Insertion Hot Spots of DIRS1 Retrotransposon and Chromosomal Diversifications among the Antarctic Teleosts Nototheniidae

 , ,
, ,
Abstract
:1. Introduction
2. Results
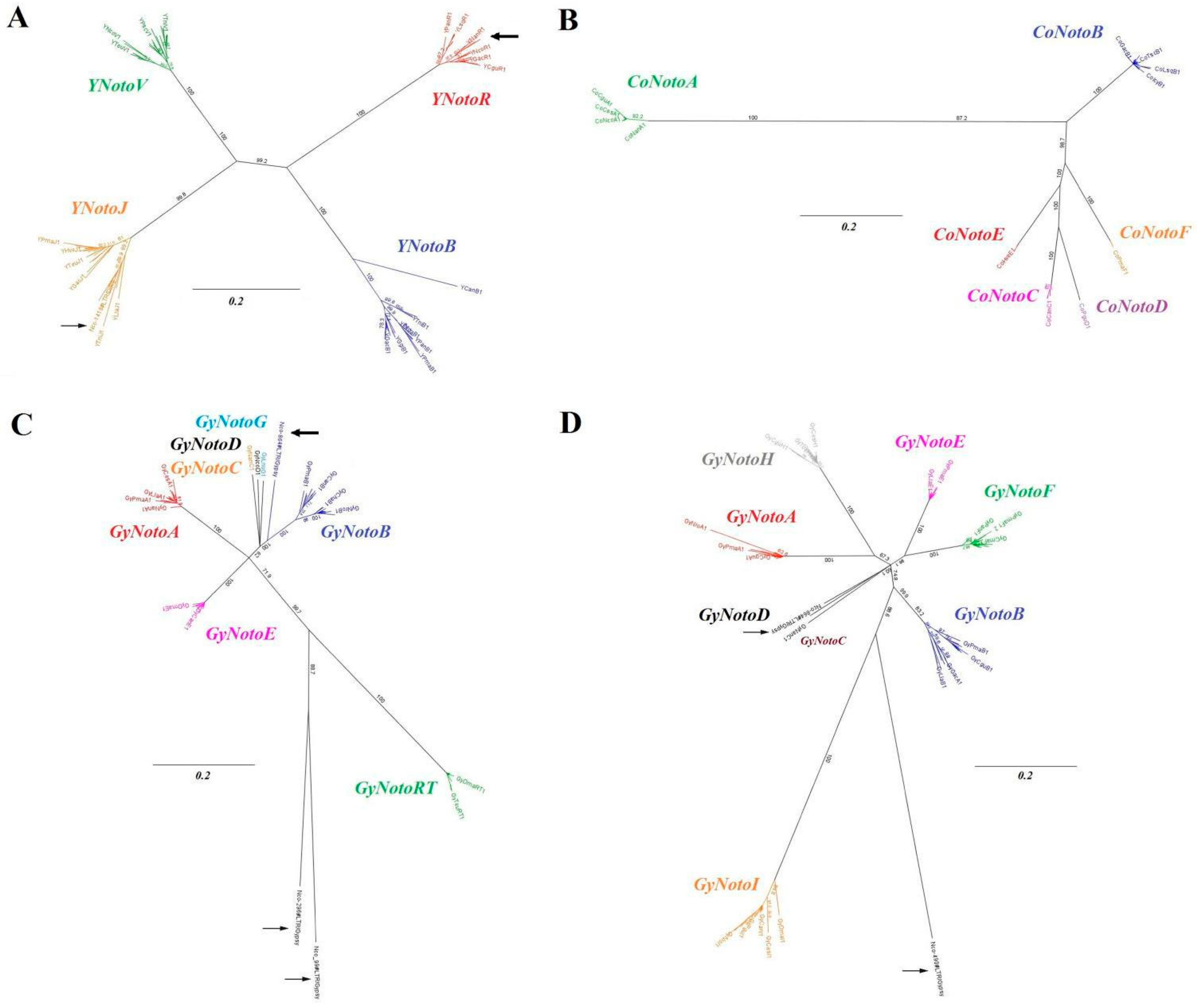
2.1. Gypsy, Copia and DIRS1 Diversity in Nototheniid Genomes
2.1.1. Identification and Distribution
2.1.2. Sequence Proximity and TE Clustering
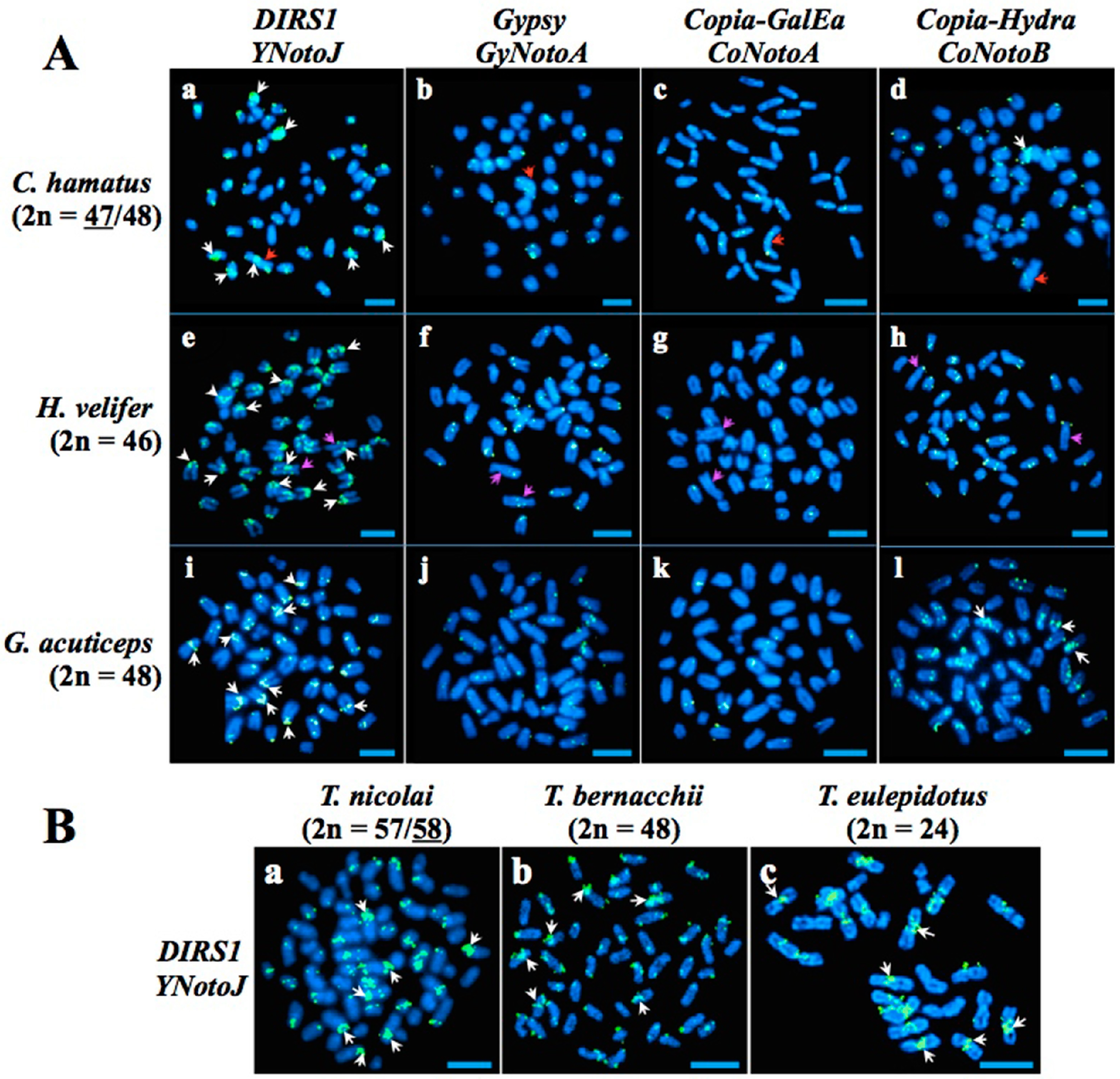
2.2. Gypsy, Copia and DIRS1 Locations on Nototheniid Chromosomes
3. Discussion
3.1. TE Diversity
3.2. A Role of the DIRS1 in the Chromosomal Fusions within the Family Nototheniidae?
3.3. Toward an Evolutionary Scenario of Nototheniid Chromosomal Speciations
4. Materials and Methods
4.1. Fish Specimens
4.2. Sample Collection and Preparation
4.2.1. Tissues for DNA Analyses
4.2.2. Chromosome Preparations
4.3. TE Amplification in Nototheniid Genomes
4.4. FISH
4.4.1. TE Probe Preparation
4.4.2. FISH with TE Probes
4.4.3. Image Acquisition and Karyotyping
4.5. Ethics Approval and Consent to Participate
4.6. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TE | Transposable Element |
| DNA | Deoxyribonucleic Acid |
| YNoto | DIRS1 elements of nototheniid genomes |
| GyNoto | Gypsy elements of nototheniid genomes |
| CoNoto | Copia elements of nototheniid genomes |
| RT | Reverse Transcriptase |
| RH | RNAseH |
| Int | Integrase |
| LTR | Long Terminal Repeat |
| YR | Tyrosine Recombinase |
| DIRS | Dictyostelium Intermediate Repeat Sequence |
| My | Million years |
| FISH | Fluorescent in-Situ Hybridization |
| DAPI | 4′,6-diamidino-2-phenylindole |
| FITC | Fluorescein Isothiocyanate |
| PCR | Polymerization Chain Reaction |
| BLAST | Basic Local Alignment Search Tool |
| MAFFT | Multiple Alignment using Fast Fourier Transform |
| NJ | Neighbor Joining |
| ML | Maximum Likelyhood |
| HCl | Hydrochloric acid |
| SSC | Saline Sodium Citrate |
References
- McClintock, B. The Significance of Responses of the Genome to Challenge. Science 1984, 226, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Böhne, A.; Brunet, F.; Galiana-Arnoux, D.; Schultheis, C.; Volff, J.-N. Transposable elements as drivers of genomic and biological diversity in vertebrates. Chromosome Res. 2008, 16, 203–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, I.A.; Naville, M.; Chalopin, D.; Levin, P.; Berger, C.S.; Galiana, D.; Volff, J.-N. Evolutionary impact of transposable elements on genomic diversity and lineage-specific innovation in vertebrates. Chromosome Res. 2015, 23, 505–531. [Google Scholar] [CrossRef] [PubMed]
- Raskina, O.; Barber, J.C.; Nevo, E.; Belyayev, A. Repetitive DNA and chromosomal rearrangements: Speciation-related events in plant genomes. Cytogenet. Genome Res. 2008, 120, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Biémont, C.; Vieira, C. What transposable elements tell us about genome organization and evolution: The case of Drosophila. Cytogenet. Genome Res. 2005, 110, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Volff, J.-N. Genome evolution and biodiversity in teleost fish. Heredity 2005, 94, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.; Bohlen, J.; Freyhof, J.; Rábová, M.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42. [Google Scholar] [CrossRef]
- Mahé, F. Phylogénie, Éléments Transposables et Évolution de la Taille des Génomes chez les Lupins. Ph.D Thesis, Université Rennes 1, Rennes, France, 2009. [Google Scholar]
- Ferreira, I.A.; Poletto, A.B.; Kocher, T.D.; Mota-Velasco, J.C.; Penman, D.J.; Martins, C. Chromosome Evolution in African Cichlid Fish: Contributions from the Physical Mapping of Repeated DNAs. Cytogenet. Genome Res. 2010, 129, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Eastman, J.T. The nature of the diversity of Antarctic fishes. Polar Biol. 2005, 28, 93–107. [Google Scholar] [CrossRef]
