Prostaglandin E2 Induces Skin Aging via E-Prostanoid 1 in Normal Human Dermal Fibroblasts


Abstract
1. Introduction
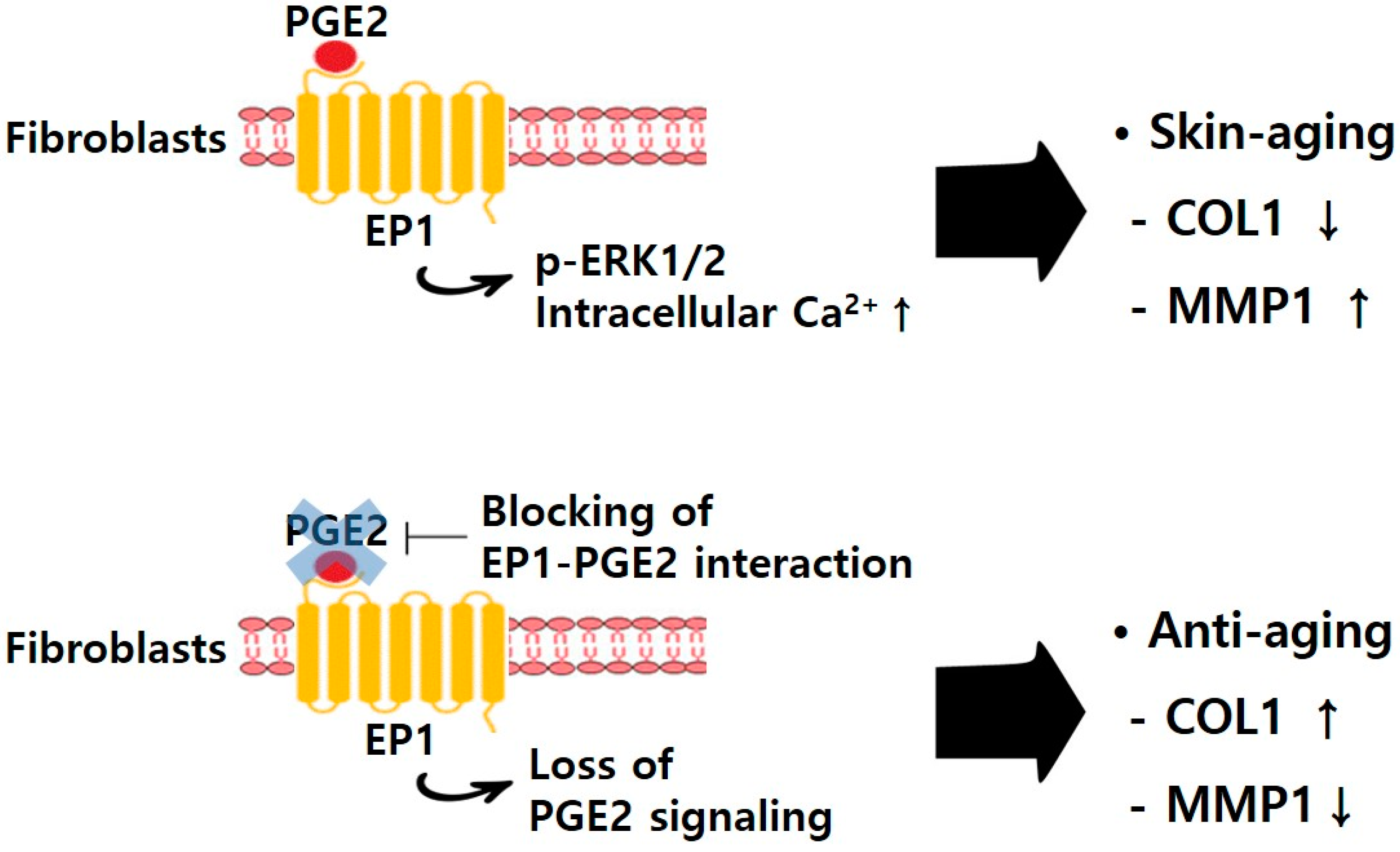
2. Results and Discussion
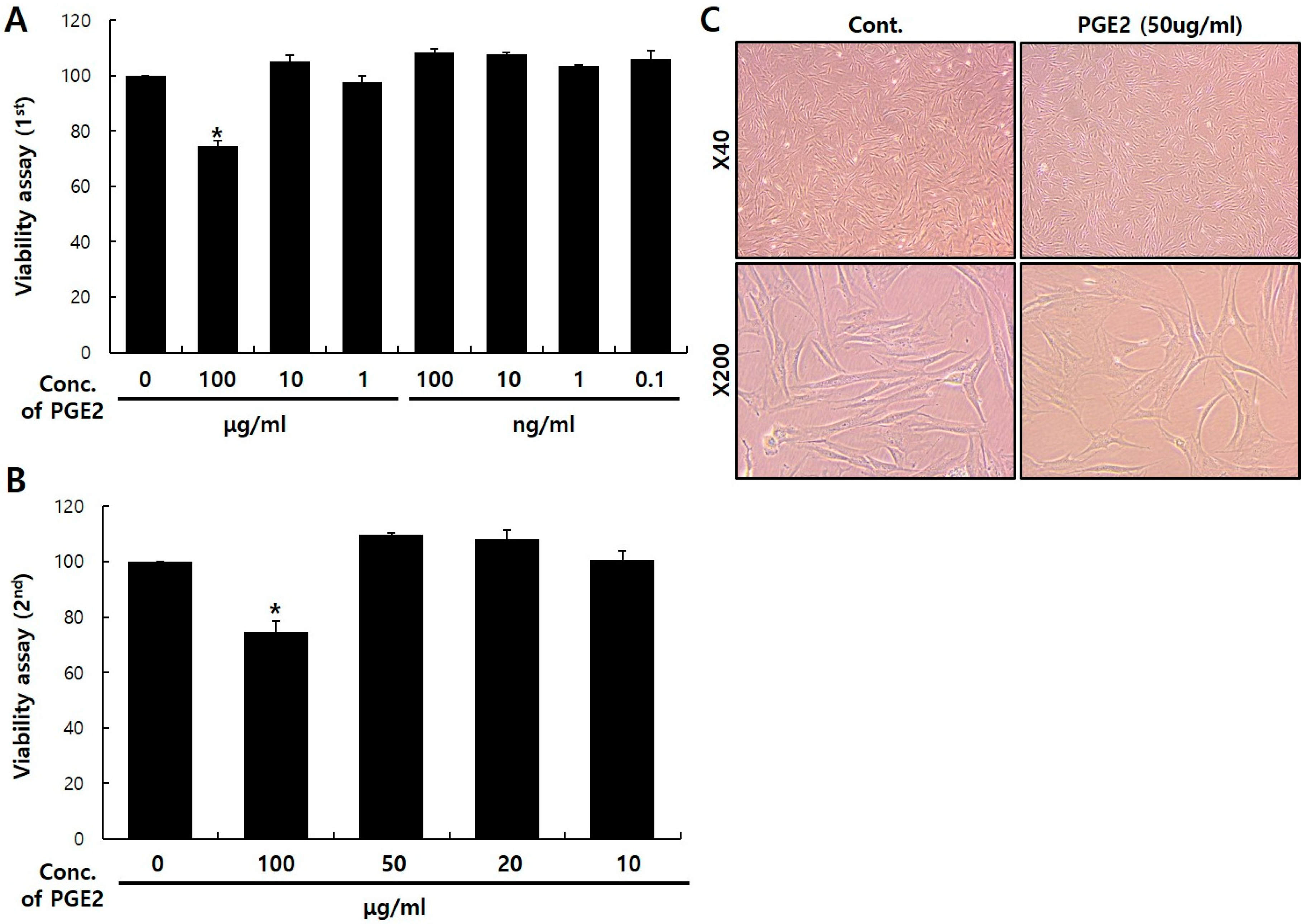
2.1. Viability of PGE2 in NHDFs
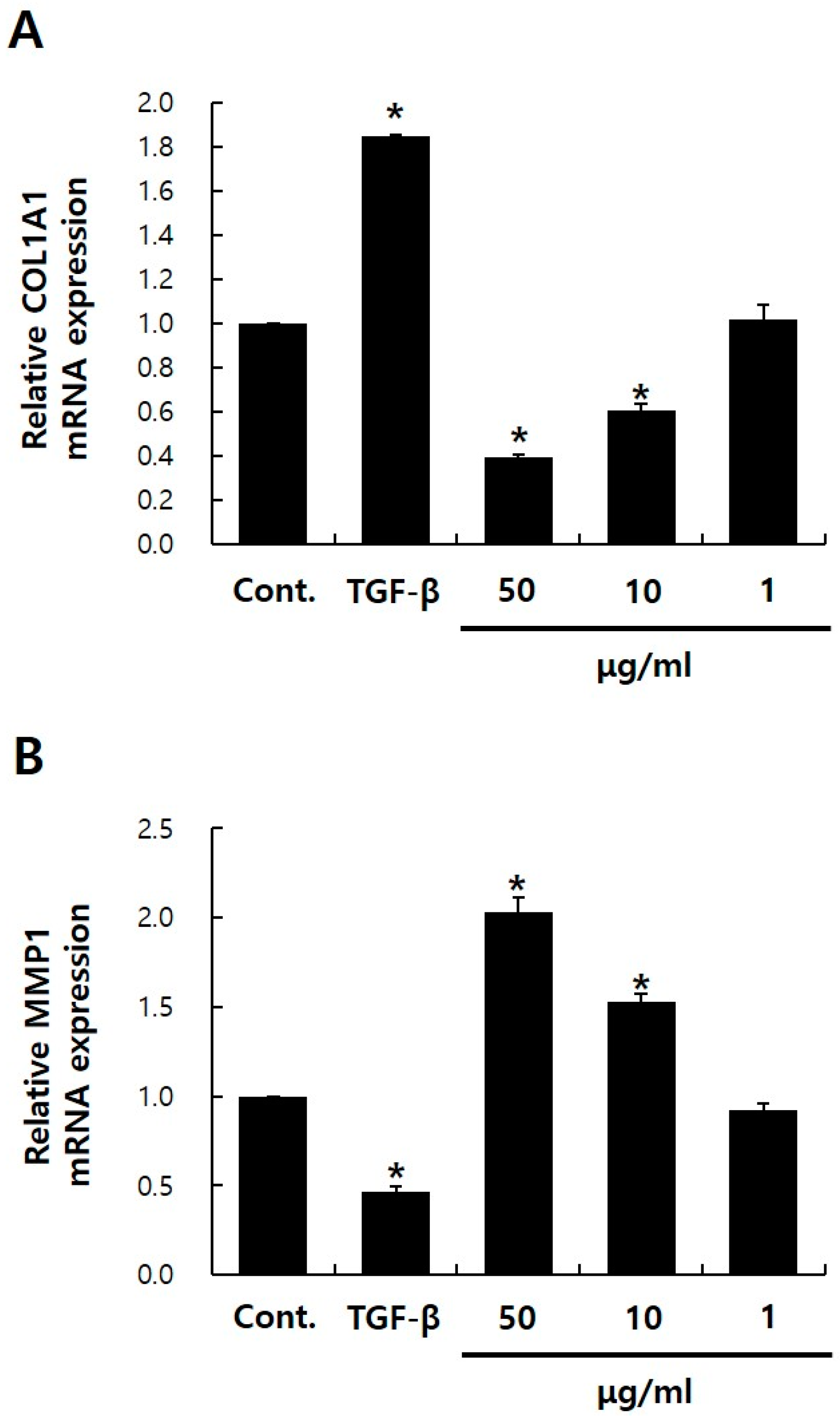
2.2. Effects of PGE2 on collagen, type I, alpha 1 (COL1A1) and MMP1 mRNA Expressions in NHDFs
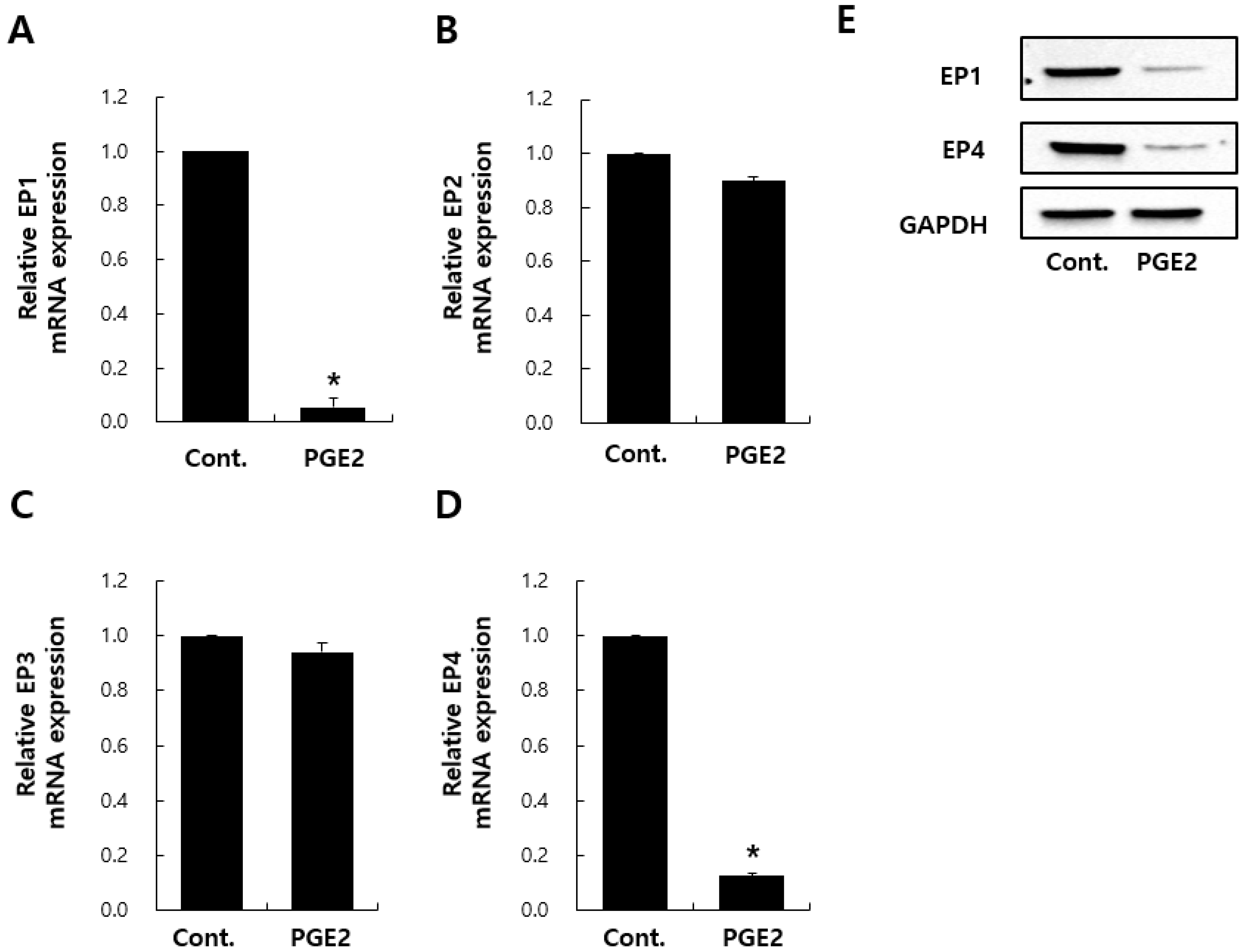
2.3. Effects of PGE2 on mRNA Expression–EP Receptors in NHDFs
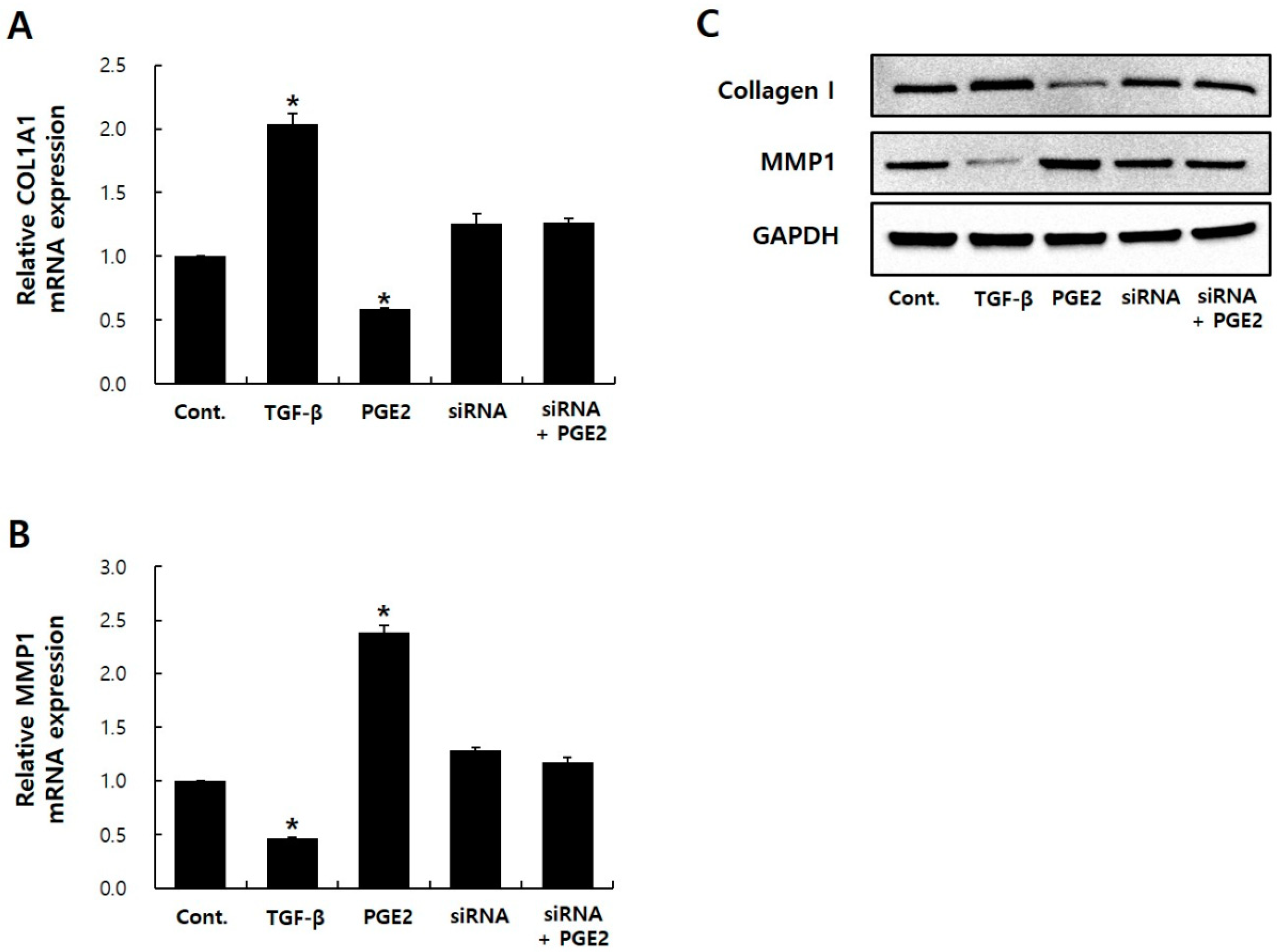
2.4. EP1 siRNA Treatment Hides PGE2 Effects
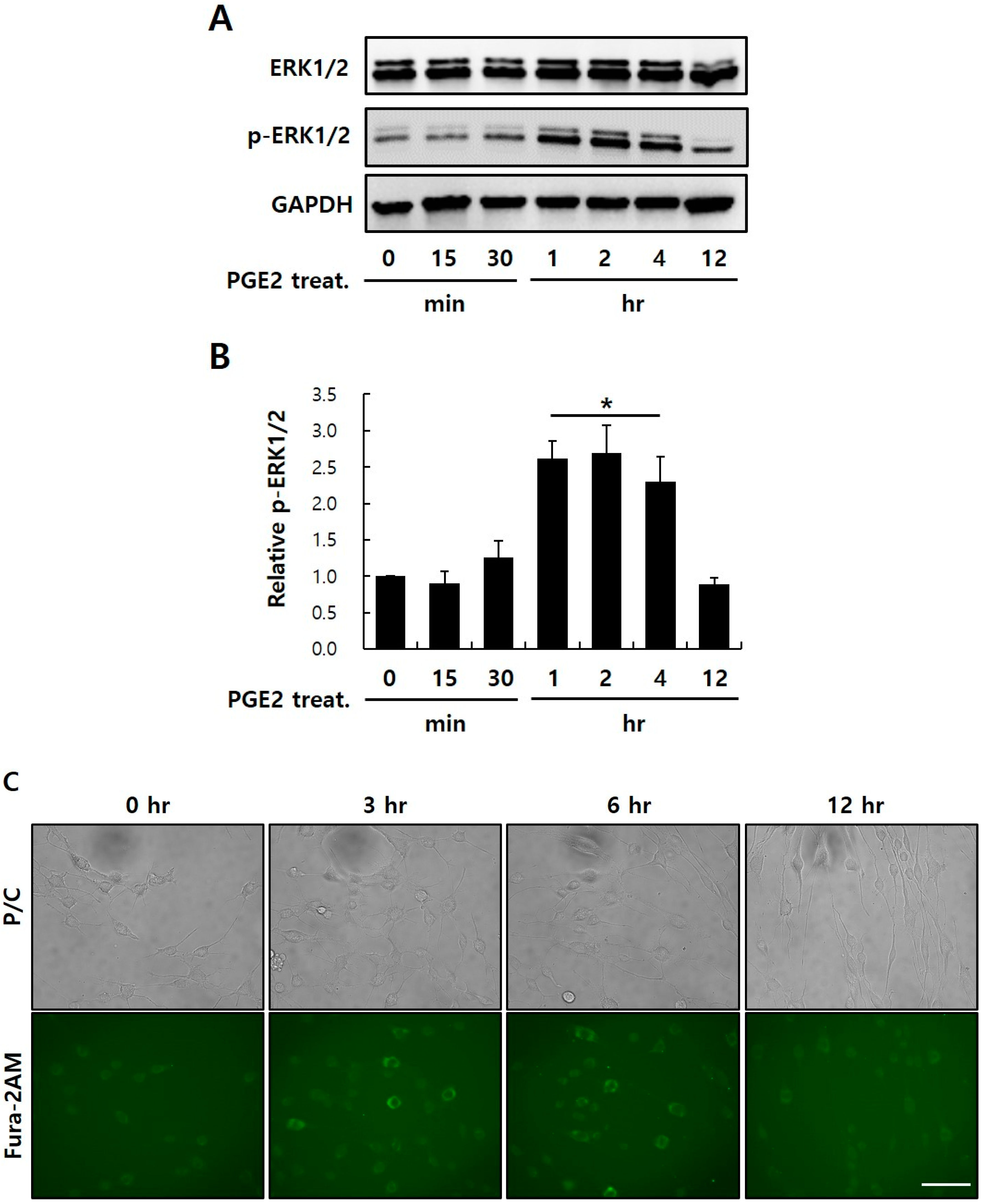
2.5. PGE2-Mediated Intracellular Signaling of EP1
3. Materials and Methods
3.1. Cell Culture of NHDFs
3.2. Effects of PGE2 on COL1A1 and MMP1 mRNA Expressions in NHDFs
3.3. In Vitro Cell Viability Assay
3.4. Real-Time qRT-PCR
3.5. EP1 siRNA Transfection
3.6. Immunoblotting Analysis
3.7. Ca2+ Imaging
3.8. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Fisher, G.J.; Varani, J.; Voorhees, J.J. Looking older: Fibroblast collapse and therapeutic implications. Arch. Dermatol. 2008, 144, 666–672. [Google Scholar] [CrossRef]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased collagen production in chronologically aged skin: Roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Verzijl, N.; DeGroot, J.; Thorpe, S.R.; Bank, R.A.; Shaw, J.N.; Lyons, T.J.; Bijlsma, J.W.; Lafeber, F.P.; Baynes, J.W.; TeKoppele, J.M. Effect of collagen turnover on the accumulation of advanced glycation end products. J. Biol. Chem. 2000, 275, 39027–39031. