Broad-Spectrum Amino Acid Transporters ClAAP3 and ClAAP6 Expressed in Watermelon Fruits


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification of Amino Acid Importers from Watermelon
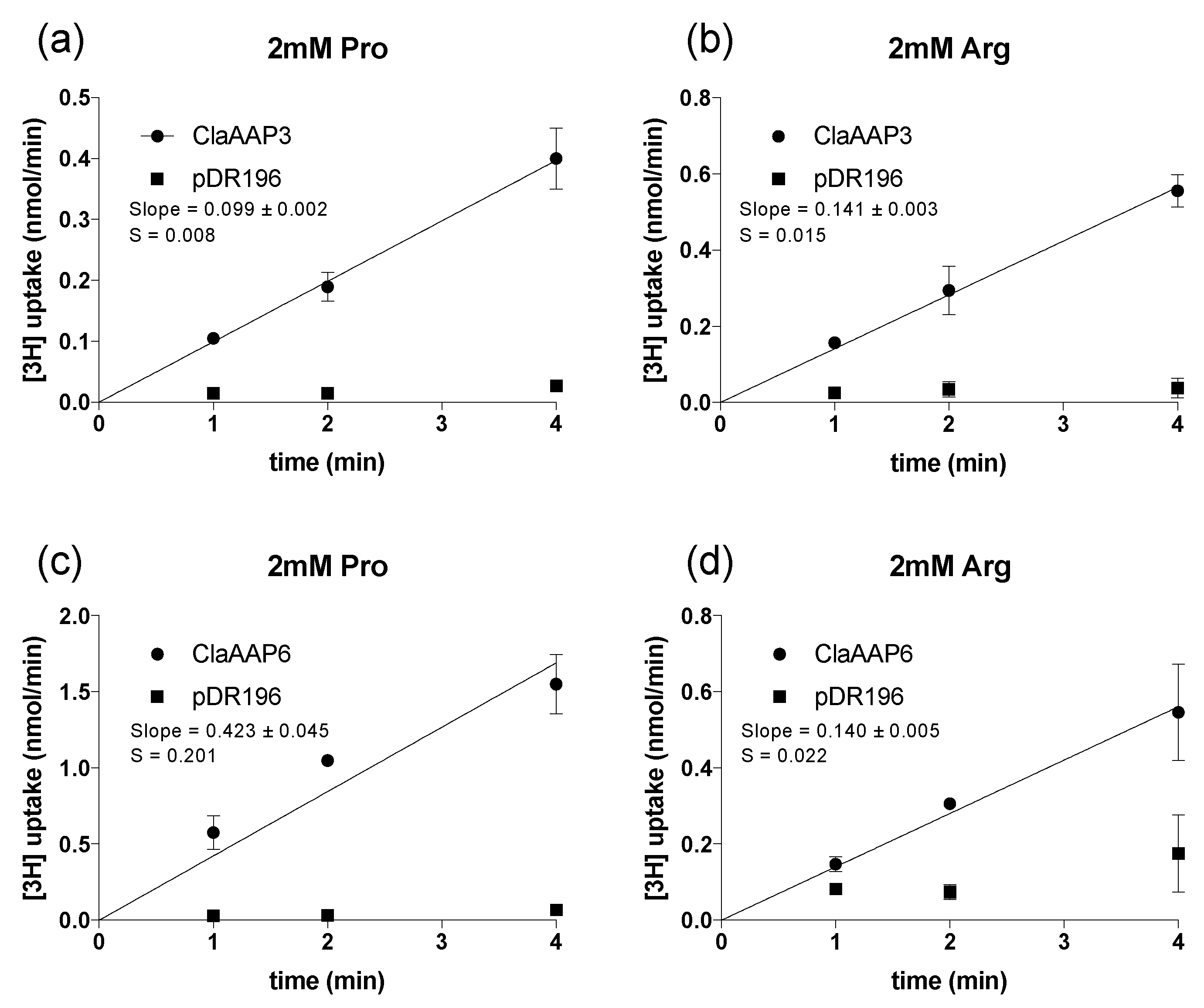
2.2. ClAAP3 and ClAAP6 Complements the Growth of Yeast Cells Deficient in Amino Acid Uptake
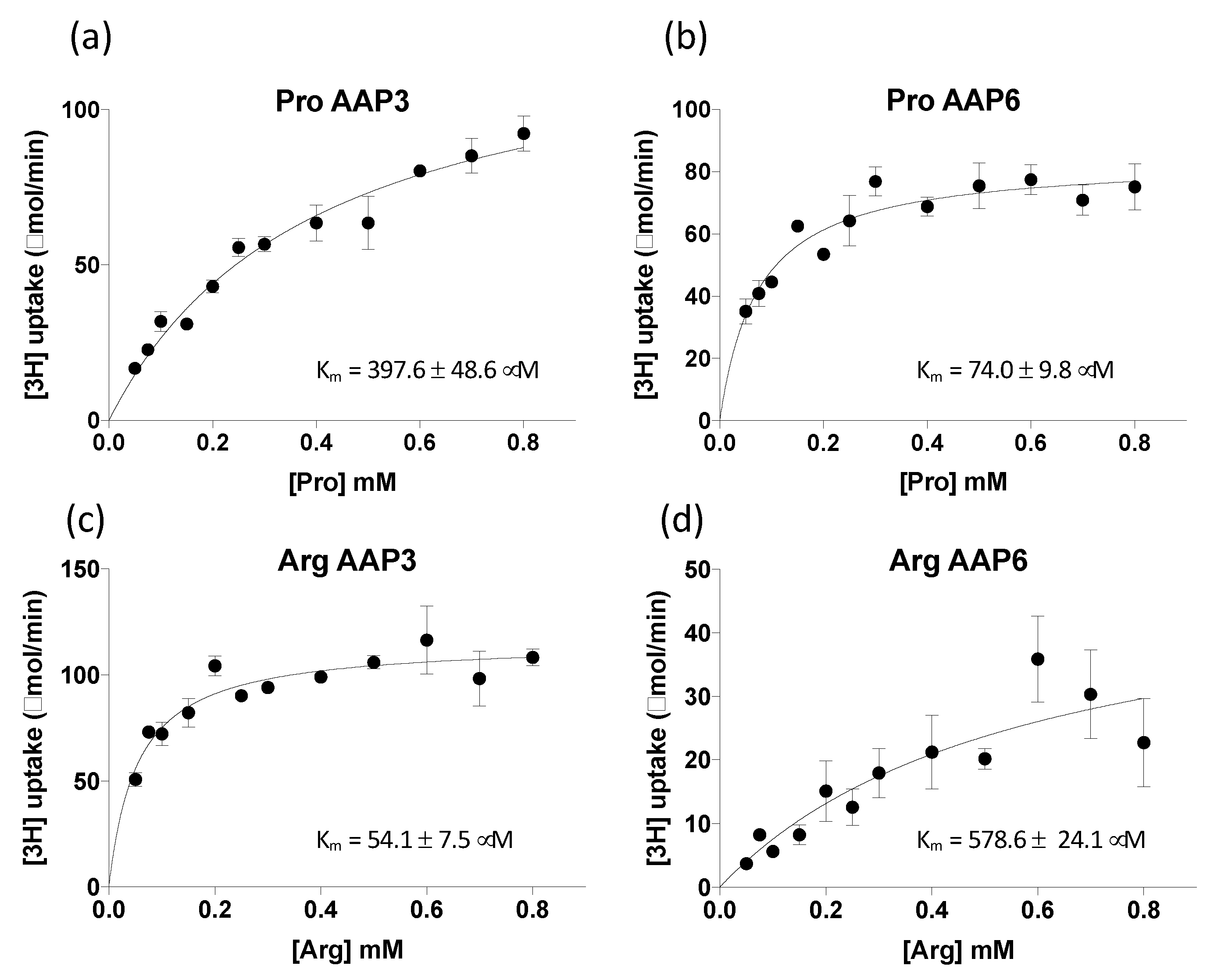
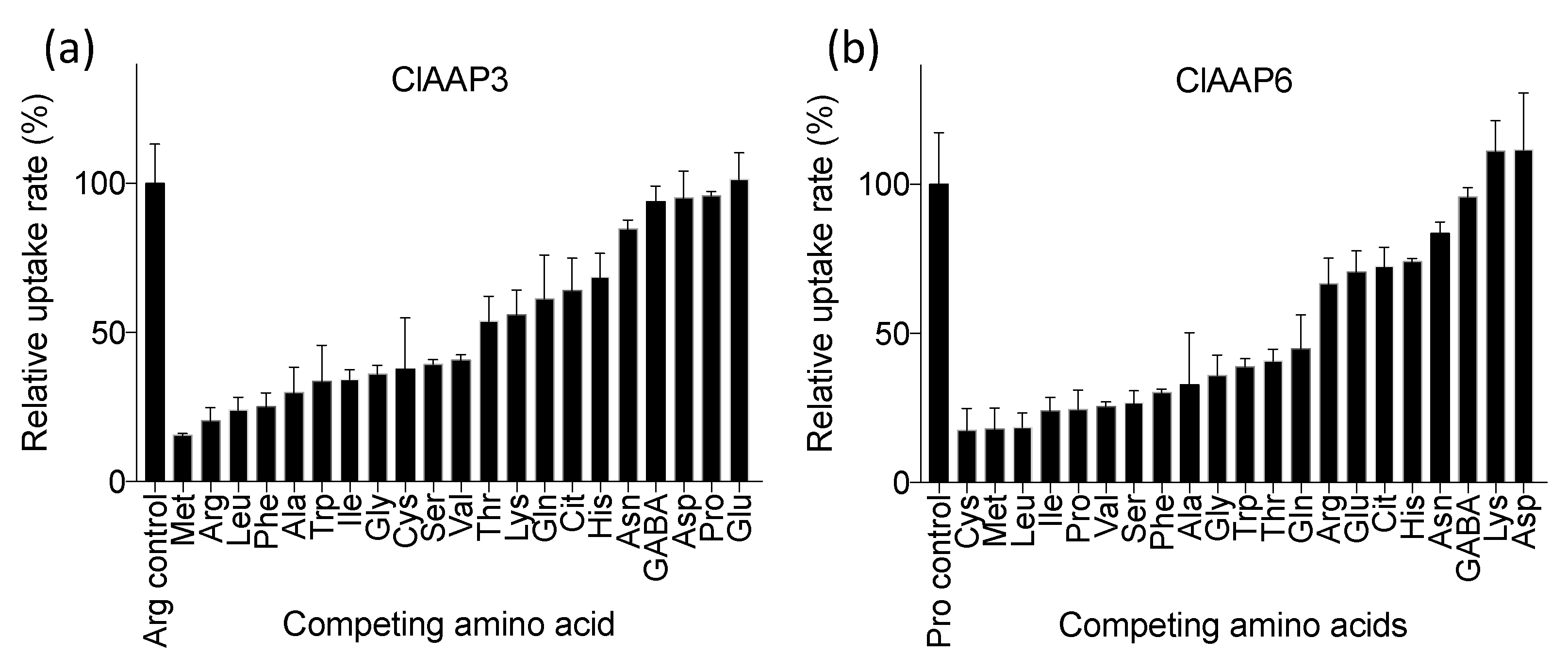
2.3. ClAAP3 and ClAAP6 are Broad-Spectrum Amino Acid Transporters
2.4. Subcellular Localization of ClaAAP3 and ClaAAP6
3. Discussion
4. Materials and Methods
4.1. Identification of Amino Acid Transporters from the Watermelon Genome
4.2. DNA Constructs
4.3. Plant Material, Growth, Transformation, and Analysis
4.4. Yeast Assays
4.5. Transient Expression of GFP Fusions in Nicotiana Benthamiana
4.6. Transient Expression of GFP Fusions in Protoplasts Isolated from Watermelon Fruits
4.7. Confocal Microscopy
4.8. Total RNA Extraction from Watermelon Tissues and qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allerton, T.D.; Proctor, D.N.; Stephens, J.M.; Dugas, T.R.; Spielmann, G.; Irving, B.A. l-Citrulline Supplementation: Impact on Cardiometabolic Health. Nutrients 2018, 10, 921. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, A.; Wong, A.; Jaime, S.J.; Gonzales, J.U. Influence of L-citrulline and watermelon supplementation on vascular function and exercise performance. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Tousoulis, D.; Kampoli, A.M.; Tentolouris, C.; Papageorgiou, N.; Stefanadis, C. The role of nitric oxide on endothelial function. Curr. Vasc. Pharm. 2012, 10, 4–18. [Google Scholar] [CrossRef]
- Bahri, S.; Zerrouk, N.; Aussel, C.; Moinard, C.; Crenn, P.; Curis, E.; Chaumeil, J.C.; Cynober, L.; Sfar, S. Citrulline: From metabolism to therapeutic use. Nutrition 2013, 29, 479–484. [Google Scholar] [CrossRef]
- Papadia, C.; Osowska, S.; Cynober, L.; Forbes, A. Citrulline in health and disease. Review on human studies. Clin. Nutr. 2018, 37, 1823–1828. [Google Scholar] [CrossRef]
