Novel Zebrafish Mono-α2,8-sialyltransferase (ST8Sia VIII): An Evolutionary Perspective of α2,8-Sialylation

 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. In Silico Identification and Sequence Analysis of Zebrafish ST8Sia Sequence
2.2. Spatio-Temporal Expression of the st8sia-Like Gene during Zebrafish Development
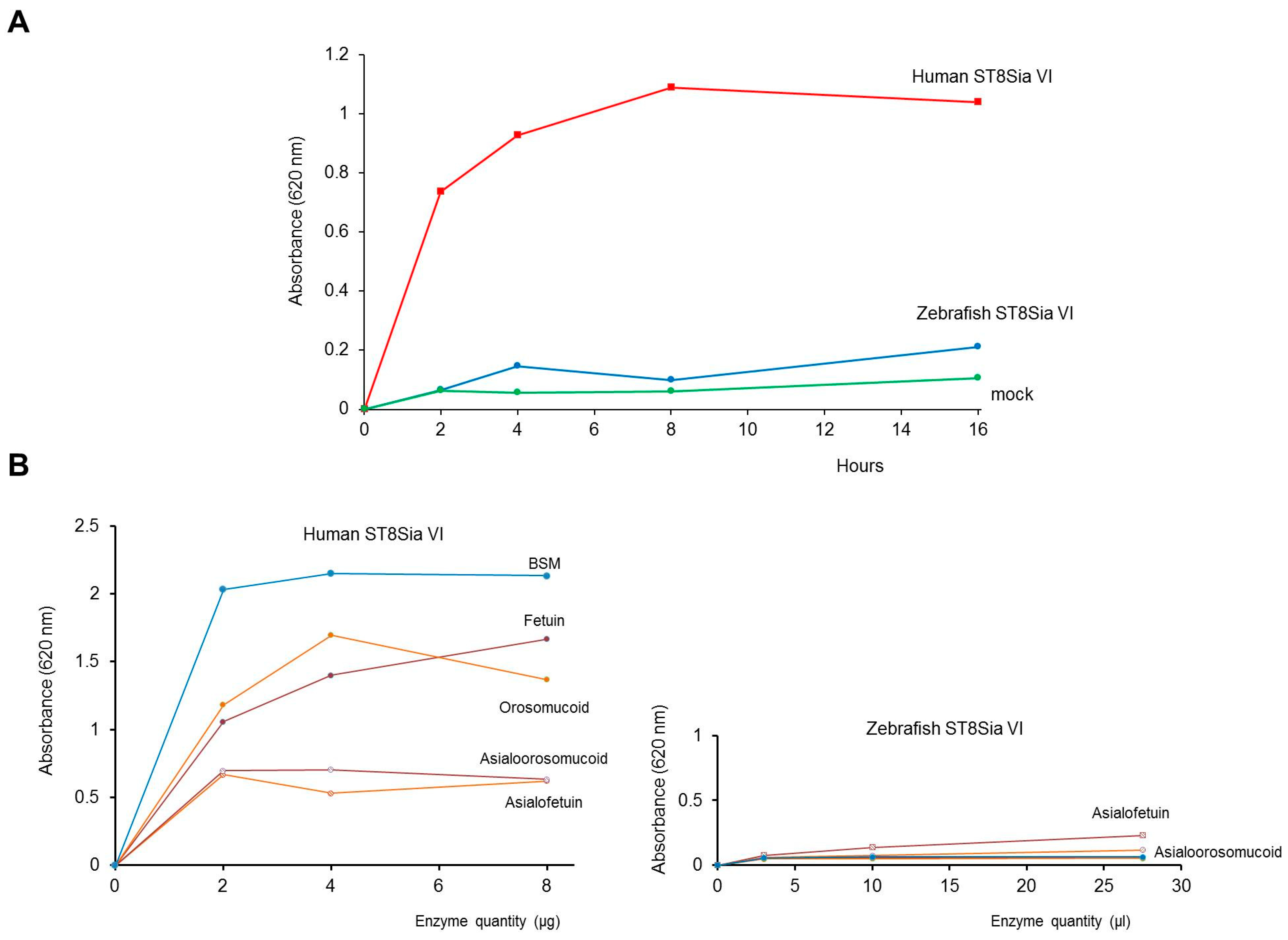
2.3. Expression of a Recombinant and Soluble Protein—Enzymatic Characterization
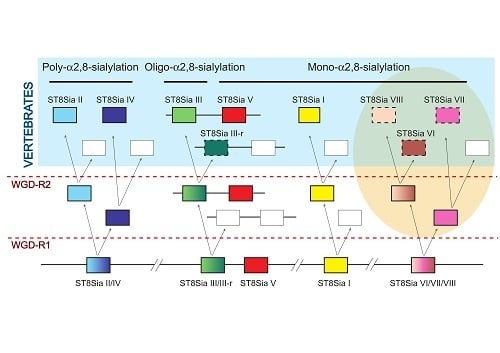
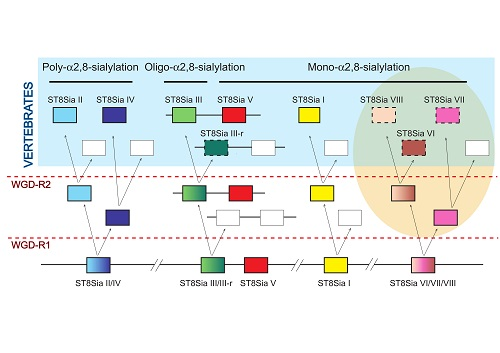
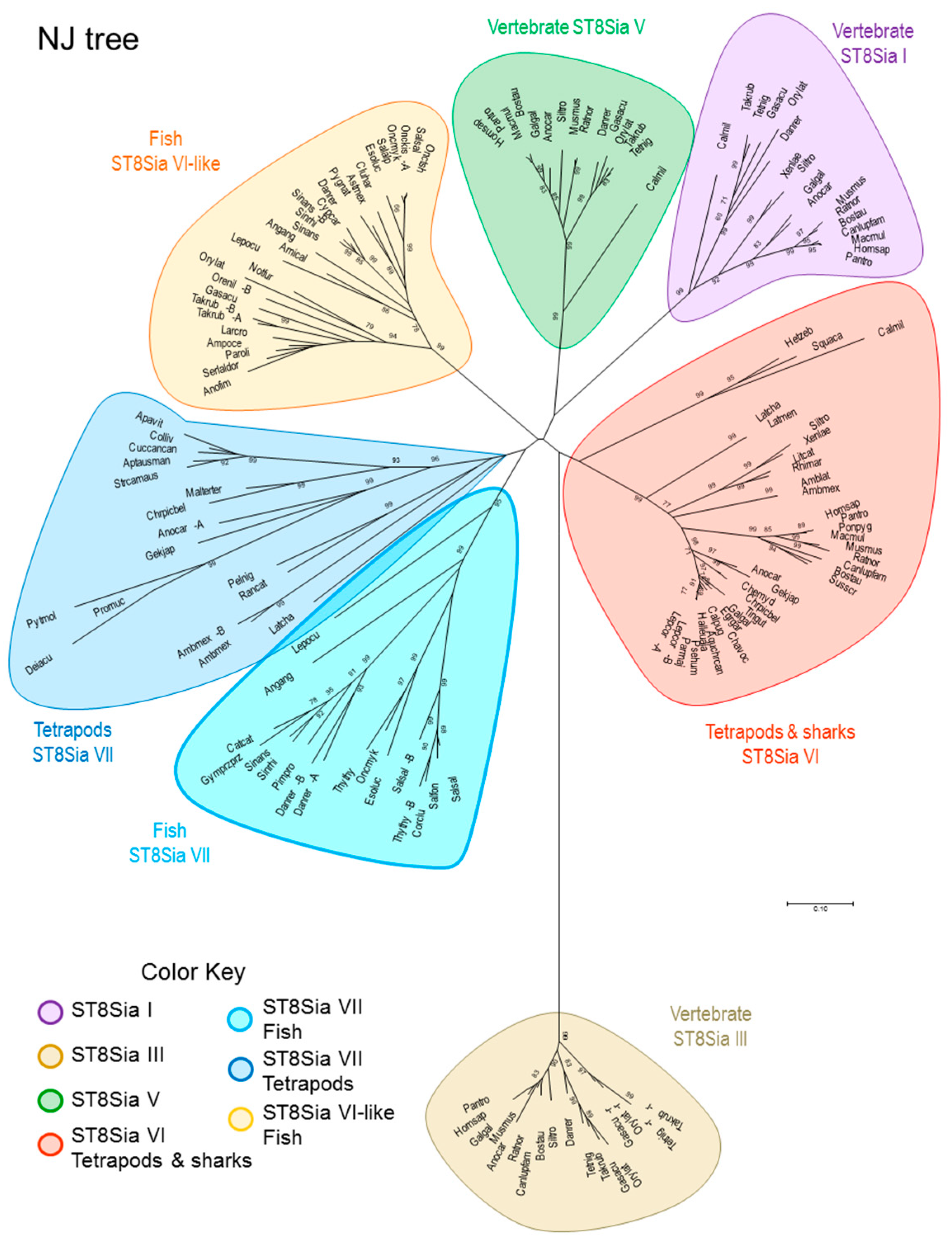
2.4. Molecular Phylogenetic and Phylogenomics Analyses Underscore Loss of the Teleost Fish st8sia6 Locus
3. Materials and Methods
3.1. Materials
3.2. In Silico Identification and Analysis of ST8Sia Sequences in Databases
3.3. Phylogenetic Analysis and Sequence Similarity Network
3.4. Synteny Analysis, Paralogon Detection and Ancestral Genome Reconstruction
3.5. RNA Extraction, cDNA Synthesis and RT-PCR
3.6. Isolation of A Zebrafish st8sia6 cDNA and Construction of an Expression Vector
3.7. Animals, Cell Culture and Transient Expression of a Soluble form of Dre ST8Sia VIII
3.8. Synthesis of Activated CMP-Sialic Acid

3.9. Enzymatic Characterization of Sialyltransferase
3.10. Real Time PCR
3.11. In Situ Hybridization
4. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| adipoQ | adiponectin |
| BSM | bovine submaxillary mucin |
| BTTAA | 2-[4-({bis[(1-tert-butyl-1H-1,2,3-triazol-4-yl)methyl]amino}methyl)-1H-1,2,3-triazol-1-yl]acetic acid |
| CMP | cytidine monophosphate |
| CuAAC | Cu(I) catalyzed azide-alkyne cycloaddition |
| diSia | disialic acid |
| DMB | 1,2-diamino-4,5-methylenedioxybenzene |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DP | degree of polymerization |
| Dre | Danio rerio |
| FBS | fetal bovine serum |
| FCS | fetal calf serum |
| Hpf | hours post fertilization |
| ISH | in situ hybridization |
| LECA | Last Common Ancestor of Eukaryotes |
| MEM | minimal essential medium |
| MPSA | Microplate sialyltransferase assay |
| Neu5Ac | N-acetylneuraminic acid |
| Neu5Gc | N-glycolylneuraminic acid |
| oligoSia | oligosialic acid |
| OSM | ovine submaxillary mucin |
| polySia or PSA | polysialic acid |
| PSGP | salmonid egg polysialoglycoprotein |
| SiaNAl | N-(4-pentynoyl)neuraminic acid |
| ST8Sia | α2,8-sialyltransferase |
| WGD | whole genome duplication |
| Kdn | 2-keto-3-deoxynononic acid |
References
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef]
- Schauer, R. Sialic acids as regulators of molecular and cellular interactions. Curr. Opin. Struct. Biol. 2009, 19, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Angata, T.; Varki, A. Chemical diversity in the sialic acids and related alpha-keto acids: An evolutionary perspective. Chem. Rev. 2002, 102, 439–469. [Google Scholar] [CrossRef] [PubMed]