- Cheng, C.; Detrich, H.W. Molecular ecophysiology of Antarctic notothenioid fishes. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 2215–2232. [Google Scholar] [CrossRef] [Green Version]
- Rutschmann, S.; Matschiner, M.; Damerau, M.; Muschick, M.; Lehmann, M.F.; Hanel, R.; Salzburger, W. Parallel ecological diversification in Antarctic notothenioid fishes as evidence for adaptive radiation: Ecological diversification in Notothenioids. Mol. Ecol. 2011, 20, 4707–4721. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.; McCune, A.R. Fishes on the Antarctic continental shelf: Evolution of a marine species flock? J. Fish Biol. 2000, 57, 84–102. [Google Scholar] [CrossRef]
- De Vries, A.L. Glycoproteins as Biological Antifreeze Agents in Antarctic Fishes. Science 1971, 172, 1152–1155. [Google Scholar] [CrossRef]
- De Vries, A. Biological antifreezes and survival in freezing environments. In Animals and Environmental Fitness; Gilles R: Oxford, UK, 1980; p. 583. [Google Scholar]
- Near, T.J.; Dornburg, A.; Kuhn, K.L.; Eastman, J.T.; Pennington, J.N.; Patarnello, T.; Zane, L.; Fernandez, D.A.; Jones, C.D. Ancient climate change, antifreeze, and the evolutionary diversification of Antarctic fishes. Proc. Natl. Acad. Sci. USA 2012, 109, 3434–3439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Near, T.J.; MacGuigan, D.J.; Parker, E.; Struthers, C.D.; Jones, C.D.; Dornburg, A. Phylogenetic analysis of Antarctic notothenioids illuminates the utility of RADseq for resolving Cenozoic adaptive radiations. Mol. Phylogenet. Evol. 2018, 129, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Dettai, A.; Berkani, M.; Lautredou, A.-C.; Couloux, A.; Lecointre, G.; Ozouf-Costaz, C.; Gallut, C. Tracking the elusive monophyly of nototheniid fishes (Teleostei) with multiple mitochondrial and nuclear markers. Mar. Genomics 2012, 8, 49–58. [Google Scholar] [CrossRef]
- Colombo, M.; Damerau, M.; Hanel, R.; Salzburger, W.; Matschiner, M. Diversity and disparity through time in the adaptive radiation of Antarctic notothenioid fishes. J. Evol. Biol. 2015, 28, 376–394. [Google Scholar] [CrossRef] [Green Version]
- Auvinet, J.; Graça, P.; Belkadi, L.; Petit, L.; Bonnivard, E.; Dettaï, A.; Detrich, W.H.; Ozouf-Costaz, C.; Higuet, D. Mobilization of retrotransposons as a cause of chromosomal diversification and rapid speciation: The case for the Antarctic teleost genus Trematomus. BMC Genomics 2018, 19, 339. [Google Scholar] [CrossRef]
- Lautrédou, A.-C.; Hinsinger, D.D.; Gallut, C.; Cheng, C.-H.C.; Berkani, M.; Ozouf-Costaz, C.; Cruaud, C.; Lecointre, G.; Dettai, A. Phylogenetic footprints of an Antarctic radiation: The Trematominae (Notothenioidei, Teleostei). Mol. Phylogenet. Evol. 2012, 65, 87–101. [Google Scholar] [CrossRef]
- Martens, K. Speciation in ancient lakes. Tree 1997, 12, 177–182. [Google Scholar] [CrossRef]
- Lecointre, G.; Améziane, N.; Boisselier, M.-C.; Bonillo, C.; Busson, F.; Causse, R.; Chenuil, A.; Couloux, A.; Coutanceau, J.-P.; Cruaud, C.; et al. Is the Species Flock Concept Operational? The Antarctic Shelf Case. PLoS ONE 2013, 8, e68787. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.B.; Shipp, S.S.; Lowe, A.L.; Wellner, J.S.; Mosola, A.B. The Antarctic Ice Sheet during the Last Glacial Maximum and its subsequent retreat history: A review. Quat. Sci. Rev. 2002, 21, 49–70. [Google Scholar] [CrossRef]
- Huybrechts, P. Sea-level changes at the LGM from ice-dynamic reconstructions of the Greenland and Antarctic ice sheets during the glacial cycles. Quat. Sci. Rev. 2002, 21, 203–231. [Google Scholar] [CrossRef] [Green Version]
