Protein p33 Reveals a Peroxisome-to-Endoplasmic Reticulum Sorting Pathway. Plant Cell 2005, 17, 3513–3531. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, T.; Hirai, K.; Kanda, A.; Ohnishi, J.; Ohki, T.; Tsuda, S. Induction of necrosis via mitochondrial targeting of Melon necrotic spot virus replication protein p29 by its second transmembrane domain. Virology 2009, 390, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Weber-Lotfi, F.; Dietrich, A.; Russo, M.; Rubino, L. Mitochondrial Targeting and Membrane Anchoring of a Viral Replicase in Plant and Yeast Cells. J. Virol. 2002, 76, 10485–10496. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, J.-F.; Sanfaçon, H. Cellular Remodeling During Plant Virus Infection. Annu. Rev. Phytopathol. 2010, 48, 69–91. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.-M.; Chen, S.; Payton, M.; Dickman, M.B.; Verchot, J. TGBp3 triggers the unfolded protein response and SKP1-dependent programmed cell death. Mol. Plant Pathol. 2013, 14, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Dickman, M.B.; Whitham, S.A.; Payton, M.; Verchot, J. The Unfolded Protein Response Is Triggered by a Plant Viral Movement Protein. Plant Physiol. 2011, 156, 741–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Yang, D.; Xie, L.; Sun, L.; Zhang, S.; Zhu, Q.; Li, J.; Wang, X.; Chen, J. Rice black-streaked dwarf virus P10 induces membranous structures at the ER and elicits the unfolded protein response in Nicotiana benthamiana. Virology 2013, 447, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chen, H.; Brandizzi, F.; Verchot, J.; Wang, A. The UPR Branch IRE1-bZIP60 in Plants Plays an Essential Role in Viral Infection and Is Complementary to the Only UPR Pathway in Yeast. PLoS Genet. 2015, 11, e1005164. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yin, M.; Wang, X.; Chen, B.; Yang, X.; Peng, J.; Zheng, H.; Zhao, J.; Lin, L.; Yu, C.; et al. The unfolded protein response and programmed cell death are induced by expression of Garlic virus X p11 in Nicotiana benthamiana. J. Gen. Virol. 2016, 97, 1462–1468. [Google Scholar] [CrossRef] [Green Version]
- Ganusova, E.E.; Burch-Smith, T.M. Review: Plant-pathogen interactions through the plasmodesma prism. Plant Sci. 2018. [Google Scholar] [CrossRef]
- Amari, K.; Lerich, A.; Schmitt-Keichinger, C.; Dolja, V.V.; Ritzenthaler, C. Tubule-Guided Cell-to-Cell Movement of a Plant Virus Requires Class XI Myosin Motors. PLoS Pathog. 2011, 7, e1002327. [Google Scholar] [CrossRef]
- Thomas, C.L.; Bayer, E.M.; Ritzenthaler, C.; Fernandez-Calvino, L.; Maule, A.J. Specific Targeting of a Plasmodesmal Protein Affecting Cell-to-Cell Communication. PLoS Biol. 2008, 6, e7. [Google Scholar] [CrossRef]
- den Hollander, P.W.; Kieper, S.N.; Borst, J.W.; van Lent, J.W.M. The role of plasmodesma-located proteins in tubule-guided virus transport is limited to the plasmodesmata. Arch. Virol. 2016, 161, 2431–2440. [Google Scholar] [CrossRef] [Green Version]