- Sato, C. Polysialic acid. In Sialobiology: Structure, Biosynthesis and Function. Sialic Acid Glycoconjugates in Health and Diseases; Tiralongo, J., Martinez-Duncker, I., Eds.; Bentham Science: Emirate of Sharjah, UAE, 2013; pp. 33–75. [Google Scholar]
- Finne, J.; Krusius, T.; Rauvala, H. Occurrence of disialosyl groups in glycoproteins. Biochem. Biophys. Res. Commun. 1977, 74, 405–410. [Google Scholar] [CrossRef]
- Sato, C.; Kitajima, K. Disialic, oligosialic and polysialic acids: Distribution, functions and related disease. J. Biochem. 2013, 154, 115–136. [Google Scholar] [CrossRef]
- Schnaar, R.L.; Gerardy-Schahn, R.; Hildebrandt, H. Sialic acids in the brain: Gangliosides and polysialic acid in nervous system development, stability, disease, and regeneration. Physiol. Rev. 2014, 94, 461–518. [Google Scholar] [CrossRef] [PubMed]
- Sato, C.; Fukuoka, H.; Ohta, K.; Matsuda, T.; Koshino, R.; Kobayashi, K.; Troy, F.A., 2nd; Kitajima, K. Frequent occurrence of pre-existing alpha 2-->8-linked disialic and oligosialic acids with chain lengths up to 7 sia residues in mammalian brain glycoproteins. Prevalence revealed by highly sensitive chemical methods and anti-di-, oligo-, and poly-sia antibodies specific for defined chain lengths. J. Biol. Chem. 2000, 275, 15422–15431. [Google Scholar]
- Kiang, W.L.; Krusius, T.; Finne, J.; Margolis, R.U.; Margolis, R.K. Glycoproteins and proteoglycans of the chromaffin granule matrix. J. Biol. Chem. 1982, 257, 1651–1659. [Google Scholar]
- Wang, S.H.; Tsai, C.M.; Lin, K.I.; Khoo, K.H. Advanced mass spectrometry and chemical analyses reveal the presence of terminal disialyl motif on mouse b-cell glycoproteins. Glycobiology 2013, 23, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Dell, A.; Fukuda, M.N. Structure of fetal lactosaminoglycan. The carbohydrate moiety of band 3 isolated from human umbilical cord erythrocytes. J. Biol. Chem. 1984, 259, 4782–4791. [Google Scholar] [PubMed]
- Canis, K.; McKinnon, T.A.; Nowak, A.; Panico, M.; Morris, H.R.; Laffan, M.; Dell, A. The plasma von willebrand factor o-glycome comprises a surprising variety of structures including ABH antigens and disialosyl motifs. J. Thromb. Haemost. 2010, 8, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Storr, S.J.; Royle, L.; Chapman, C.J.; Hamid, U.M.; Robertson, J.F.; Murray, A.; Dwek, R.A.; Rudd, P.M. The o-linked glycosylation of secretory/shed muc1 from an advanced breast cancer patient’s serum. Glycobiology 2008, 18, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Lee, Y.C.; Troy ll, F.A. Sialobiology and Other Novel Forms of Glycosylation; Gakushin Publishing Company: London, UK, 1999; 307p. [Google Scholar]
- Sato, C.; Yasukawa, Z.; Honda, N.; Matsuda, T.; Kitajima, K. Identification and adipocyte differentiation-dependent expression of the unique disialic acid residue in an adipose tissue-specific glycoprotein, adipo q. J. Biol. Chem. 2001, 276, 28849–28856. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Iwasaki, M. Isolation of a novel glycoprotein from the eggs of rainbow trout: Occurrence of disialosyl groups on all carbohydrate chains. Biochem. Biophys. Res. Commun. 1978, 83, 1018–1023. [Google Scholar] [CrossRef]