- Thatje, S.; Hillenbrand, C.-D.; Larter, R. On the origin of Antarctic marine benthic community structure. Trends Ecol. Evol. 2005, 20, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Smale, D.; Barnes, D.; Fraser, K.; Peck, L. Benthic community response to iceberg scouring at an intensely disturbed shallow water site at Adelaide Island, Antarctica. Mar. Ecol. Prog. Ser. 2008, 355, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Ozouf-Costaz, C.; Hureau, J.-C.; Beaunier, M. Chromosome studies on fish of the suborder Notothenioidei collected in the Weddell sea during EPOS 3 cruise. Cybium 1991, 15, 271–289. [Google Scholar]
- Pisano, E.; Ozouf-Costaz, C. Chromosome change and the evolution in the Antarctic fish suborder Notothenioidei. Antarctic Sci. 2000, 12, 334–342. [Google Scholar] [CrossRef]
- Ghigliotti, L.; Cheng, C.C.-H.; Ozouf-Costaz, C.; Vacchi, M.; Pisano, E. Cytogenetic diversity of notothenioid fish from the Ross sea: Historical overview and updates. Hydrobiologia 2015, 761, 373–396. [Google Scholar] [CrossRef]
- Molina, W.F. Chromosomal changes and stasis in marine fish groups. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Forest, F., Kapoor, B.G., Eds.; Enfield: London, UK, 2006; pp. 69–110. [Google Scholar]
- Molina, W.F.; de F. Bacurau, T.O. Structural and Numerical Chromosome Diversification in Marine Perciformes (Priacanthidae and Gerreidae). Cytologia 2006, 71, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Pisano, E.; Cocca, E.; Mazzei, F.; Ghigliotti, L.; Di Prisco, G.; Detrich III, H.W.; Ozouf-Costaz, C. Mapping of α-and β-globin genes on Antarctic fish chromosomes by fluorescence in-situ hybridization. Chromosome Res. 2003, 11, 633–640. [Google Scholar] [CrossRef]
- Mazzei, F.; Ghigliotti, L.; Lecointre, G.; Ozouf-Costaz, C.; Coutanceau, J.-P.; Detrich, W.; Pisano, E. Karyotypes of basal lineages in notothenioid fishes: The genus Bovichtus. Polar Biol. 2006, 29, 1071–1076. [Google Scholar] [CrossRef]
- Mazzei, F.; Ghigliotti, L.; Coutanceau, J.-P.; Detrich, H.W.; Prirodina, V.; Ozouf-Costaz, C.; Pisano, E. Chromosomal characteristics of the temperate notothenioid fish Eleginops maclovinus (Cuvier). Polar Biol. 2008, 31, 629–634. [Google Scholar] [CrossRef]
- Morescalchi, A.; Pisano, E.; Stanyon, R.; Morescalchi, M.A. Cytotaxonomy of antarctic teleosts of the Pagothenia/Trematomus complex (Nototheniidae, Perciformes). Polar Biol. 1992, 12, 553–558. [Google Scholar] [CrossRef]
- Morescalchi, A.; Hureau, J.C.; Olmo, E.; Ozouf-Costaz, C.; Pisano, E.; Stanyon, R. A multiple sex-chromosome system in Antarctic ice-fishes. Polar Biol. 1992, 11, 655–661. [Google Scholar] [CrossRef]
- Ghigliotti, L.; Cheng, C.-H.C.; Bonillo, C.; Coutanceau, J.-P.; Pisano, E. In Situ Gene Mapping of Two Genes Supports Independent Evolution of Sex Chromosomes in Cold-Adapted Antarctic Fish. BioMed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pisano, E.; Angelini, C.; Mazzei, F.; Stanyon, R. Adaptive radiation in Antarctic notothenioid fish: Studies of genomic change at chromosomal level. Ital. J. Zool. 2000, 67, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Ozouf-Costaz, C.; Pisano, E.; Thaeron, C.; Hureau, J.-C. Antarctic fish chromosome banding: Significance for evolutionary studies. Cybium 1997, 21, 399–409. [Google Scholar]
- Kazazian, H.H. Mobile Elements: Drivers of Genome Evolution. Science 2004, 303, 1626–1632. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.R.; Greene, W.K. Transposable elements: Powerful facilitators of evolution. BioEssays 2009, 31, 703–714. [Google Scholar] [CrossRef]
- Rebollo, R.; Horard, B.; Hubert, B.; Vieira, C. Jumping genes and epigenetics: Towards new species. Gene 2010, 454, 1–7. [Google Scholar] [CrossRef]