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Quan, T.; Purohit, T.; Shao, Y.; Cho, M.K.; He, T.; Varani, J.; Kang, S.; Voorhees, J.J. Collagen fragmentation promotes oxidative stress and elevates matrix metalloproteinase-1 in fibroblasts in aged human skin. Am. J. Pathol. 2009, 174, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, L.; Cho, H.; Xie, X.; Eliceiri, G.L. Additional MDA-MB-231 breast cancer cell matrix metalloproteinases promote invasiveness. J. Cell Physiol. 2008, 216, 480–485. [Google Scholar] [CrossRef]
- Mammoto, T.; Mammoto, A.; Ingber, D.E. Mechanobiology and developmental control. Annu. Rev. Cell Dev. Biol. 2013, 29, 27–61. [Google Scholar] [CrossRef]
- Tarutani, M.; Cai, T.; Dajee, M.; Khavari, P.A. Inducible activation of Ras and Raf in adult epidermis. Cancer Res. 2003, 63, 319–923. [Google Scholar]
- Xia, W.; Hammerberg, C.; Li, Y.; He, T.; Quan, T.; Voorhees, J.J.; Fisher, G.J. Expression of catalytically active matrix metalloproteinase-1 in dermal fibroblasts induces collagen fragmentation and functional alterations that resemble aged human skin. Aging Cell 2013, 12, 661–671. [Google Scholar] [CrossRef]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef]
