New Insights on Arabidopsis thaliana Root Adaption to Ammonium Nutrition by the Use of a Quantitative Proteomic Approach

 ,
,
Abstract
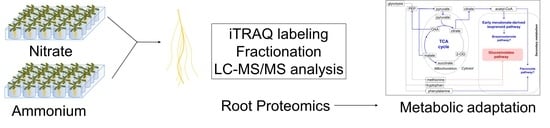
:
1. Introduction
2. Results and Discussion
2.1. Physiological Response of Arabidopsis Roots under Ammonium Nutrition
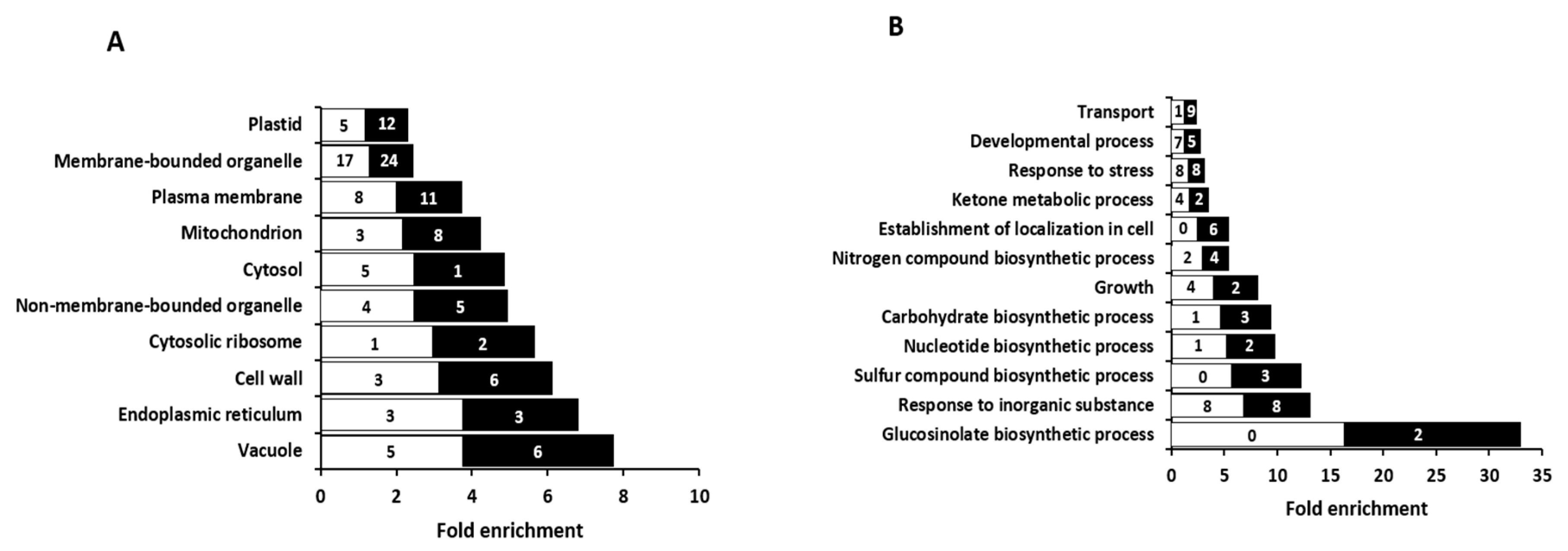
2.2. Overview of Proteomic Analysis in Arabidopsis Roots Grown under Exclusive Nitrate or Ammonium Supply
2.3. Glucosinolate Biosynthesis is Modulated by Ammonium or Nitrate as N Source
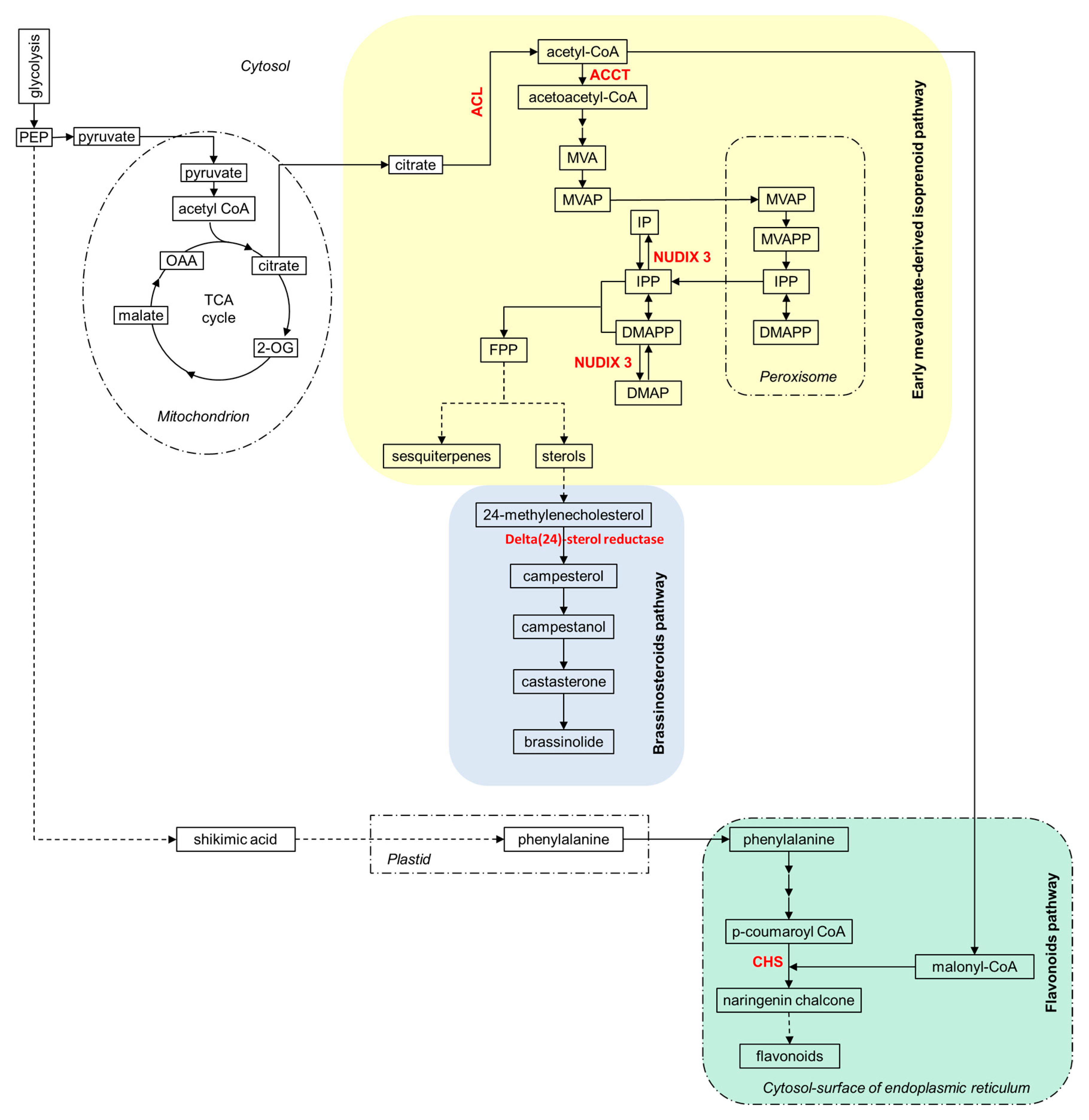
2.4. Ammonium Nutrition and Secondary Metabolism in Arabidopsis Roots: Brassinosteroids and Hormonal Signaling Pathways
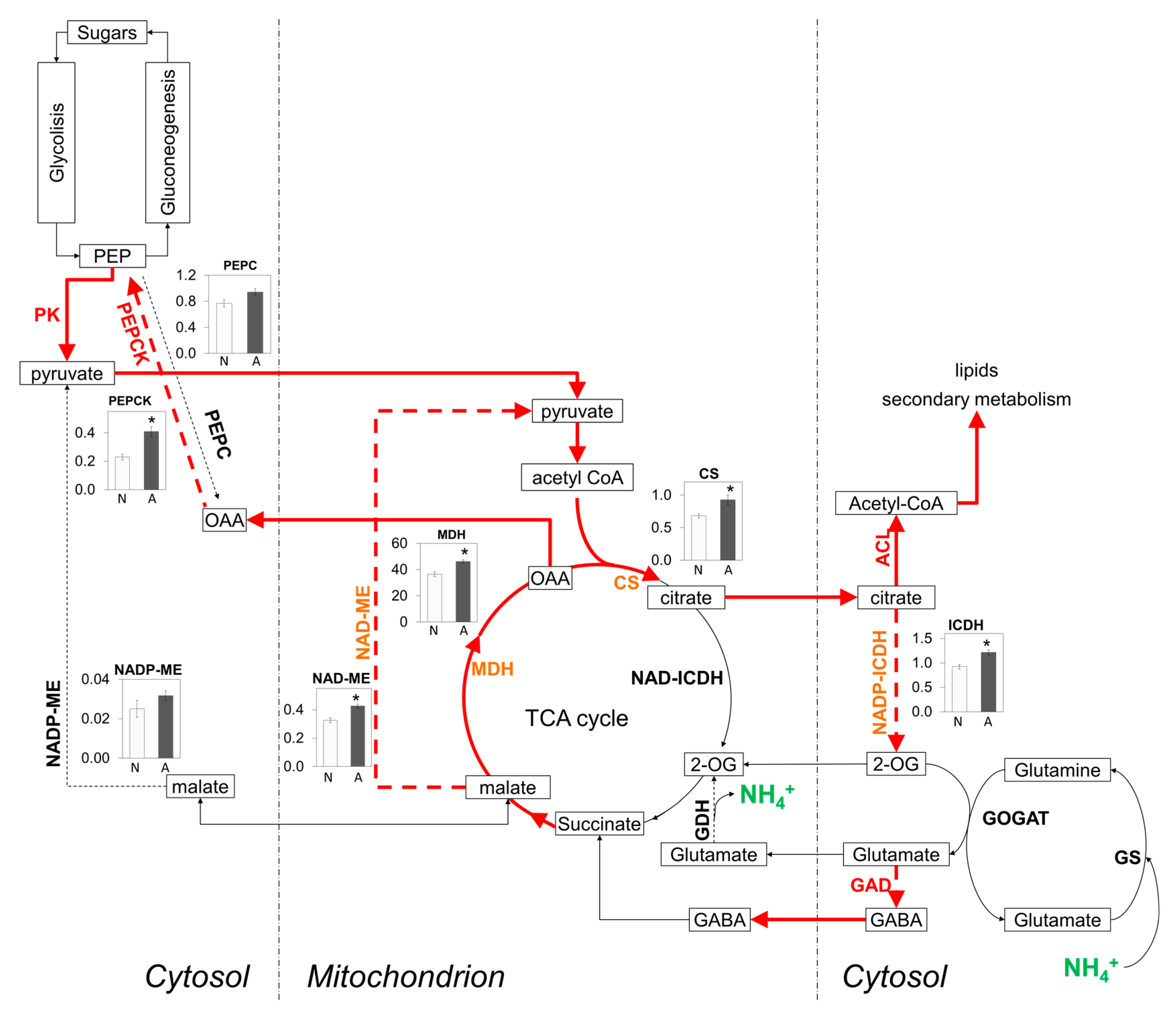
2.5. C/N Metabolism Modulation in Ammonium-Fed Plants May Be Driven by Alternative C Provision Routes to Tricarboxylic Acid (TCA) Cycle while Contributing to H+ Balance
3. Materials and Methods
3.1. Plant Culture and Experimental Design
3.2. Ammonium and Total Free Amino Acid Quantification
3.3. Soluble Protein Quantification and Enzyme Activities Determination
3.4. Glucosinolate Determination
3.5. Proteomic Analysis
3.5.1. Sample Preparation and Labeling for Proteomic Analysis
3.5.2. Peptide Fractionation
3.5.3. Triple-TOF 5600 Mass Spectrometry (MS) Analysis
3.5.4. Data Analysis
3.5.5. Functional Classification and Gene Ontology Enrichment Analysis
3.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-OG | 2-Oxoglutarate |
| AACT | Acetoacetyl-CoA thiolase |
| ACCT | Acetyl-CoA acetyltransferase |
| ACL | ATP-citrate lyase |
| AMT | Ammonium transporter |
| AOX | Alternative oxidase |
| ATP | Adenosine triphosphate |
| BR | Brassinosteroid |
| CHS | Chalcone synthase |
| CoA | Coenzyme A |
| CS | Citrate synthase |
| DMAP | Dimethylallyl phosphate |
| DMAPP | Dimethylallyl diphosphate |
| FDR | False discovery rate |
| FPP | Farnesyl diphosphate |
| FW | Fresh weight |
| GAD | Glutamate decarboxylase |
| GABA | Gamma-aminobutyric acid |
| GDH | Glutamate dehydrogenase |
| GLS | Glucosinolate(s) |
| GO | Gene ontology |
| GOGAT | Glutamate synthase |
| GS | Glutamine synthetase |
| ICDH | Isocitrate dehydrogenase |
| IP | Isopentenyl phosphate |