- Amari, K.; Boutant, E.; Hofmann, C.; Schmitt-Keichinger, C.; Fernandez-Calvino, L.; Didier, P.; Lerich, A.; Mutterer, J.; Thomas, C.L.; Heinlein, M.; et al. A Family of Plasmodesmal Proteins with Receptor-Like Properties for Plant Viral Movement Proteins. PLoS Pathog. 2010, 6, e1001119. [Google Scholar] [CrossRef]
- Sandhu, D.; Atkinson, T.; Noll, A.; Johnson, C.; Espinosa, K.; Boelter, J.; Abel, S.; Dhatt, B.K.; Barta, T.; Singsaas, E. Soybean proteins GmTic110 and GmPsbP are crucial for chloroplast development and function. Plant Sci. 2016, 252, 76–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, P.; Sy, D.A.; Pilgrim, M.L.; Parry, D.H.; Nussaume, L.; Hoffman, N.E. Arabidopsis mutants lacking the 43- and 54-kilodalton subunits of the chloroplast signal recognition particle have distinct phenotypes. Plant Physiol. 1999, 121, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Luo, X.; Hu, B.; Yong, W.; Xie, J. YGL138 (t), encoding a putative signal recognition particle 54 kDa protein, is involved in chloroplast development of rice. Rice 2013, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Navarro, B.; Gisel, A.; Rodio, M.E.; Delgado, S.; Flores, R.; Di, S.F. Small RNAs containing the pathogenic determinant of a chloroplast-replicating viroid guide the degradation of a host mRNA as predicted by RNA silencing. Plant J. 2012, 70, 991–1003. [Google Scholar] [CrossRef]
- Hörmann, F. The Chloroplast Protein Import Machinery: A Review. Methods Mol. Biol. 2010, 619, 307–321. [Google Scholar]
- Erika, K.B.; Philipp, B.J.; Jürgen, S.; Bettina, B. Tic20 forms a channel independent of Tic110 in chloroplasts. BMC Plant Biol. 2011, 11, 133. [Google Scholar]
- Inaba, A.-H.; Li, B.E. Arabidopsis tic110 is essential for the assembly and function of the protein import machinery of plastids. Plant Cell 2005, 17, 1482–1496. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.E.; Yeung, C.; Babaeirad, R.; Zhao, R. Cosuppression of the chloroplast localized molecular chaperone HSP90.5 impairs plant development and chloroplast biogenesis in Arabidopsis. BMC Res. Notes 2014, 7, 643. [Google Scholar] [CrossRef]
- Inoue, H.; Li, M.; Schnell, D.J. An essential role for chloroplast heat shock protein 90 (Hsp90C) in protein import into chloroplasts. Proc. Natl. Acad. Sci. USA 2013, 110, 3173–3178. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.E. The Role of Chloroplast Localized Molecular Chaperone HSP90 in Arabidopsis thaliana Development. Master’s Thesis, Department of Cell & Systems Biology, University of Toronto, Toronto, ON, Canada, 2012. [Google Scholar]
- Fei, Y.; Zhang, H.M.; Adams, M.J.; Jian, Y.; Peng, J.J.; Antoniw, J.F.; Zhou, Y.J.; Chen, J.P. Characterization of siRNAs derived from rice stripe virus in infected rice plants by deep sequencing. Arch. Virol. 2010, 155, 935–940. [Google Scholar]
- Jiang, S.; Lu, Y.; Li, K.; Lin, L.; Zheng, H.; Yan, F.; Chen, J. Heat shock protein 70 is necessary for Rice stripe virus infection in plants. Mol. Plant Pathol. 2014, 15, 907–917. [Google Scholar] [PubMed]