- Li, Y.; Lei, D.; Swindell, W.R.; Xia, W.; Weng, S.; Fu, J.; Worthen, C.A.; Okubo, T.; Johnston, A.; Gudjonsson, J.E.; et al. Age-associated increase in skin fibroblast-derived prostaglandin E2 contributes to reduced collagen levels in elderly human skin. J. Investig. Dermatol. 2015, 135, 2181–2188. [Google Scholar] [CrossRef]
- Liu, X.; Wu, H.; Byrne, M.; Jeffrey, J.; Krane, S.; Jaenisch, R. A targeted mutation at the known collagenase cleavage site in mouse type I collagen impairs tissue remodeling. J. Cell Biol. 1995, 130, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Iskratsch, T.; Wolfenson, H.; Sheetz, M.P. Appreciating force and shape-the rise of mechanotransduction in cell biology. Nat. Rev. Mol. Cell Biol. 2014, 15, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Pickup, M.; Novitskiy, S.; Moses, H.L. The roles of TGFbeta in the tumour microenvironment. Nat. Rev. Cancer 2013, 13, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; Fisher, G. Role of age-associated alterations of the dermal extracellular matrix microenvironment in human skin aging. Gerontology. 2015, 61, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Verrecchia, F.; Chu, M.L.; Mauviel, A. Identification of novel TGF-beta/Smad gene targets in dermal fibroblasts using a combined cDNA microarray/promoter transactivation approach. J. Biol. Chem. 2001, 276, 17058–17062. [Google Scholar] [CrossRef] [PubMed]
- Muller-Decker, K. Cyclooxygenase-dependent signaling is causally linked to non-melanoma skin carcinogenesis: Pharmacological, genetic, and clinical evidence. Cancer Metastasis Rev. 2011, 30, 343–361. [Google Scholar] [CrossRef] [PubMed]