- Sato, C.; Kitajima, K.; Tazawa, I.; Inoue, Y.; Inoue, S.; Troy, F.A., 2nd. Structural diversity in the alpha 2-->8-linked polysialic acid chains in salmonid fish egg glycoproteins. Occurrence of poly(Neu5Ac), poly(Neu5Gc), poly(Neu5Ac, Neu5Gc), poly(KDN), and their partially acetylated forms. J. Biol. Chem. 1993, 268, 23675–23684. [Google Scholar]
- Guerardel, Y.; Chang, L.Y.; Maes, E.; Huang, C.J.; Khoo, K.H. Glycomic survey mapping of zebrafish identifies unique sialylation pattern. Glycobiology 2006, 16, 244–257. [Google Scholar] [CrossRef]
- Hanzawa, K.; Suzuki, N.; Natsuka, S. Structures and developmental alterations of n-glycans of zebrafish embryos. Glycobiology 2017, 27, 228–245. [Google Scholar] [CrossRef]
- Chang, L.Y.; Harduin-Lepers, A.; Kitajima, K.; Sato, C.; Huang, C.J.; Khoo, K.H.; Guerardel, Y. Developmental regulation of oligosialylation in zebrafish. Glycoconj. J. 2009, 26, 247–261. [Google Scholar] [CrossRef]
- Yamakawa, N.; Vanbeselaere, J.; Chang, L.-Y.; Yu, S.-Y.; Ducrocq, L.; Harduin-Lepers, A.; Kurata, J.; Aoki-Kinoshita, K.F.; Sato, C.; Khoo, K.-H.; et al. Systems glycomics of adult zebrafish identifies organ-specific sialylation and glycosylation patterns. Nat. Commun. 2018, 9, 4647. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (cazy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef]
- Petit, D.; Teppa, E.; Cenci, U.; Ball, S.; Harduin-Lepers, A. Reconstruction of the sialylation pathway in the ancestor of eukaryotes. Sci. Rep. 2018, 8, 2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, A.K.; Paulson, J.C. The sialyltransferase “sialylmotif” participates in binding the donor substrate CMP-NeuAc. J. Biol. Chem. 1995, 270, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.K.; Paulson, J.C. Sialylmotifs of sialyltransferases. Indian J. Biochem. Biophys. 1997, 34, 157–165. [Google Scholar] [PubMed]
- Drickamer, K. A conserved disulphide bond in sialyltransferases. Glycobiology 1993, 3, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Geremia, R.A.; Harduin-Lepers, A.; Delannoy, P. Identification of two novel conserved amino acid residues in eukaryotic sialyltransferases: Implications for their mechanism of action. Glycobiology 1997, 7, 5–7. [Google Scholar] [CrossRef]
- Jeanneau, C.; Chazalet, V.; Auge, C.; Soumpasis, D.M.; Harduin-Lepers, A.; Delannoy, P.; Imberty, A.; Breton, C. Structure-function analysis of the human sialyltransferase ST3Gal I: Role of N-glycosylation and a novel conserved sialylmotif. J. Biol. Chem. 2004, 279, 13461–13468. [Google Scholar] [CrossRef] [PubMed]
- Harduin-Lepers, A. Comprehensive analysis of sialyltransferases in vertebrate genomes. Glycobiol. Insights 2010, 2, 29–61. [Google Scholar] [CrossRef]