| IPP | Isopentenyl diphosphate |
| iTRAQ | Isobaric tags for relative and absolute quantitation |
| MDH | Malate dehydrogenase |
| ME | Malic enzyme |
| MVA | Mevalonate |
| MVAP | Phosphomevalonate |
| NAD(H) | Nicotinamide adenine dinucleotide (reduced) |
| NADP(H) | Nicotinamide adenine dinucleotide phosphate (reduced) |
| NUDIX 3 | Nudix hydrolase and a dipeptidyl peptidase III |
| OAA | Oxaloacetate |
| PEP | Phosphoenolpyruvate |
| PEPCK | Phosphoenolpyruvate carboxykinase |
| PEPC | Phosphoenolpyruvate carboxylase |
| PK | Pyruvate kinase |
| TCA | Tricarboxylic acid |
References
- Bloom, A.J. The increasing importance of distinguishing among plant nitrogen sources. Curr. Opin. Plant Biol. 2015, 25, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. Ecological significance and complexity of N-source preference in plants. Ann. Bot. 2013, 112, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Huérfano, X.; Fuertes-Mendizábal, T.; Duñabeitia, M.K.; González-Murua, C.; Estavillo, J.M.; Menéndez, S. Splitting the application of 3,4-dimethylpyrazole phosphate (DMPP): Influence on greenhouse gases emissions and wheat yield and quality under humid Mediterranean conditions. Eur. J. Agron. 2015, 64, 47–57. [Google Scholar] [CrossRef]
- Torralbo, F.; Menéndez, S.; Barrena, I.; Estavillo, J.M.; Marino, D.; González-Murua, C. Dimethyl pyrazol-based nitrification inhibitors effect on nitrifying and denitrifying bacteria to mitigate N2O emission. Sci. Rep. 2017, 7, 13810. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.; Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Bio, A.; Martins-Loução, M.A.; Moran, J.F. Intra-specific variation in pea responses to ammonium nutrition leads to different degrees of tolerance. Environ. Exp. Bot. 2011, 70, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Sarasketa, A.; González-Moro, M.B.; González-Murua, C.; Marino, D. Exploring ammonium tolerance in a large panel of Arabidopsis thaliana natural accessions. J. Exp. Bot. 2014, 65, 6023–6033. [Google Scholar] [CrossRef]
- Sarasketa, A.; González-Moro, M.B.; González-Murua, C.; Marino, D. Nitrogen source and external medium pH interaction differentially affects root and shoot metabolism in Arabidopsis. Front. Plant Sci. 2016, 7, 29. [Google Scholar] [CrossRef]