- Fitzpatrick, L.M.; Keegstra, K. A method for isolating a high yield of Arabidopsis chloroplasts capable of efficient import of precursor proteins. Plant J. 2001, 27, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Wang, Z.; Cheng, J.; Zhao, W.; Li, X.; Wang, H.; Zhang, Z.; Sui, X. An efficient cucumber (Cucumis sativus L.) protoplast isolation and transient expression system. Sci. Hortic. 2013, 150, 206–212. [Google Scholar] [CrossRef]
- Rao, A.L. Preparation and inoculation of mesophyll protoplasts from monocotyledenous and dicotyledenous hosts. Curr. Protoc. Microbiol. 2007. [Google Scholar] [CrossRef]
- Larkin, P.J. Purification and viability determinations of plant protoplasts. Planta 1976, 128, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Kunst, L. Preparation of physiologically active chloroplasts from Arabidopsis. Methods Mol. Biol. 1998, 82, 43–48. [Google Scholar]
- Shao, J.; Zhang, Y.; Yu, J.; Guo, L.; Ding, Y. Isolation of thylakoid membrane complexes from rice by a new double-strips BN/SDS-PAGE and bioinformatics prediction of stromal ridge subunits interaction. PLoS ONE 2011, 6, e20342. [Google Scholar] [CrossRef]
- Qureshi, M.I.; D’Amici, G.M.; Fagioni, M.; Rinalducci, S.; Zolla, L. Iron stabilizes thylakoid protein-pigment complexes in Indian mustard during Cd-phytoremediation as revealed by BN-SDS-PAGE and ESI-MS/MS. J. Plant Physiol. 2010, 167, 761–770. [Google Scholar] [CrossRef]
- Chatterjee, M.; Gupta, S.; Bhar, A.; Das, S. Optimization of an Efficient Protein Extraction Protocol Compatible with Two-Dimensional Electrophoresis and Mass Spectrometry from Recalcitrant Phenolic Rich Roots of Chickpea (Cicer arietinum L.). Int. J. Proteom. 2012, 2012, 536963. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt AC | Protein Names | GO IDs |
|---|---|---|
| K4C8H3 | Uncharacterized protein | GO:0003842; GO:0004028; GO:0004029; GO:0005739; GO:0008270; GO:0009507; GO:0009651; GO:0010133; GO:0050897; GO:0072593 |
| Q711R1 | Allene oxide cyclase | GO:0009507; GO:0046423 |
| K4B2I9 | Uncharacterized protein | GO:0005524; GO:0006457 |
| K4BT58 | Uncharacterized protein | GO:0005509; GO:0009534; GO:0009570; GO:0009654; GO:0015979; GO:0019898; GO:0031977 |
| K4B778 | Uncharacterized protein | GO:0005741; GO:0008308 |
| A8UDS9 | Copper/zinc superoxide dismutase | GO:0006801; GO:0046872; GO:0055114 |
| A9CM21 | Voltage-dependent anion channel | GO:0005741; GO:0008308 |
| A0A075F1U0 | Constitutive plastid-lipid associated protein | |
| K4C647 | Uncharacterized protein | GO:0009570; GO:0009941; GO:0016491 |
| K4CNE8 | ATP-dependent Clp protease proteolytic subunit | GO:0000302; GO:0004252; GO:0009570; GO:0009941; GO:0010468 |
| K4B858 | Uncharacterized protein | GO:0005622 |
| Q10712 | Leucine aminopeptidase 1 | GO:0004177; GO:0008235; GO:0009507; GO:0030145 |
| K4B3J8 | Uncharacterized protein | GO:0008836; GO:0009089 |
| K4ASX2 | Uncharacterized protein | GO:0005623; GO:0045454; GO:0051920 |