- Harduin-Lepers, A. Vertebrate sialyltransferases. In Sialobiology: Structure, Biosynthesis and Function. Sialic Acid Glycoconjugates in Health and Diseases; Tiralongo, J., Martinez-Duncker, I., Eds.; Bentham Science: Emirate of Sharjah, UAE, 2013; pp. 139–187. [Google Scholar]
- Harduin-Lepers, A.; Mollicone, R.; Delannoy, P.; Oriol, R. The animal sialyltransferases and sialyltransferase-related genes: A phylogenetic approach. Glycobiology 2005, 15, 805–817. [Google Scholar] [CrossRef]
- Petit, D.; Teppa, R.E.; Petit, J.M.; Harduin-Lepers, A. A practical approach to reconstruct evolutionary history of animal sialyltransferases and gain insights into the sequence-function relationships of golgi-glycosyltransferases. In Glycosyltransferases: Methods and Protocols; Brochausen, I., Ed.; Springer, Humana Press: New York, NY, USA, 2013; Volume 1022, pp. 73–97. [Google Scholar]
- Harduin-Lepers, A.; Vallejo-Ruiz, V.; Krzewinski-Recchi, M.A.; Samyn-Petit, B.; Julien, S.; Delannoy, P. The human sialyltransferase family. Biochimie 2001, 83, 727–737. [Google Scholar] [CrossRef]
- Patel, R.Y.; Balaji, P.V. Identification of linkage-specific sequence motifs in sialyltransferases. Glycobiology 2006, 16, 108–116. [Google Scholar] [CrossRef]
- Harduin-Lepers, A.; Petit, D.; Mollicone, R.; Delannoy, P.; Petit, J.M.; Oriol, R. Evolutionary history of the alpha2,8-sialyltransferase (ST8Sia) gene family: Tandem duplications in early deuterostomes explain most of the diversity found in the vertebrate ST8Sia genes. BMC Evol. Biol. 2008, 8, 258. [Google Scholar] [CrossRef] [PubMed]
- Onho, S. Gene duplication and the uniqueness of vertebrate genomes circa 1970–1999. Semin. Cell Dev. Biol. 1999, 10, 517–522. [Google Scholar]
- Petit, D.; Mir, A.M.; Petit, J.M.; Thisse, C.; Delannoy, P.; Oriol, R.; Thisse, B.; Harduin-Lepers, A. Molecular phylogeny and functional genomics of β-galactoside α2,6-sialyltransferases that explain ubiquitous expression of st6gal1 gene in amniotes. J. Biol. Chem. 2010, 285, 38399–38414. [Google Scholar] [CrossRef]
- Petit, D.; Teppa, E.; Mir, A.M.; Vicogne, D.; Thisse, C.; Thisse, B.; Filloux, C.; Harduin-Lepers, A. Integrative view of α2,3-sialyltransferases (ST3Gal) molecular and functional evolution in deuterostomes: Significance of lineage-specific losses. Mol. Biol. Evol. 2015, 32, 906–927. [Google Scholar] [CrossRef] [PubMed]
- Teppa, R.E.; Petit, D.; Plechakova, O.; Cogez, V.; Harduin-Lepers, A. Phylogenetic-derived insights into the evolution of sialylation in eukaryotes: Comprehensive analysis of vertebrate beta-galactoside α2,3/6-sialyltransferases (St3Gal and St6Gal). Int. J. Mol. Sci. 2016, 17, 1286. [Google Scholar] [CrossRef] [PubMed]
- Vanbeselaere, J.; Chang, L.Y.; Harduin-Lepers, A.; Fabre, E.; Yamakawa, N.; Slomianny, C.; Biot, C.; Khoo, K.H.; Guerardel, Y. Mapping the expressed glycome and glycosyltransferases of zebrafish liver cells as a relevant model system for glycosylation studies. J. Proteome Res. 2012, 11, 2164–2177. [Google Scholar] [CrossRef] [PubMed]