- Zhu, Y.; Lian, J.; Zeng, H.; Gan, L.; Di, T.; Shen, Q.; Xu, G. Involvement of plasma membrane H+-ATPase in adaption of rice to ammonium nutrient. Rice Sci. 2011, 18, 335–342. [Google Scholar] [CrossRef]
- Wang, F.; Gao, J.; Tian, Z.; Liu, Y.; Abid, M.; Jiang, D.; Cao, W.; Dai, T. Adaptation to rhizosphere acidification is a necessary prerequisite for wheat (Triticum aestivum L.) seedling resistance to ammonium stress. Plant Physiol. Biochem. 2016, 108, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Dynowski, M.; Ludewig, U. Ammonium ion transport by the AMT/Rh homologue LeAMT1;1. Biochem. J. 2006, 396, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants; Elsevier: London, UK, 2012; ISBN 9780124735422. [Google Scholar]
- Szczerba, M.W.; Britto, D.T.; Ali, S.A.; Balkos, K.D.; Kronzucker, H.J. NH4+-stimulated and -inhibited components of K+ transport in rice (Oryza sativa L.). J. Exp. Bot. 2008, 59, 3415–3423. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Ariz, I.; Lasa, B.; Santamaría, E.; Fernández-Irigoyen, J.; González-Murua, C.; Aparicio Tejo, P.M. Quantitative proteomics reveals the importance of nitrogen source to control glucosinolate metabolism in Arabidopsis thaliana and Brassica oleracea. J. Exp. Bot. 2016, 67. [Google Scholar] [CrossRef] [PubMed]
- Podgórska, A.; Burian, M.; Rychter, A.M.; Rasmusson, A.G.; Szal, B. Short-term ammonium supply induces cellular defence to prevent oxidative stress in Arabidopsis leaves. Physiol. Plant. 2017, 160, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Gerendás, J.; Abbadi, J.; Sattelmacher, B. Potassium efficiency of safflower (Carthamus tinctorius L.) and sunflower (Helianthus annuus L.). J. Plant Nutr. Soil Sci. 2008, 171, 431–439. [Google Scholar] [CrossRef]
- Walker, R.; Chen, Z.-H. Phosphoenolpyruvate carboxykinase: Structure, function and regulation. Adv. Bot. Res. 2002, 38, 93–189. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Hirai, Y. Good and bad protons: genetic aspects of acidity stress responses in plants. J. Exp. Bot. 2016, 67, 15–30. [Google Scholar] [CrossRef]