- Akashi, K.; Yoshimura, K.; Nanasato, Y.; Takahara, K.; Munekage, Y.; Yokota, A. Wild plant resources for studying molecular mechanisms of drought/strong light stress tolerance. Plant. Biotechnol. 2008, 25, 257–263. [Google Scholar] [CrossRef]
- Joshi, V.; Fernie, A.R. Citrulline metabolism in plants. Amino Acids 2017, 49, 1543–1559. [Google Scholar] [CrossRef]
- Guo, S.G.; Zhang, J.G.; Sun, H.H.; Salse, J.; Lucas, W.J.; Zhang, H.Y.; Zheng, Y.; Mao, L.Y.; Ren, Y.; Wang, Z.W.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51. [Google Scholar] [CrossRef]
- Guo, S.; Sun, H.; Zhang, H.; Liu, J.; Ren, Y.; Gong, G.; Jiao, C.; Zheng, Y.; Yang, W.; Fei, Z.; et al. Comparative Transcriptome Analysis of Cultivated and Wild Watermelon during Fruit Development. PLoS ONE 2015, 10, e0130267. [Google Scholar] [CrossRef]
- Joshi, V.; Joshi, M.; Silwal, D.; Noonan, K.; Rodriguez, S.; Penalosa, A. Systematized biosynthesis and catabolism regulate citrulline accumulation in watermelon. Phytochemistry 2019, 162, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Kasting, R.; Delwiche, C.C. Ornithine, Citrulline, and Arginine Metabolism in Watermelon Seedlings. Plant. Physiol. 1958, 33, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.E.; Madore, M.A. Patterns of Assimilate Production and Translocation in Muskmelon (Cucumis-Melo L.): 2. Low-Temperature Effects. Plant. Physiol. 1992, 99, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Lohaus, G.; Moellers, C. Phloem transport of amino acids in two Brassica napus L. genotypes and one B-carinata genotype in relation to their seed protein content. Planta 2000, 211, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.; Collier, R.; Trethewy, A.; Gould, G.; Sieker, R.; Tegeder, M. AAP1 regulates import of amino acids into developing Arabidopsis embryos. Plant. J. 2009, 59, 540–552. [Google Scholar] [CrossRef]
- Zhang, L.; Garneau, M.G.; Majumdar, R.; Grant, J.; Tegeder, M. Improvement of pea biomass and seed productivity by simultaneous increase of phloem and embryo loading with amino acids. Plant. J. 2015, 81, 134–146. [Google Scholar] [CrossRef]