- Teintenier-Lelievre, M.; Julien, S.; Juliant, S.; Guerardel, Y.; Duonor-Cerutti, M.; Delannoy, P.; Harduin-Lepers, A. Molecular cloning and expression of a human hST8Sia VI (α2,8-sialyltransferase) responsible for the synthesis of the disia motif on O-glycosylproteins. Biochem. J. 2005, 392, 665–674. [Google Scholar] [CrossRef]
- Takashima, S.; Ishida, H.K.; Inazu, T.; Ando, T.; Ishida, H.; Kiso, M.; Tsuji, S.; Tsujimoto, M. Molecular cloning and expression of a sixth type of α2,8-sialyltransferase (ST8Sia VI) that sialylates O-glycans. J. Biol. Chem. 2002, 277, 24030–24038. [Google Scholar] [CrossRef]
- Gilormini, P.A.; Lion, C.; Noel, M.; Krzewinski-Recchi, M.A.; Harduin-Lepers, A.; Guerardel, Y.; Biot, C. Improved workflow for the efficient preparation of ready to use cmp-activated sialic acids. Glycobiology 2016, 26, 1151–1156. [Google Scholar] [CrossRef]
- Noel, M.; Gilormini, P.A.; Cogez, V.; Yamakawa, N.; Vicogne, D.; Lion, C.; Biot, C.; Guerardel, Y.; Harduin-Lepers, A. Probing the cmp-sialic acid donor specificity of two human γ-d-galactoside sialyltransferases (St3Gal I and ST6Gal I) selectively acting on O- and N-glycosylproteins. Chembiochem 2017, 18, 1251–1259. [Google Scholar] [CrossRef]
- Noel, M.; Gilormini, P.A.; Cogez, V.; Lion, C.; Biot, C.; Harduin-Lepers, A.; Guerardel, Y. Microplate sialyltransferase assay (MPSA): A rapid and sensitive assay based on an unnatural sialic acid donor and bioorthogonal chemistry. Bioconj. Chem. 2018, 29, 3377–3384. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped blast and psi-blast: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, J.; Braasch, I.; Batzel, P.; Cabau, C.; Montfort, J.; Nguyen, T.; Jouanno, E.; Berthelot, C.; Klopp, C.; Journot, L.; et al. Evolution of gene expression after whole-genome duplication: New insights from the spotted gar genome. J. Exp. Zool. B Mol. Dev. Evol. 2017, 328, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Bentrop, J.; Marx, M.; Schattschneider, S.; Rivera-Milla, E.; Bastmeyer, M. Molecular evolution and expression of zebrafish ST8SiaIII, an α-2,8-sialyltransferase involved in myotome development. Dev. Dyn. 2008, 237, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Marx, M.; Rivera-Milla, E.; Stummeyer, K.; Gerardy-Schahn, R.; Bastmeyer, M. Divergent evolution of the vertebrate polysialyltransferase stx and pst genes revealed by fish-to-mammal comparison. Dev. Biol. 2007, 306, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Marx, M.; Rutishauser, U.; Bastmeyer, M. Dual function of polysialic acid during zebrafish central nervous system development. Development 2001, 128, 4949–4958. [Google Scholar] [PubMed]
- Yanagisawa, K. Role of gangliosides in Alzheimer’s disease. Biochim. Biophys. Acta 2007, 1768, 1943–1951. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.Y.; Mir, A.M.; Thisse, C.; Guerardel, Y.; Delannoy, P.; Thisse, B.; Harduin-Lepers, A. Molecular cloning and characterization of the expression pattern of the zebrafish α2, 8-sialyltransferases (ST8Sia) in the developing nervous system. Glycoconj. J. 2009, 26, 263–275. [Google Scholar] [CrossRef]