- Raven, J.A. Biochemical disposal of excess H+ in growing plants? New Phytol. 1986, 104, 175–206. [Google Scholar] [CrossRef]
- Fuertes-Mendizábal, T.; González-Torralba, J.; Arregui, L.M.; González-Murua, C.; González-Moro, M.B.; Estavillo, J.M. Ammonium as sole N source improves grain quality in wheat. J. Sci. Food Agric. 2013, 93, 2162–2171. [Google Scholar] [CrossRef]
- Coleto, I.; de la Peña, M.; Rodríguez-Escalante, J.; Bejarano, I.; Glauser, G.; Aparicio-Tejo, P.M.; González-Moro, M.B.; Marino, D. Leaves play a central role in the adaptation of nitrogen and sulfur metabolism to ammonium nutrition in oilseed rape (Brassica napus). BMC Plant Biol. 2017, 17, 157. [Google Scholar] [CrossRef]
- Gao, Y.; Li, Y.; Yang, X.; Li, H.; Shen, Q.; Guo, S. Ammonium nutrition increases water absorption in rice seedlings (Oryza sativa L.) under water stress. Plant Soil 2010, 331, 193–201. [Google Scholar] [CrossRef]
- Fernández-Crespo, E.; Camañes, G.; García-Agustín, P. Ammonium enhances resistance to salinity stress in citrus plants. J. Plant Physiol. 2012, 169, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Crespo, E.; Gómez-Pastor, R.; Scalschi, L.; Llorens, E.; Camañes, G.; García-Agustín, P. NH4+ induces antioxidant cellular machinery and provides resistance to salt stress in citrus plants. Trees 2014, 28, 1693–1704. [Google Scholar] [CrossRef]
- Hessini, K.; Hamed, K. Ben; Gandour, M.; Mejri, M.; Abdelly, C.; Cruz, C. Ammonium nutrition in the halophyte Spartina alterniflora under salt stress: evidence for a priming effect of ammonium? Plant Soil 2013, 370, 163–173. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Asensio, J.S.R.; Cousins, A.B. Carbon dioxide enrichment inhibits nitrate assimilation in wheat and arabidopsis. Science 2010, 328, 899–903. [Google Scholar] [CrossRef]
- Andrews, M.; Condron, L.M.; Kemp, P.D.; Topping, J.F.; Lindsey, K.; Hodge, S.; Raven, J.A. Elevated CO2 effects on nitrogen assimilation and growth of C3 vascular plants are similar regardless of N-form assimilated. J. Exp. Bot. 2019, 70, 683–690. [Google Scholar] [CrossRef]
- Vega-Mas, I.; Marino, D.; Sánchez-Zabala, J.; González-Murua, C.; Estavillo, J.M.; González-Moro, M.B. CO2 enrichment modulates ammonium nutrition in tomato adjusting carbon and nitrogen metabolism to stomatal conductance. Plant Sci. 2015, 241, 32–44. [Google Scholar] [CrossRef]