- Zhu, Q.; Gao, P.; Liu, S.; Zhu, Z.; Amanullah, S.; Davis, A.R.; Luan, F. Comparative transcriptome analysis of two contrasting watermelon genotypes during fruit development and ripening. BMC Genom. 2017, 18, 3. [Google Scholar] [CrossRef]
- Fischer, W.N.; Kwart, M.; Hummel, S.; Frommer, W.B. Substrate-Specificity and Expression Profile of Amino-Acid Transporters (Aaps) in Arabidopsis. J. Biol. Chem. 1995, 270, 16315–16320. [Google Scholar] [CrossRef]
- Fischer, W.N.; Loo, D.D.; Koch, W.; Ludewig, U.; Boorer, K.J.; Tegeder, M.; Rentsch, D.; Wright, E.M.; Frommer, W.B. Low and high affinity amino acid H+-cotransporters for cellular import of neutral and charged amino acids. Plant. J. 2002, 29, 717–731. [Google Scholar] [CrossRef]
- Neelam, A.; Marvier, A.C.; Hall, J.L.; Williams, L.E. Functional characterization and expression analysis of the amino acid permease RcAAP3 from castor bean. Plant. Physiol. 1999, 120, 1049–1056. [Google Scholar] [CrossRef]
- Taylor, M.R.; Reinders, A.; Ward, J.M. Transport Function of Rice Amino Acid Permeases (AAPs). Plant. Cell Physiol. 2015, 56, 1355–1363. [Google Scholar] [CrossRef]
- Besnard, J.; Pratelli, R.; Zhao, C.; Sonawala, U.; Collakova, E.; Pilot, G.; Okumoto, S. UMAMIT14 is an amino acid exporter involved in phloem unloading in Arabidopsis roots. J. Exp. Bot. 2016, 67, 6385–6397. [Google Scholar] [CrossRef] [PubMed]
- Fish, W.W. The Expression of Citrulline and other Members of the Arginine Metabolic Family in Developing Watermelon Fruit Int. J. Agric. Res. Innov. Technol. 2014, 2, 665–672. [Google Scholar]
- DeBusk, R.M.; Ogilvie, S. Participation of an extracellular deaminase in amino acid utilization by Neurospora crassa. J. Bacteriol. 1984, 159, 583–589. [Google Scholar] [PubMed]
- DeBusk, R.M.; Ogilvie-Villa, S. Physiological adaptation to the loss of amino acid transport ability. J. Bacteriol. 1982, 152, 545–548. [Google Scholar] [PubMed]
- Su, Y.H.; Frommer, W.B.; Ludewig, U. Molecular and functional characterization of a family of amino acid transporters from Arabidopsis. Plant. Physiol. 2004, 136, 3104–3113. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Yu, C.K.; Qu, S.; San, M.W.; Li, K.Y.; Lo, S.W.; Jiang, L. The Golgi-localized Arabidopsis endomembrane protein12 contains both endoplasmic reticulum export and Golgi retention signals at its C terminus. Plant. Cell 2012, 24, 2086–2104. [Google Scholar] [CrossRef]