- Aparicio, S.; Chapman, J.; Stupka, E.; Putman, N.; Chia, J. Whole-Genome Shotgun Assembly and Analysis of the Genome of Fugu rubripes. Science 2002, 297, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, M.; Naruse, K.; Sasaki, S.; Nakatani, Y.; Qu, W.; Ahsan, B.; Yamada, T.; Nagayasu, Y.; Doi, K.; Kasai, Y.; et al. The medaka draft genome and insights into vertebrate genome evolution. Nature 2007, 447, 714–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.-N. Comparative Analysis of Transposable Elements Highlights Mobilome Diversity and Evolution in Vertebrates. Genome Biol. Evol. 2015, 7, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotero-Caio, C.G.; Platt, R.N.; Suh, A.; Ray, D.A. Evolution and Diversity of Transposable Elements in Vertebrate Genomes. Genome Biol. Evol. 2017, 9, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Elgar, G. Generation and Analysis of 25 Mb of Genomic DNA from the Pufferfish Fugu rubripes by Sequence Scanning. Genome Res. 1999, 9, 960–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crollius, H.R. Characterization and Repeat Analysis of the Compact Genome of the Freshwater Pufferfish Tetraodon nigroviridis. Genome Res. 2000, 10, 939–949. [Google Scholar] [CrossRef] [Green Version]
- Volff, J.-N.; Bouneau, L.; Ozouf-Costaz, C.; Fischer, C. Diversity of retrotransposable elements in compact pufferfish genomes. Trends Genet. 2003, 19, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Duvernell, D.D.; Pryor, S.R.; Adams, S.M. Teleost Fish Genomes Contain a Diverse Array of L1 RetrotransposonLineages That Exhibit a Low Copy Number and High Rate of Turnover. J. Mol. Evol. 2004, 59, 298–308. [Google Scholar] [CrossRef]
- Brawand, D.; Wagner, C.E.; Li, Y.I.; Malinsky, M.; Keller, I.; Fan, S.; Simakov, O.; Ng, A.Y.; Lim, Z.W.; Bezault, E.; et al. The genomic substrate for adaptive radiation in African cichlid fish. Nature 2014, 513, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Braasch, I.; Gehrke, A.R.; Smith, J.J.; Kawasaki, K.; Manousaki, T.; Pasquier, J.; Amores, A.; Desvignes, T.; Batzel, P.; Catchen, J.; et al. The spotted gar genome illuminates vertebrate evolution and facilitates human-teleost comparisons. Nat. Genet. 2016, 48, 427–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detrich, H.W.; Stuart, A.; Schoenborn, M.; Parker, S.K.; Methé, B.A.; Amemiya, C.T. Genome enablement of the notothenioidei: Genome size estimates from 11 species and BAC libraries from 2 Representative Taxa. J. Exp. Zool. Part B Mol. Dev. Evol. 2010, 314B, 369–381. [Google Scholar] [CrossRef]
- Detrich, H.W.; Amemiya, C.T. Antarctic Notothenioid Fishes: Genomic Resources and Strategies for Analyzing an Adaptive Radiation. Integr. Comp. Biol. 2010, 50, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Capriglione, T.; De Paolo, S.; Cocca, E. Helinoto, a Helitron2 transposon from the icefish Chionodraco hamatus, contains a region with three deubiquitinase-like domains that exhibit transcriptional activity. Comp. Biochem. Physiol. Part D Genomics Proteomics 2014, 11, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Capriglione, T.; Odierna, G.; Caputo, V.; Canapa, A.; Olmo, E. Characterization of a Tc1-like transposon in the Antarctic ice-fish, Chionodraco hamatus. Gene 2002, 295, 193–198. [Google Scholar] [CrossRef]