- Chaillou, S.; Vessey, J.K.; Morot-gaudry, J.F.; Raper, C.D.; Henry, L.T.; Boutin, J.P. Expression of characteristics of ammonium nutrition as affected by ph of the root medium. J. Exp. Bot. 1991, 42, 189–196. [Google Scholar] [CrossRef]
- Britto, D.T.; Siddiqi, M.Y.; Glass, A.D.M.; Kronzucker, H.J. Futile transmembrane NH4+ cycling: A cellular hypothesis to explain ammonium toxicity in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 4255–4258. [Google Scholar] [CrossRef]
- Cruz, C.; Bio, A.F.M.; Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Martins-Loução, M.A. How does glutamine synthetase activity determine plant tolerance to ammonium? Planta 2006, 223, 1068–1080. [Google Scholar] [CrossRef]
- Usadel, B.; Nagel, A.; Thimm, O.; Redestig, H.; Blaesing, O.E.; Palacios-Rojas, N.; Selbig, J.; Hannemann, J.; Piques, M.C.; Steinhauser, D.; et al. Extension of the visualization tool MapMan to allow statistical analysis of arrays, display of corresponding genes, and comparison with known responses. Plant Physiol. 2005, 138, 1195–1204. [Google Scholar] [CrossRef]
- Schubert, S.; Yan, F. Nitrate and ammonium nutrition of plants: Effects on acid/base balance and adaptation of root cell plasmalemma H+ ATPase. J. Plant Nutr. Soil Sci. 1997, 160, 275–281. [Google Scholar] [CrossRef]
- Anzi, C.; Pelucchi, P.; Vazzola, V.; Murgia, I.; Gomarasca, S.; Beretta Piccoli, M.; Morandini, P. The proton pump interactor (Ppi) gene family of Arabidopsis thaliana: expression pattern of Ppi1 and characterisation of knockout mutants for Ppi1 and 2. Plant Biol. 2008, 10, 237–249. [Google Scholar] [CrossRef]
- Vega-Mas, I.; Pérez-Delgado, C.M.; Marino, D.; Fuertes-Mendizábal, T.; González-Murua, C.; Márquez, A.J.; Betti, M.; Estavillo, J.M.; Gonzáez-Moro, M.B. Elevated CO2 induces root defensive mechanisms in tomato plants when dealing with ammonium toxicity. Plant Cell Physiol. 2017, 58, 2112–2125. [Google Scholar] [CrossRef]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct signalling pathways and transcriptome response signatures differentiate ammonium- and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef]