- Rundhaug, J.E.; Simper, M.S.; Surh, I.; Fischer, S.M. The role of the EP receptors for prostaglandin E2 in skin and skin cancer. Cancer Metastasis Rev. 2011, 30, 465–480. [Google Scholar] [CrossRef]
- Majumder, M.; Nandi, P.; Omar, A.; Ugwuagbo, K.C.; Lala, P.K. EP4 as a therapeutic target for aggressive human breast cancer. Int. J. Mol. Sci. 2018, 19, E1019. [Google Scholar] [CrossRef]
- Samuelsson, B.; Morgenstern, R.; Jakobsson, P.J. Membrane prostaglandin E synthase-1: A novel therapeutic target. Pharm. Rev. 2007, 59, 207–224. [Google Scholar] [CrossRef]
- Ho, A.T.V.; Palla, A.R.; Blake, M.R.; Yucel, N.D.; Wang, Y.X.; Magnusson, K.E.G.; Holbrook, C.A.; Kraft, P.E.; Delp, S.L.; Blau, H.M. Prostaglandin E2 is essential for efficacious skeletal muscle stem-cell function, augmenting regeneration and strength. Proc. Natl. Acad. Sci. USA 2017, 27, 6675–6684. [Google Scholar] [CrossRef]
- Qin, Z.; Voorhees, J.J.; Fisher, G.J.; Quan, T. Age-associated reduction of cellular spreading/mechanical force up-regulates matrix metalloproteinase-1 expression and collagen fibril fragmentation via c-Jun/AP-1 in human dermal fibroblasts. Aging Cell. 2014, 13, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.A.; Smith, W.L.; Narumiya, S. International Union of Pharmacology classification of prostanoid receptors: Properties, distribution, and structure of the receptors and their subtypes. Pharm. Rev. 1994, 46, 205–229. [Google Scholar] [PubMed]