- Kasaras, A.; Kunze, R. Dual-targeting of Arabidopsis DMP1 isoforms to the tonoplast and the plasma membrane. PLoS ONE 2017, 12, e0174062. [Google Scholar] [CrossRef]
- Nelson, B.K.; Cai, X.; Nebenführ, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant. J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef]
- Akashi, K.; Mifune, Y.; Morita, K.; Ishitsuka, S.; Tsujimoto, H.; Ishihara, T. Spatial accumulation pattern of citrulline and other nutrients in immature and mature watermelon fruits. J. Sci. Food Agr. 2017, 97, 479–487. [Google Scholar] [CrossRef]
- Okumoto, S.; Koch, W.; Tegeder, M.; Fischer, W.N.; Biehl, A.; Leister, D.; Stierhof, Y.D.; Frommer, W.B. Root phloem-specific expression of the plasma membrane amino acid proton co-transporter AAP3. J. Exp. Bot. 2004, 55, 2155–2168. [Google Scholar] [CrossRef]
- Peng, B.; Kong, H.; Li, Y.; Wang, L.; Zhong, M.; Sun, L.; Gao, G.; Zhang, Q.; Luo, L.; Wang, G.; et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 2014, 5, 4847. [Google Scholar] [CrossRef] [PubMed]
- Palacin, M.; Kanai, Y. The ancillary proteins of HATs: SLC3 family of amino acid transporters. Pflug. Arch. 2004, 447, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Santiago, P.; Lagunas-Gomez, D.; Barkla, B.J.; Vera-Estrella, R.; Lalonde, S.; Jones, A.; Frommer, W.B.; Zimmermannova, O.; Sychrova, H.; Pantoja, O. Identification of rice cornichon as a possible cargo receptor for the Golgi-localized sodium transporter OsHKT1;3. J. Exp. Bot. 2015, 66, 2733–2748. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Santiago, P.; Lagunas-Gomez, D.; Yanez-Dominguez, C.; Vera-Estrella, R.; Zimmermannova, O.; Sychrova, H.; Pantoja, O. Plant and yeast cornichon possess a conserved acidic motif required for correct targeting of plasma membrane cargos. Biochim. Biophys. Acta 2017, 1864, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, A.M.; Liu, Y.; Vidensky, S.; Maier, S.; Jung, E.; Farhan, H.; Robinson, M.B.; Sitte, H.H.; Rothstein, J.D. The