- Ozouf-Costaz, C.; Brandt, J.; Körting, C.; Pisano, E.; Bonillo, C.; Coutanceau, J.-P.; Volff, J.-N. Genome dynamics and chromosomal localization of the non-LTR retrotransposons Rex1 and Rex3 in Antarctic fish. Antarctic Sci. 2004, 16, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Eickbush, T.H.; Jamburuthugoda, V.K. The diversity of retrotransposons and the properties of their reverse transcriptases. Virus Res. 2008, 134, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Eickbush, T.H.; Malik, H.S. Origins and Evolution of Retrotransposons. In Mobile DNA II; Craig, N.L., Lambowitz, A.M., Craigie, R., Gellert, M., Eds.; American Society of Microbiology: Washington, DC, USA, 2002; pp. 1111–1144. ISBN 978-1-55581-209-6. [Google Scholar]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef]
- Duhamel, G.; Hulley, P.; Causse, R.; Koubbi, P.; Vacchi, M.; Pruvost, P.; Vigetta, S.; Irisson, J.; Mormède, S.; Belchier, M.; et al. Biogegographic atlas of the Southern Ocean; De Broyer, C., Koubbi, P., Griffiths, H.J., Raymond, B., Udekem d’Acoz, C., Eds.; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 328–352. [Google Scholar]
- Terrat, Y.; Bonnivard, E.; Higuet, D. GalEa retrotransposons from galatheid squat lobsters (Decapoda, Anomura) define a new clade of Ty1/copia-like elements restricted to aquatic species. Mol. Genet. Genomics 2008, 279, 63–73. [Google Scholar] [CrossRef]
- Llorens, C.; Muñoz-Pomer, A.; Bernad, L.; Botella, H.; Moya, A. Network dynamics of eukaryotic LTR retroelements beyond phylogenetic trees. Biol. Direct 2009, 4, 41. [Google Scholar] [CrossRef]
- Shao, F.; Wang, J.; Xu, H.; Peng, Z. FishTEDB: A collective database of transposable elements identified in the complete genomes of fish. Database 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.C.; Ahn, D.H.; Kim, S.J.; Pyo, C.W.; Lee, H.; Kim, M.-K.; Lee, J.; Lee, J.E.; Detrich, H.W.; Postlethwait, J.H. The genome sequence of the Antarctic bullhead notothen reveals evolutionary adaptations to a cold environment. Genome Biol. 2014, 15, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisano, E.; Mazzei, F.; Derome, N.; Ozouf-Costaz, C.; Hureau, J.; Di Prisco, G. Cytogenetics of the bathydraconid fish Gymnodraco acuticeps (Perciformes, Notothenioidei) from Terra Nova Bay, Ross Sea. Polar Biol. 2001, 24, 846–852. [Google Scholar]
- Caputo, V.; Splendiani, A.; Nisi Cerioni, P.; Olmo, E. The chromosomal complement of the artedidraconid fish Histiodraco velifer (Perciformes: Notothenioidei) from Terra Nova Bay, Ross Sea. Cytogenet. Genome Res. 2003, 101, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Morescalchi, A.; Morescalchi, M.A.; Odierna, G.; Sitingo, V.; Capriglione, T. Karyotype and genome size of zoarcids and notothenioids (Taleostei, Perciformes) from the Ross Sea: Cytotaxonomic implications. Polar Biol. 1996, 16, 559–564. [Google Scholar] [CrossRef]
- Amores, A.; Wilson, C.A.; Allard, C.A.H.; Detrich, H.W.; Postlethwait, J.H. Cold Fusion: Massive Karyotype Evolution in the Antarctic Bullhead Notothen Notothenia coriiceps. G3 2017, 7, 2195–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Valen, L. A New Evolutionary Law; The University of Chicago: Chicago, IL, USA, 1973; Volume 1. [Google Scholar]
- Matschiner, M.; Hanel, R.; Salzburger, W. On the Origin and Trigger of the Notothenioid Adaptive Radiation. PLoS ONE 2011, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Betancur, R.; Wiley, E.O.; Arratia, G.; Acero, A.; Bailly, N.; Miya, M.; Lecointre, G.; Orti, G. Phylogenetic classification of bony fishes. BMC Evol. Biol. 2017, 17, 1–40. [Google Scholar] [CrossRef]
- Fontdevila, A. Hybrid genome evolution by transposition. Cytogenet. Genome Res. 2005, 110, 49–55. [Google Scholar] [CrossRef]
- Piednoël, M.; Donnart, T.; Esnault, C.; Graça, P.; Higuet, D.; Bonnivard, E. LTR-Retrotransposons in R. exoculata and Other Crustaceans: The Outstanding Success of GalEa-Like Copia Elements. PLoS ONE 2013, 8, e57675. [Google Scholar] [CrossRef] [Green Version]