| P37222 | NADP-dependent malic enzyme | GO:0004471; GO:0004473; GO:0006108; GO:0008948; GO:0009507; GO:0046872; GO:0051287 |
| A0A140G1U2 | ATP-dependent Clp protease proteolytic subunit | GO:0004252; GO:0009536 |
| K4AZT2 | Uncharacterized protein | GO:0009579; GO:0016021 |
| K4DC13 | Uncharacterized protein | GO:0005524; GO:0005759; GO:0006986; GO:0009507; GO:0046872; GO:0046914; GO:0051082; GO:0051085; GO:0051087; GO:1901671 |
| K4BXN9 | Uncharacterized protein | GO:0005524; GO:0005739; GO:0005774; GO:0006457; GO:0009408; GO:0009414; GO:0009570; GO:0009651; GO:0009704; GO:0009941; GO:0010157; GO:0045037 |
| K4D4L5 | Uncharacterized protein | GO:0005886; GO:0016655 |
| K4C841 | Uncharacterized protein | GO:0008152; GO:0016597 |
| K4BFT9 | Uncharacterized protein | GO:0016021 |
| K4B413 | Uncharacterized protein | GO:0000302; GO:0004130; GO:0005739; GO:0005774; GO:0005777; GO:0009506; GO:0009507; GO:0009735; GO:0009941; GO:0016021; GO:0020037; GO:0034599; GO:0042744 |
| K4BBZ1 | Uncharacterized protein | |
| A7U630 | Chloroplast methionine sulfoxide reductase B1 | GO:0006979; GO:0030091; GO:0033743 |
| K4CH79 | Uncharacterized protein | GO:0003735; GO:0005840; GO:0006412; GO:0009570; GO:0009941; GO:0032544 |
| Q8W3Z8 | Plastidic phosphoglucomutase | GO:0005975; GO:0016868 |
| K4CXW3 | Glutamyl-tRNA (Gln) amidotransferase subunit A | GO:0004040; GO:0005524; GO:0005739; GO:0009570; GO:0030956; GO:0032543; GO:0050567; GO:0070681 |
| K4BCV6 | Uncharacterized protein | GO:0004222; GO:0006518; GO:0009507; GO:0046872 |
| K4BCU7 | Uncharacterized protein | GO:0004747; GO:0006014 |
| K4CGD5 | Uncharacterized protein | GO:0003743; GO:0003924; GO:0005525; GO:0005622; GO:0006412; GO:0009570; GO:0009941 |
| K4BXX3 | Alpha-1,4 glucan phosphorylase | GO:0005737; GO:0005980; GO:0008184; GO:0030170 |
| K4BLP5 | Uncharacterized protein | |
| E5LBC4 | Prephenate aminotransferase | GO:0008483; GO:0009058; GO:0030170 |
| Q6T7F1 | Phosphoribosylaminoimidazole carboxylase | GO:0004638; GO:0005524; GO:0006189; GO:0046872 |
| K4C5B9 | Uroporphyrinogen decarboxylase | GO:0004853; GO:0005829; GO:0006782; GO:0006783; GO:0009570; GO:0009735; GO:0009941 |
| K4BJL2 | Uncharacterized protein | GO:0003735; GO:0005840; GO:0006412 |
| K4CPX9 | Uncharacterized protein | GO:0000027; GO:0003735; GO:0005762; GO:0006412; GO:0019843 |
| K4ASZ2 | Uncharacterized protein | GO:0003824; GO:0009058; GO:0030170 |
| K4BDV0 | Uncharacterized protein | GO:0000103; GO:0004020; GO:0004781 |
| K4ASJ9 | Uncharacterized protein | GO:0009535; GO:0009941 |
| K4BEB0 | Uncharacterized protein | GO:0000287; GO:0004614; GO:0005829; GO:0005978; GO:0006006; GO:0009570; GO:0009590; GO:0010319; GO:0019252; GO:0019388 |
| K4B075 | Uncharacterized protein | GO:0003723; GO:0004827; GO:0005524; GO:0005737; GO:0005739; GO:0006433; GO:0009553; GO:0009570; GO:0010109; GO:0017101; GO:0048316; GO:0048481 |
| K4BX77 | 3-isopropylmalate dehydrogenase | GO:0000287; GO:0003862; GO:0009098; GO:0009570; GO:0051287 |
| K4CJD3 | Uncharacterized protein | |