- Garaffo, G.; Provero, P.; Molineris, I.; Pinciroli, P.; Peano, C.; Battaglia, C.; Tomaiuolo, D.; Etzion, T.; Gothilf, Y.; Santoro, M.; et al. Profiling, bioinformatic, and functional data on the developing olfactory/GnRH system reveal cellular and molecular pathways essential for this process and potentially relevant for the kallmann syndrome. Front. Endocrinol. 2013, 4, 203. [Google Scholar] [CrossRef]
- Moremen, K.W.; Ramiah, A.; Stuart, M.; Steel, J.; Meng, L.; Forouhar, F.; Moniz, H.A.; Gahlay, G.; Gao, Z.; Chapla, D.; et al. Expression system for structural and functional studies of human glycosylation enzymes. Nat. Chem. Biol. 2018, 14, 156–162. [Google Scholar] [CrossRef]
- El-Battari, A.; Prorok, M.; Angata, K.; Mathieu, S.; Zerfaoui, M.; Ong, E.; Suzuki, M.; Lombardo, D.; Fukuda, M. Different glycosyltransferases are differentially processed for secretion, dimerization, and autoglycosylation. Glycobiology 2003, 13, 941–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellokumpu, S.; Hassinen, A.; Glumoff, T. Glycosyltransferase complexes in eukaryotes: Long-known, prevalent but still unrecognized. Cell Mol. Life Sci. 2016, 73, 305–325. [Google Scholar] [CrossRef] [PubMed]
- Harduin-Lepers, A.; Stokes, D.C.; Steelant, W.F.; Samyn-Petit, B.; Krzewinski-Recchi, M.A.; Vallejo-Ruiz, V.; Zanetta, J.P.; Auge, C.; Delannoy, P. Cloning, expression and gene organization of a human Neu5Ac α2-3Galβ1-3GalNAc- α2,6-sialyltransferase: HST6GalNAc IV. Biochem. J. 2000, 352 Pt 1, 37–48. [Google Scholar]
- Rohfritsch, P.F.; Joosten, J.A.; Krzewinski-Recchi, M.A.; Harduin-Lepers, A.; Laporte, B.; Juliant, S.; Cerutti, M.; Delannoy, P.; Vliegenthart, J.F.; Kamerling, J.P. Probing the substrate specificity of four different sialyltransferases using synthetic α-d-galp-(1-->4)-β-d-glcpnac-(1-->2)-α-d-Manp-(1-->O) (ch(2))7CH3 analogues general activating effect of replacing N-acetylglucosamine by n-propionylglucosamine. Biochim. Biophys. Acta 2006, 1760, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Spiro, R.G.; Bhoyroo, V.D. Structure of the O-glycosidically linked carbohydrate units of fetuin. J. Biol. Chem. 1974, 249, 5704–5717. [Google Scholar] [PubMed]
- Takasaki, S.; Kobata, A. Asparagine-linked sugar chains of fetuin: Occurrence of tetrasialyl triantennary sugar chains containing the Gal β 1-3GlcNAc sequence. Biochemistry 1986, 25, 5709–5715. [Google Scholar] [CrossRef] [PubMed]
- Rostovtsev, V.V.; Green, L.G.; Fokin, V.V.; Sharpless, K.B. A stepwise huisgen cycloaddition process: Copper(i)-catalyzed regioselective “ligation” of azides and terminal alkynes. Angew. Chem. Int. Ed. Engl. 2002, 41, 2596–2599. [Google Scholar] [CrossRef]
- Tornoe, C.W.; Christensen, C.; Meldal, M. Peptidotriazoles on solid phase: (1,2,3)-triazoles by regiospecific copper(I)-catalyzed 1,3-dipolar cycloadditions of terminal alkynes to azides. J. Org. Chem. 2002, 67, 3057–3064. [Google Scholar] [CrossRef]