- Negishi, M.; Sugimoto, Y.; Ichikawa, A. Molecular mechanisms of diverse actions of prostanoid receptors. Biochim. Biophys. Acta 1995, 26, 109–119. [Google Scholar] [CrossRef]
- Narumiya, S. Prostanoid receptors and signal transduction. Prog. Brain Res. 1996, 113, 231–241. [Google Scholar]
- Choung, J.; Taylor, L.; Thomas, K.; Zhou, X.; Kagan, H.; Yang, X.; Polgar, P. Role of EP2 receptors and cAMP in prostaglandin E2 regulated expression of type I collagen alpha1, lysyl oxidase, and cyclooxygenase-1 genes in human embryo lung fibroblasts. J. Cell Biochem. 1998, 71, 254–263. [Google Scholar] [CrossRef]
- Kolodsick, J.E.; Peters-Golden, M.; Larios, J.; Toews, G.B.; Thannickal, V.J.; Moore, B.B. Prostaglandin E2 inhibits fibroblast to myofibroblast transition via E. prostanoid receptor 2 signaling and cyclic adenosine monophosphate elevation. Am. J. Respir. Cell Mol. Biol. 2003, 29, 537–544. [Google Scholar] [CrossRef]
- Hayashi, T.; Nishihira, J.; Koyama, Y.; Sasaki, S.; Yamamoto, Y. Decreased prostaglandin E2 production by inflammatory cytokine and lower expression of EP2 receptor result in increased collagen synthesis in keloid fibroblasts. J. Investig. Dermatol. 2006, 26, 990–997. [Google Scholar] [CrossRef]