- Zeng, H.; Di, T.; Zhu, Y.; Subbarao, G.V. Transcriptional response of plasma membrane H+-ATPase genes to ammonium nutrition and its functional link to the release of biological nitrification inhibitors from sorghum roots. Plant Soil 2016, 398, 301–312. [Google Scholar] [CrossRef]
- Felle, H.H. pH Regulation in anoxic plants. Ann. Bot. 2005, 96, 519. [Google Scholar] [CrossRef]
- Katari, M.S.; Nowicki, S.D.; Aceituno, F.F.; Nero, D.; Kelfer, J.; Thompson, L.P.; Cabello, J.M.; Davidson, R.S.; Goldberg, A.P.; Shasha, D.E.; et al. VirtualPlant: A software platform to support systems biology research. Plant Physiol. 2010, 152, 500–515. [Google Scholar] [CrossRef]
- Sønderby, I.E.; Geu-Flores, F.; Halkier, B.A. Biosynthesis of glucosinolates—gene discovery and beyond. Trends Plant Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef]
- Wittstock, U.; Burow, M. Glucosinolate breakdown in Arabidopsis: mechanism, regulation and biological significance. Arab. B. 2010, 8, e0134. [Google Scholar] [CrossRef]
- Qasim, M.; Ashraf, M.; Ashraf, M.Y.; Rehman, S.-U.; Rha, E.S. Salt-induced changes in two canola cultivars differing in salt tolerance. Biol. Plant. 2003, 46, 629–632. [Google Scholar] [CrossRef]
- Zhang, H.; Schonhof, I.; Krumbein, A.; Gutezeit, B.; Li, L.; Stützel, H.; Schreiner, M. Water supply and growing season influence glucosinolate concentration and composition in turnip root (Brassica rapa ssp.rapifera L.). J. Plant Nutr. Soil Sci. 2008, 171, 255–265. [Google Scholar] [CrossRef]
- Schonhof, I.; Blankenburg, D.; Müller, S.; Krumbein, A. Sulfur and nitrogen supply influence growth, product appearance, and glucosinolate concentration of broccoli. J. Plant Nutr. Soil Sci. 2007, 170, 65–72. [Google Scholar] [CrossRef]
- Omirou, M.D.; Papadopoulou, K.K.; Papastylianou, I.; Constantinou, M.; Karpouzas, D.G.; Asimakopoulos, I.; Ehaliotis, C. Impact of nitrogen and sulfur fertilization on the composition of glucosinolates in relation to sulfur assimilation in different plant organs of Broccoli. J. Agric. Food Chem. 2009, 57, 9408–9417. [Google Scholar] [CrossRef]