endoplasmic reticulum exit of glutamate transporter is regulated by the inducible mammalian Yip6b/GTRAP3-18 protein. J. Biol. Chem. 2008, 283, 6175–6183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besnard, J.; Zhao, C.; Avice, J.C.; Vitha, S.; Hyodo, A.; Pilot, G.; Okumoto, S. Arabidopsis UMAMIT24 and 25 are amino acid exporters involved in seed loading. J. Exp. Bot. 2018, 69, 5221–5232. [Google Scholar] [CrossRef]
- Yang, H.; Krebs, M.; Stierhof, Y.D.; Ludewig, U. Characterization of the putative amino acid transporter genes AtCAT2, 3 &4: The tonoplast localized AtCAT2 regulates soluble leaf amino acids. J. Plant. Physiol. 2014, 171, 594–601. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.S.; Eddy, S.R.; Portugaly, E. Hidden Markov model speed heuristic and iterative HMM search procedure. BMC Bioinform. 2010, 11, 431. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalonde, S.; Sero, A.; Pratelli, R.; Pilot, G.; Chen, J.; Sardi, M.I.; Parsa, S.A.; Kim, D.Y.; Acharya, B.R.; Stein, E.V.; et al. A membrane protein/signaling protein interaction network for Arabidopsis version AMPv2. Front. Physiol. 2010, 1, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loque, D.; Lalonde, S.; Looger, L.L.; von Wiren, N.; Frommer, W.B. A cytosolic trans-activation domain essential for ammonium uptake. Nature 2007, 446, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulos, A.; Sutikovic, Z.; Wenzl, C.; Maegele, I.; Lohmann, J.U.; Forner, J. GreenGate---a novel, versatile, and efficient cloning system for plant transgenesis. PLoS ONE 2013, 8, e83043. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.; Suzuki, T.; Murata, S.; Nakamura, S.; Hino, T.; Maeo, K.; Tabata, R.; Kawai, T.; Tanaka, K.; Niwa, Y.; et al. Improved Gateway binary vectors: High-performance vectors for creation of fusion constructs in transgenic analysis of plants. Biosci. Biotechnol. Biochem. 2007, 71, 2095–2100. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Eskandari, S.; Grallath, S.; Rentsch, D. AtGAT1, a high affinity transporter for gamma-aminobutyric acid in Arabidopsis thaliana. J. Biol. Chem. 2006, 281, 7197–7204. [Google Scholar] [CrossRef] [Green Version]