- Zhao, J.; Shu, B.; Chen, L.; Tang, J.; Zhang, L.; Xie, J.; Liu, X.; Xu, Y.; Qi, S. Prostaglandin E2 inhibits collagen synthesis in dermal fibroblasts and prevents hypertrophic scar formation in vivo. Exp. Dermatol. 2016, 25, 604–610. [Google Scholar] [CrossRef]
- Saad, M.J.; Folli, F.; Araki, E.; Hashimoto, N.; Csermely, P.; Kahn, C.R. Regulation of insulin receptor, insulin receptor substrate-1 and phosphatidylinositol 3-kinase in 3T3-F442A adipocytes. Effects of differentiation, insulin, and dexamethasone. Mol. Endocrinol. 1994, 8, 545–557. [Google Scholar]
- Korn, J.H. Fibroblast prostaglandin E2 synthesis. Persistence of an abnormal phenotype after short-term exposure to mononuclear cell products. J. Clin. Investig. 1983, 71, 1240–1246. [Google Scholar] [CrossRef]
- Yeh, F.L.; Shen, H.D.; Lin, M.W.; Chang, C.Y.; Tai, H.Y.; Huang, M.H. Keloid-derived fibroblasts have a diminished capacity to produce prostaglandin E2. Burns 2006, 32, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Kimelberg, H.K.; Cai, Z.; Rastogi, P.; Charniga, C.J.; Goderie, S.; Dave, V.; Jalonen, T.O. Transmitter-induced calcium responses differ in astrocytes acutely isolated from rat brain and in culture. J. Neurochem. 1997, 68, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Toivari, E.; Manninen, T.; Nahata, A.K.; Jalonen, T.O.; Linne, M.L. Effects of transmitters and amyloid-beta peptide on calcium signals in rat cortical astrocytes: Fura-2AM measurements and stochastic model simulations. PLoS ONE 2011, 29, e17914. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Chou, C.C.; Li, R.; Liu, J.; Zhang, L.; Zhu, W.; Hu, J.; Yang, B.; Tian, J. Chalcomoracin is a potent anticancer agent acting through triggering Oxidative stress via a mitophagy- and paraptosis-dependent mechanism. Sci. Rep. 2018, 22, 9566. [Google Scholar] [CrossRef]
- Miki, T.; Matsunami, M.; Nakamura, S.; Okada, H.; Matsuya, H.; Kawabata, A. ONO-8130, a selective prostanoid EP1 receptor antagonist, relieves bladder pain in mice with cyclophosphamide-induced cystitis. Pain 2011, 152, 1373–1381. [Google Scholar] [CrossRef]
- Kondo, T.; Sei, H.; Yamasaki, T.; Tomita, T.; Ohda, Y.; Oshima, T.; Fukui, H.; Watari, J.; Miwa, H. A novel prostanoid EP1 receptor antagonist, ONO-8539, reduces acid-induced heartburn symptoms in healthy male volunteers: A randomized clinical trial. J. Gastroenterol. 2017, 52, 1081–1089. [Google Scholar] [CrossRef]
- Ostenfeld, T.; Beaumont, C.; Bullman, J.; Beaumont, M.; Jeffrey, P. Human microdose evaluation of the novel EP1 receptor antagonist GSK269984A. Br. J. Clin. Pharmacol. 2012, 74, 1033–1044. [Google Scholar] [CrossRef]
- Hall, A.; Brown, S.H.; Budd, C.; Clayton, N.M.; Giblin, G.M.; Goldsmith, P.; Hayhow, T.G.; Hurst, D.N.; Naylor, A.; Anthony, R.D.; et al. Discovery of GSK345931A: An EP(1) receptor antagonist with efficacy in preclinical models of inflammatory pain. Bioorg. Med. Chem. Lett. 2009, 15, 497–501. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene Name | Assay ID |
|---|---|---|