- Sun, T.; Yu, S.H.; Zhao, P.; Meng, L.; Moremen, K.W.; Wells, L.; Steet, R.; Boons, G.J. One-step selective exoenzymatic labeling (seel) strategy for the biotinylation and identification of glycoproteins of living cells. J. Am. Chem. Soc. 2016, 138, 11575–11582. [Google Scholar] [CrossRef]
- Asahina, S.; Sato, C.; Matsuno, M.; Matsuda, T.; Colley, K.; Kitajima, K. Involvement of the α2,8-polysialyltransferases II/STX and IV/PST in the biosynthesis of polysialic acid chains on the o-linked glycoproteins in rainbow trout ovary. J. Biochem. 2006, 140, 687–701. [Google Scholar] [CrossRef]
- Lehmann, F.; Kelm, S.; Dietz, F.; von Itzstein, M.; Tiralongo, J. The evolution of galactose alpha2,3-sialyltransferase: Ciona intestinalis St3Gal I/II and takifugu rubripes St3Gal II sialylate galbeta1,3galnac structures on glycoproteins but not glycolipids. Glycoconj. J. 2008, 25, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.G. Building phylogenetic trees from molecular data with mega. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, H.J.; Morris, J.H.; Ferrin, T.E.; Babbitt, P.C. Using sequence similarity networks for visualization of relationships across diverse protein superfamilies. PLoS ONE 2009, 4, e4345. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S. Evolution by Gene Duplication; Springer: Heidelberg, Germany, 1970. [Google Scholar]
- Braasch, I.; Gehrke, A.R.; Smith, J.J.; Kawasaki, K.; Manousaki, T.; Pasquier, J.; Amores, A.; Desvignes, T.; Batzel, P.; Catchen, J.; et al. The spotted gar genome illuminates vertebrate evolution and facilitates human-teleost comparisons. Nat. Genet. 2016, 48, 427–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatani, Y.; Takeda, H.; Kohara, Y.; Morishita, S. Reconstruction of the vertebrate ancestral genome reveals dynamic genome reorganization in early vertebrates. Genome Res. 2007, 17, 1254–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Yu, J.K.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muffato, M.; Louis, A.; Poisnel, C.E.; Roest Crollius, H. Genomicus: A database and a browser to study gene synteny in modern and ancestral genomes. Bioinformatics 2010, 26, 1119–1121. [Google Scholar] [CrossRef]
- Kasahara, M.; Naruse, K.; Sasaki, S.; Nakatani, Y.; Qu, W.; Ahsan, B.; Yamada, T.; Nagayasu, Y.; Doi, K.; Kasai, Y.; et al. The medaka draft genome and insights into vertebrate genome evolution. Nature 2007, 447, 714–719. [Google Scholar] [CrossRef] [Green Version]