- Wydro, M.; Kozubek, E.; Lehmann, P. Optimization of transient Agrobacterium-mediated gene expression system in leaves of Nicotiana benthamiana. Acta Biochim. Pol. 2006, 53, 289–298. [Google Scholar]
- Weigel, D.; Glazebrook, J. Arabidopsis: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2002. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LHT (8) | ProT (1) | GAT (4) | AAP (8) | ATLa (5) | ATLb (8) | LAX/AUX (7) | PUT (5) | CAT (8) |
|---|---|---|---|---|---|---|---|---|
| Cla006527 | Cla004309 | Cla020511 | Cla005590 | Cla010799 | Cla000539 | Cla015837 | Cla005022 | Cla003079 |
| Cla004045 | Cla020512 | Cla023090 | Cla009967 | Cla015743 | Cla020298 | Cla001768 | Cla003075 | |
| Cla009988 | Cla011382 | Cla013912 | Cla015343 | Cla012146 | Cla000681 | Cla007330 | Cla006711 | |
| Cla019489 | Cla010921 | Cla023187 | Cla007263 | Cla017006 | Cla017975 | Cla016085 | Cla013686 | |
| Cla020372 | Cla011628 | Cla007265 | Cla002877 | Cla018110 | Cla002221 | Cla019526 | ||
| Cla011987 | Cla011631 | Cla002878 | Cla006581 | Cla012403 | ||||
| Cla020709 | Cla011627 | Cla020546 | Cla004339 | Cla005396 | ||||
| Cla015783 | Cla011629 | Cla013755 | Cla002657 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, T.; Joshi, V.; Joshi, M.; Vitha, S.; Gibbs, H.; Wang, K.; Okumoto, S. Broad-Spectrum Amino Acid Transporters ClAAP3 and ClAAP6 Expressed in Watermelon Fruits. Int. J. Mol. Sci. 2019, 20, 5855. https://doi.org/10.3390/ijms20235855
Shi T, Joshi V, Joshi M, Vitha S, Gibbs H, Wang K, Okumoto S. Broad-Spectrum Amino Acid Transporters ClAAP3 and ClAAP6 Expressed in Watermelon Fruits. International Journal of Molecular Sciences. 2019; 20(23):5855. https://doi.org/10.3390/ijms20235855
Chicago/Turabian StyleShi, Tianran, Vijay Joshi, Madhumita Joshi, Stanislav Vitha, Holly Gibbs, Kehua Wang, and Sakiko Okumoto. 2019. "Broad-Spectrum Amino Acid Transporters ClAAP3 and ClAAP6 Expressed in Watermelon Fruits" International Journal of Molecular Sciences 20, no. 23: 5855. https://doi.org/10.3390/ijms20235855