- La, G.-X.; Yang, T.-G.; Fang, P.; Guo, H.-X.; Hao, X.; Huang, S.-M. Effect of NH4+/NO3− ratios on the growth and bolting stem glucosinolate content of Chinese kale (Brassica alboglabra L.H. Bailey). Aust. J. Crop Sci. 2013, 7, 618–624. [Google Scholar]
- Traka, M.H.; Mithen, R.F. Plant science and human nutrition: Challenges in assessing health-promoting properties of phytochemicals. Plant Cell 2011, 23, 2483–2497. [Google Scholar] [CrossRef]
- Menz, J.; Range, T.; Trini, J.; Ludewig, U.; Neuhäuser, B. Molecular basis of differential nitrogen use efficiencies and nitrogen source preferences in contrasting Arabidopsis accessions. Sci. Rep. 2018, 8, 3373. [Google Scholar] [CrossRef]
- Fatland, B.L.; Nikolau, B.J.; Wurtele, E.S. Reverse genetic characterization of cytosolic acetyl-CoA generation by ATP-citrate lyase in Arabidopsis. Plant Cell 2005, 17, 182–203. [Google Scholar] [CrossRef]
- Henry, L.K.; Thomas, S.T.; Widhalm, J.R.; Lynch, J.H.; Davis, T.C.; Kessler, S.A.; Bohlmann, J.; Noel, J.P.; Dudareva, N. Contribution of isopentenyl phosphate to plant terpenoid metabolism. Nat. Plants 2018, 4, 721–729. [Google Scholar] [CrossRef]
- Karačić, Z.; Vukelić, B.; Ho, G.H.; Jozić, I.; Sučec, I.; Salopek-Sondi, B.; Kozlović, M.; Brenner, S.E.; Ludwig-Müller, J.; Abramić, M. A novel plant enzyme with dual activity: an atypical Nudix hydrolase and a dipeptidyl peptidase III. Biol. Chem. 2017, 398, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.T.; Zhu, X.F.; Jung, J.H.; Xuan, Y.H. Effect of brassinosteroids on ammonium uptake via regulation of ammonium transporter and N-metabolism genes in Arabidopsis. Biol. Plant. 2016, 60, 563–571. [Google Scholar] [CrossRef]
- Xuan, Y.H.; Duan, F.Y.; Je, B. Il; Kim, C.M.; Li, T.Y.; Liu, J.M.; Park, S.J.; Cho, J.H.; Kim, T.H.; von Wiren, N.; et al. Related to ABI3/VP1-Like 1 ( RAVL1 ) regulates brassinosteroid-mediated activation of AMT1;2 in rice (Oryza sativa). J. Exp. Bot. 2016, 68, erw442. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Ariz, I.; Artola, E.; Asensio, A.C.; Cruchaga, S.; Aparicio-Tejo, P.M.; Moran, J.F. High irradiance increases NH4+ tolerance in Pisum sativum: Higher carbon and energy availability improve ion balance but not N assimilation. J. Plant Physiol. 2011, 168, 1009–1015. [Google Scholar] [CrossRef]