- Ahn, D.-H.; Shin, S.C.; Kim, B.-M.; Kang, S.; Kim, J.-H.; Ahn, I.; Park, J.; Park, H. Draft genome of the Antarctic dragonfish, Parachaenichthys charcoti. GigaScience 2017, 6, 1–6. [Google Scholar] [CrossRef]
- Devos, K.M.; Brown, J.K.; Bennetzen, J.L. Genome size reduction through illegitimate recombination counteracts genome expansion in Arabidopsis. Genome Res. 2002, 12, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Bonnivard, E.; Higuet, D. Fluidity of eukaryotic genomes. C. R. Biol. 2009, 332, 234–240. [Google Scholar] [CrossRef]
- Lönnig, W.-E.; Saedler, H. Chromosome Rearrangements and Transposable Elements. Annu. Rev. Genet. 2002, 36, 389–410. [Google Scholar] [CrossRef] [PubMed]
- Kehrer-Sawatzki, H.; Cooper, D.N. Understanding the recent evolution of the human genome: Insights from human-chimpanzee genome comparisons. Hum. Mutat. 2007, 28, 99–130. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.T.; Mazzuchelli, J.; Ferreira, I.A.; Poletto, A.B.; Fantinatti, B.E.A.; Martins, C. Cytogenetic Mapping of the Retroelements Rex1, Rex3 and Rex6 among Cichlid Fish: New Insights on the Chromosomal Distribution of Transposable Elements. Cytogenet. Genome Res. 2011, 133, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Kraaijeveld, K. Genome Size and Species Diversification. Evol. Biol. 2010, 37, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Chénais, B.; Caruso, A.; Hiard, S.; Casse, N. The impact of transposable elements on eukaryotic genomes: From genome size increase to genetic adaptation to stressful environments. Gene 2012, 509, 7–15. [Google Scholar] [CrossRef]
- Kennett, J. Miocene Paleoceanography and Plankton Evolution. In Mesozoic and Cenozoic Oceans; Hsü, K.J., Ed.; American Geophysical Union: Washington, DC, USA, 1986; Volume 15, pp. 19–122. [Google Scholar]
- Kennett, J.; Barker, P. Latest Cretaceous to Cenozoic climate and oceanographic development in the Weddell Sea: An ocean drilling perspective. In Proceedings of the Ocean Drilling Program, Scientific Results; Ocean Drilling Program: College Station, TX, USA, 1990; Volume 113, pp. 937–960. [Google Scholar]
- Stott, L.D.; Kennett, J.P.; Shackleton, N.J.; Corfield, R.M. The evolution of antarctic surface waters during the paleogene: Inferences from the stable isotopic composition of planktonic foraminifers. In Proceedings of the Ocean Drilling Program, Scientific Results; Ocean Drilling Program: College Station, TX, USA, 1990; Volume 113, pp. 849–863. [Google Scholar]
- Cappello, J.; Zuker, C.; Lodish, H.F. Repetitive Dictyostelium heat-shock promotor functions in Saccharomyces cerevisiae. Mol. Cell. Biol. 1984, 4, 591–598. [Google Scholar] [CrossRef]
- Westphal, T.; Reuter, G. Recombinogenic Effects of Suppressors of Position-Effect Variegation in Drosophila. Genetics 2002, 609–621. [Google Scholar]
- White, M.J.D. Chain Processes in Chromosomal Speciation. Syst. Zool. 1978, 27, 285. [Google Scholar] [CrossRef]
- Bush, G. Modes of Animal Speciation. Annu. Rev. Ecol. Syst. 1975, 6, 339–364. [Google Scholar] [CrossRef]
- Baker, R.J.; Bickham, J.W. Speciation by monobrachial centric fusions. Proc. Natl. Acad. Sci. USA 1986, 83, 8245–8248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieseberg, L.H. Chromosomal rearrangements and speciation. Trends Ecol. Evol. 2001, 16, 351–358. [Google Scholar] [CrossRef]
- Barton, N.H.; Rouhani, S. The probability of fixation of a new karyotype in a continuous population. Evolution 1991, 45, 499–517. [Google Scholar] [CrossRef] [PubMed]