- Lopez Aguilar, A.; Briard, J.G.; Yang, L.; Ovryn, B.; Macauley, M.S.; Wu, P. Tools for studying glycans: Recent advances in chemoenzymatic glycan labeling. ACS Chem. Biol. 2017, 12, 611–621. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Yegorov, S.; Good, S. Using paleogenomics to study the evolution of gene families: Origin and duplication history of the relaxin family hormones and their receptors. PLoS ONE 2012, 7, e32923. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book. A Guide for Laboratory Use of Zebrafish (Danio rerio); University of Oregon Press, Institute of neuroscience: Eugene, OR, USA, 1995; p. 385. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Gilormini, P.A.; Lion, C.; Vicogne, D.; Levade, T.; Potelle, S.; Mariller, C.; Guerardel, Y.; Biot, C.; Foulquier, F. A sequential bioorthogonal dual strategy: Mannal and sianal as distinct tools to unravel sialic acid metabolic pathways. Chem. Commun. 2016, 52, 2318–2321. [Google Scholar] [CrossRef] [PubMed]
- Thisse, C.; Thisse, B. High resolution whole-mount in situ hybridization. In Zebrafish Science Monitor; University of Oregon Press: Eugene, OR, USA, 1998; Volume 5, ISBN 97403-5274. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Homo sapiens | Accession Number | Danio rerio ST8Sia VI-like (AJ715551) |
|---|---|---|
| ST8Sia I | D26360 | 33.3% |
| ST8Sia II | U33551 | 29.7% |
| ST8Sia III | AF004668 | 29.9% |
| ST8Sia IV | L41680 | 27.7% |
| ST8Sia V | U91641 | 35.8% |
| ST8Sia VI | AJ621583 | 35.8% |
| ST3Gal I | L29555 | 21.4% |
| ST3Gal II | X96667 | 23.7% |
| ST3Gal III | L23768 | 20.3% |
| ST3Gal IV | L23767 | 23.6% |
| ST3Gal V | AB018356 | 24.8% |
| ST3Gal VI | AF119391 | 22.1% |
| ST6Gal I | X17247 | 19% |
| ST6Gal II | AB059555 | 17.2% |
| ST6GalNAc I | Y11339 | 16% |
| ST6GalNAc II | AJ251053 | 23.6% |
| ST6GalNAc III | AJ507291 | 16.9% |
| ST6GalNAc IV | AJ271734 | 20% |
| ST6GalNAc V | AJ507292 | 20.5% |
| ST6GalNAc VI | AJ507293 | 19.4% |
| Sialyltransferase | Homo sapiens | Danio rerio | Identity (%) |
|---|---|---|---|
| ST8Sia I | D26360 | AJ715535 | 58.7% |
| ST8Sia II | U33551 | AY055462 | 60.9% |
| ST8Sia III | AF004668 | AJ15541 | 77.9% |
| ST8Sia IV | L41680 | AJ715545 | 67.8% |
| ST8Sia V | U91641 | AJ715546 | 72.1% |
| ST8Sia VI | AJ621583 | AJ715551 | 35.8% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, L.-Y.; Teppa, E.; Noel, M.; Gilormini, P.-A.; Decloquement, M.; Lion, C.; Biot, C.; Mir, A.-M.; Cogez, V.; Delannoy, P.; et al. Novel Zebrafish Mono-α2,8-sialyltransferase (ST8Sia VIII): An Evolutionary Perspective of α2,8-Sialylation. Int. J. Mol. Sci. 2019, 20, 622. https://doi.org/10.3390/ijms20030622
Chang L-Y, Teppa E, Noel M, Gilormini P-A, Decloquement M, Lion C, Biot C, Mir A-M, Cogez V, Delannoy P, et al. Novel Zebrafish Mono-α2,8-sialyltransferase (ST8Sia VIII): An Evolutionary Perspective of α2,8-Sialylation. International Journal of Molecular Sciences. 2019; 20(3):622. https://doi.org/10.3390/ijms20030622
Chicago/Turabian StyleChang, Lan-Yi, Elin Teppa, Maxence Noel, Pierre-André Gilormini, Mathieu Decloquement, Cédric Lion, Christophe Biot, Anne-Marie Mir, Virginie Cogez, Philippe Delannoy, and et al. 2019. "Novel Zebrafish Mono-α2,8-sialyltransferase (ST8Sia VIII): An Evolutionary Perspective of α2,8-Sialylation" International Journal of Molecular Sciences 20, no. 3: 622. https://doi.org/10.3390/ijms20030622