| A8UDS7 | ABA2 | GO:0016491; GO:0071949 |
| Q2MI78 | 30S ribosomal protein S18 | GO:0003735; GO:0006412; GO:0009507; GO:0019843; GO:0022627 |
| K4CR23 | Uncharacterized protein | GO:0003924; GO:0005525; GO:0006614; GO:0008312; GO:0009570; GO:0080085 |
| K4CNE7 | Uncharacterized protein | GO:0009535; GO:0022891; GO:0055085 |
| K4D5G2 | Uncharacterized protein | GO:0000287; GO:0005737; GO:0009570; GO:0009941; GO:0016791 |
| K4BU13 | Uncharacterized protein | GO:0000166; GO:0003729; GO:0009570 |
| K4BY59 | Uncharacterized protein | GO:0003824; GO:0009058; GO:0030170 |
| Q84RD7 | ZIP | GO:0015979; GO:0015995; GO:0046872; GO:0048529 |
| K4BPB0 | Uncharacterized protein | GO:0003735; GO:0006412; GO:0015934; GO:0022626; GO:0042254 |
| K4BXB1 | Uncharacterized protein | GO:0003735; GO:0006412; GO:0009570; GO:0009735; GO:0009941; GO:0015934 |
| K4CSN4 | Uncharacterized protein | GO:0009658; GO:0009941; GO:0016021; GO:0045037 |
| K4B2B2 | Phospho-2-dehydro-3-deoxyheptonate aldolase | GO:0003849; GO:0009073; GO:0009507 |
| A0A0H3U3I7 | Geranylgeranyl pyrophosphate synthase 5 | GO:0008299; GO:0016740 |
| K4AXX7 | Protein translocase subunit SecA | GO:0005524; GO:0006605; GO:0009658; GO:0009941; GO:0010109; GO:0015462; GO:0016020; GO:0017038 |
| K4C9K5 | Uncharacterized protein | GO:0006457; GO:0015031 |
| Q0MW94 | 1-deoxy-d-xylulose-5-phosphate reductoisomerase | GO:0008299; GO:0016853; GO:0030604; GO:0046872; GO:0070402 |
| K4BDU3 | Uncharacterized protein | GO:0009055; GO:0051536 |
| K9JIM2 | Heat shock protein 101 | GO:0005524 |
| K4BVZ1 | Uncharacterized protein | GO:0016021 |
| A0A0K1D9E3 | Granule-bound starch synthase I | |
| K4CHH4 | Uncharacterized protein | GO:0006457; GO:0015031 |
| GO ID | GO Term | p-Value | UniProt Accession | Protein Name |
|---|---|---|---|---|
| GO:0045036 | protein targeting to chloroplast | 0.00242 | K4CSN4 K4CR23 K4BXN9 | Tic110 cpSRP54 HSP90C |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Xu, J.; Chen, B.; Cui, W.; Zhou, Z.; Song, X.; Chen, Z.; Zheng, H.; Lin, L.; Peng, J.; et al. Characterization of Proteins Involved in Chloroplast Targeting Disturbed by Rice Stripe Virus by Novel Protoplast–Chloroplast Proteomics. Int. J. Mol. Sci. 2019, 20, 253. https://doi.org/10.3390/ijms20020253
Zhao J, Xu J, Chen B, Cui W, Zhou Z, Song X, Chen Z, Zheng H, Lin L, Peng J, et al. Characterization of Proteins Involved in Chloroplast Targeting Disturbed by Rice Stripe Virus by Novel Protoplast–Chloroplast Proteomics. International Journal of Molecular Sciences. 2019; 20(2):253. https://doi.org/10.3390/ijms20020253
Chicago/Turabian StyleZhao, Jinping, Jingjing Xu, Binghua Chen, Weijun Cui, Zhongjing Zhou, Xijiao Song, Zhuo Chen, Hongying Zheng, Lin Lin, Jiejun Peng, and et al. 2019. "Characterization of Proteins Involved in Chloroplast Targeting Disturbed by Rice Stripe Virus by Novel Protoplast–Chloroplast Proteomics" International Journal of Molecular Sciences 20, no. 2: 253. https://doi.org/10.3390/ijms20020253