| COL1A1 | Collagen, type I, alpha 1 | Hs00164004_m1 |
| MMP1 | Matrix metallopeptidase 1 (interstitial collagenase) | Hs00899658_m1 |
| EP1 | Prostaglandin E receptor 1 | Hs00168752_m1 |
| EP2 | Prostaglandin E receptor 2 | Hs00168754_m1 |
| EP3 | Prostaglandin E receptor 3 | Hs00168755_m1 |
| EP4 | Prostaglandin E receptor 4 | Hs00168761_m1 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | 43333764F |
| Symbol | ID Number |
|---|---|
| EP1 siRNA | S194725 |
| EP2 siRNA | S11448 |
| EP3 siRNA | S11451 |
| EP4 siRNA | S11454 |
| Control siRNA | D-001810-01-05 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shim, J.H. Prostaglandin E2 Induces Skin Aging via E-Prostanoid 1 in Normal Human Dermal Fibroblasts. Int. J. Mol. Sci. 2019, 20, 5555. https://doi.org/10.3390/ijms20225555
Shim JH. Prostaglandin E2 Induces Skin Aging via E-Prostanoid 1 in Normal Human Dermal Fibroblasts. International Journal of Molecular Sciences. 2019; 20(22):5555. https://doi.org/10.3390/ijms20225555
Chicago/Turabian StyleShim, Joong Hyun. 2019. "Prostaglandin E2 Induces Skin Aging via E-Prostanoid 1 in Normal Human Dermal Fibroblasts" International Journal of Molecular Sciences 20, no. 22: 5555. https://doi.org/10.3390/ijms20225555
APA StyleShim, J. H. (2019). Prostaglandin E2 Induces Skin Aging via E-Prostanoid 1 in Normal Human Dermal Fibroblasts. International Journal of Molecular Sciences, 20(22), 5555. https://doi.org/10.3390/ijms20225555