- Garagna, S.; Marziliano, N.; Zuccotti, M.; Searle, J.B.; Capanna, E.; Redi, C.A. Pericentromeric organization at the fusion point of mouse Robertsonian translocation chromosomes. Proc. Natl. Acad. Sci. USA 2001, 98, 171–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, A.; Johnston, I. Evolution and adaptive radiation of Antarctic fishes. Tree 1996, 11, 212–218. [Google Scholar] [CrossRef]
- Britton-Davidian, J.; Catalan, J.; da Graça Ramalhinho, M.; Ganem, G.; Auffray, J.-C.; Capela, R.; Biscoito, M.; Searle, J.B.; da Luz Mathias, M. Rapid chromosomal evolution in island mice: Environmental genetics. Nature 2000, 403, 158. [Google Scholar] [CrossRef]
- Auffray, J.-C. Developmental Stability in House Mice Heterozygous for Single Robertsonian Fusions. J. Heredity 2001, 92, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Eastman, J.T. Bathymetric distributions of notothenioid fishes. Polar Biol. 2017, 40, 2077–2095. [Google Scholar] [CrossRef]
- Eastman, J.T.; DeVries, A.L. Adaptations for cryopelagic life in the antarctic notothenioid fish Pagothenia borchgrevinki. Polar Biol. 1985, 4, 45–52. [Google Scholar] [CrossRef]
- Murphy, K.R. Investigating the Larval/Juvenile Notothenioïd Fish Species Assemblage in McMurdo Sound, Antarctica. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Champaign, IL, USA, 2015. [Google Scholar]
- Winnepennincks, B.; Backeljau, T.; De Watcher, R. Extraction of high molecular weight DNA from molluscs. Trends Genet. 1993, 9, 407. [Google Scholar]
- Doussau de Bazignan, M.; Ozouf-Costaz, C. Une technique rapide d’analyse chromosomique appliquée à sept espèces de poissons antarctiques. Cybium 1985, 9, 57–74. [Google Scholar]
- Rey, O.; d’Hont, A.; Coutanceau, J.; Pisano, E.; Chilmonczyk, S.; Ozouf-Costaz, C. Cephalic Kidney and Spleen Cell Culture in Antarctic Teleosts. In Fish Cytogenetic Techniques; Ozouf-Costaz, C., Pisano, E., Foresti, F., de Almeida, L., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 74–81. ISBN 978-1-4822-1198-6. [Google Scholar]
- Alva, V.; Nam, S.-Z.; Söding, J.; Lupas, A.N. The MPI bioinformatics Toolkit as an integrative platform for advanced protein sequence and structure analysis. Nucleic Acids Res. 2016, 44, W410–W415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef]
- Bonillo, C.; Coutanceau, J.P.; D’Cotta, H.; Ghigliotti, L.; Ozouf-Costaz, C.; Pisano, E. Standard Fluorescence In Situ Hybridization Procedures; Fish Cytogenetic Techniques: Ray-Fin Fishes and Chondrichthyans; CRC Press: Boca Raton, FL, USA, 2015; Volume 103. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TE Super-Families | DIRS1 (YNoto) | Gyspy (GyNoto) | Copia (CoNoto) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TE Families | YB | YJ | YR | YV | GyA | GyB | GyE | GyF | GyH | GyI | GyRT | CoA | CoB | Other |
| Species/TE Regions | RT/RH | RH/Int | Int | RT | RT/RH | RT/RH | ||||||||
| PLEURAGRAMMINAE | ||||||||||||||
| Pleuragramma antarctica | + | + | + | + | − | + | − | + | + | + | + | − | + | |
| DISSOSTICHINAE | ||||||||||||||
| Dissostichus mawsoni * | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| TREMATOMINAE | ||||||||||||||
| Trematomus bernacchii * | + | + | + | + | + | + | + | + | + | + | + | − | + | |
| Trematomus Nicolai * | + | + | + | + | + | + | + | + | + | + | + | − | + | |
| Trematomus eulepidotus * | + | + | + | + | + | + | + | + | + | + | + | − | + | |
| Lepidonotothen nudifrons | + | + | + | + | + | + | − | + | + | + | + | − | + | |
| Lepidonotothen larseni | + | + | + | + | + | + | + | + | + | + | + | − | + | |
| Lepidonotothen squamifrons | + | + | + | + | + | + | + | + | + | + | + | − | + | |
| Patagonotothen guentheri | + | + | + | + | + | + | + | + | + | + | + | − | − | CoNotoD |
| GOBIONOTOTHENIINAE | ||||||||||||||
| Gobionotothen gibberifrons | + | + | + | + | + | + | − | + | + | + | + | + | − | CoNotoC |
| NOTOTHENIINAE | ||||||||||||||
| Notothenia coriiceps * | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Notothenia angustata | + | + | + | + | + | + | + | + | + | + | + | + | − | CoNotoF |
| Notothenia rossii | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Paranotothenia magellanica | + | + | + | + | + | + | + | + | + | + | + | − | − | CoNotoF |
| ARTEDIDRACONINAE | ||||||||||||||
| Pogonophryne scotti | + | + | + | + | + | − | + | + | + | + | + | + | + | |
| Histiodraco velifer | + | + | + | + | + | + | + | + | + | + | + | − | − | CoNotoE |
| GYMNODRACONINAE | ||||||||||||||
| Gymnodraco acuticeps | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| CHANNICHTHYINAE | ||||||||||||||
| Chionodraco hamatus | + | + | + | + | + | + | + | + | + | + | − | − | − | |
| Cryodraco antarcticus | + | + | + | + | + | + | + | + | + | + | − | + | − | CoNotoC |
| Champsocephalus gunnari | + | + | + | + | + | + | + | + | + | + | − | + | − | CoNotoC |
| Champsocephalus esox | + | + | + | + | + | + | + | + | + | + | − | + | − | CoNotoC |
| CYGNODRACONINAE | ||||||||||||||
| Cygnodraco mawsoni | + | + | + | + | + | + | + | + | + | + | + | − | − | CoNotoE |
| Clades | TEs | Trem | Pleu | Noto | Diss | Chan | Cygn | Gymn | Arte | Gobi |
|---|---|---|---|---|---|---|---|---|---|---|
| Trem | YNoto | 93.2 | 86.9 | 87.1 | 85.9 | 87.8 | 88.5 | 89.3 | 88.2 | 89.0 |
| GyNoto | 93.8 | 90.0 | 91.9 | 92.1 | 92.4 | 93.6 | 91.4 | 92.6 | 91.2 | |
| CoNoto | 96.0 | 95.3 | 95.8 | 92.2 | NA | NA | 97.1 | 88.4 | NA | |
| Pleu | YNoto | NA | 85.1 | 83.0 | 85.9 | 86.3 | 90.0 | 86.4 | 85.5 | |
| GyNoto | NA | 88.5 | 84.5 | 89.0 | 95.0 | 82.0 | 84.1 | 96.3 | ||
| CoNoto | NA | 95.8 | 92.1 | NA | NA | 96.5 | 88.2 | NA | ||
| Noto | YNoto | 87.4 | 88.9 | 86.4 | 88.5 | 87.4 | 89.7 | 89.5 | ||
| GyNoto | 90.2 | 91.0 | 92.1 | 92.3 | 91.5 | 92.6 | 90.5 | |||
| CoNoto | 96.4 | 92.6 | NA | NA | 96.9 | 89.7 | NA | |||
| Diss | YNoto | NA | 85.1 | 85.6 | 89.5 | 86.6 | 82.0 | |||
| GyNoto | NA | 92.1 | 92.8 | 92.8 | 93.0 | 88.8 | ||||
| CoNoto | NA | NA | NA | 96.2 | 86.7 | NA | ||||
| Chan | YNoto | 86.5 | 91.2 | 90.3 | 92.0 | 86.5 | ||||
| GyNoto | 92.7 | 93.7 | 92.6 | 93.5 | 91.3 | |||||
| CoNoto | NA | NA | NA | NA | NA | |||||
| Cygn | YNoto | NA | 94.6 | 93.2 | 91.0 | |||||
| GyNoto | NA | 90.0 | 93.3 | 92.0 | ||||||
| CoNoto | NA | NA | NA | NA | ||||||
| Gymn | YNoto | NA | 91.9 | 86.0 | ||||||
| GyNoto | NA | 96.7 | 82.8 | |||||||
| CoNoto | NA | 89.5 | NA | |||||||
| Arte | YNoto | 95.5 | 89.5 | |||||||
| GyNoto | 97.3 | 89.3 | ||||||||
| CoNoto | NA | NA | ||||||||
| Gobi | YNoto | NA | ||||||||
| GyNoto | NA | |||||||||
| CoNoto | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Auvinet, J.; Graça, P.; Ghigliotti, L.; Pisano, E.; Dettaï, A.; Ozouf-Costaz, C.; Higuet, D. Insertion Hot Spots of DIRS1 Retrotransposon and Chromosomal Diversifications among the Antarctic Teleosts Nototheniidae. Int. J. Mol. Sci. 2019, 20, 701. https://doi.org/10.3390/ijms20030701
Auvinet J, Graça P, Ghigliotti L, Pisano E, Dettaï A, Ozouf-Costaz C, Higuet D. Insertion Hot Spots of DIRS1 Retrotransposon and Chromosomal Diversifications among the Antarctic Teleosts Nototheniidae. International Journal of Molecular Sciences. 2019; 20(3):701. https://doi.org/10.3390/ijms20030701
Chicago/Turabian StyleAuvinet, Juliette, Paula Graça, Laura Ghigliotti, Eva Pisano, Agnès Dettaï, Catherine Ozouf-Costaz, and Dominique Higuet. 2019. "Insertion Hot Spots of DIRS1 Retrotransposon and Chromosomal Diversifications among the Antarctic Teleosts Nototheniidae" International Journal of Molecular Sciences 20, no. 3: 701. https://doi.org/10.3390/ijms20030701