- Escobar, M.A.; Geisler, D.A.; Rasmusson, A.G. Reorganization of the alternative pathways of the Arabidopsis respiratory chain by nitrogen supply: Opposing effects of ammonium and nitrate. Plant J. 2006, 45, 775–788. [Google Scholar] [CrossRef]
- Hachiya, T.; Noguchi, K. Integrative response of plant mitochondrial electron transport chain to nitrogen source. Plant Cell Rep. 2011, 30, 195–204. [Google Scholar] [CrossRef]
- Frechilla, S.; Lasa, B.; Aleu, M.; Juanarena, N.; Lamsfus, C.; Aparicio-Tejo, P.M. Short-term ammonium supply stimulates glutamate dehydrogenase activity and alternative pathway respiration in roots of pea plants. J. Plant Physiol. 2002, 159, 811–818. [Google Scholar] [CrossRef]
- Sakano, K. Revision of biochemical pH-stat: Involvement of alternative pathway metabolisms. Plant Cell Physiol. 1998, 39, 467–473. [Google Scholar] [CrossRef]
- Sakano, K. Metabolic regulation of pH in plant cells: role of cytoplasmic pH in defense reaction and secondary metabolism. Int. Rev. Cytol. 2001, 206, 1–44. [Google Scholar]
- Hachiya, T.; Watanabe, C.K.; Boom, C.; Tholen, D.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Ammonium-dependent respiratory increase is dependent on the cytochrome pathway in Arabidopsis thaliana shoots. Plant Cell Environ. 2010, 33, 1888–1897. [Google Scholar] [CrossRef]
- Hagendoom, M.J.M.; Wagner, A.M.; Segers, C.; Van Der Plas, L.H.W.; Oostdam, A.; Van Walraven, H.S. Cytoplasmic acidification and secondary metabolite production in different plant cell suspensions (A comparative study). Plant Physiol. 1994, 106, 723–730. [Google Scholar] [CrossRef]
- Walker, R.P.; Chen, Z.; Johnson, K.E.; Famiani, F.; Tecsi, L.; Leegood, R.C. Using immunohistochemistry to study plant metabolism: the examples of its use in the localization of amino acids in plant tissues, and of phosphoenolpyruvate carboxykinase and its possible role in pH regulation. J. Exp. Bot. 2001, 52, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.P.; Benincasa, P.; Battistelli, A.; Moscatello, S.; Técsi, L.; Leegood, R.C.; Famiani, F. Gluconeogenesis and nitrogen metabolism in maize. Plant Physiol. Biochem. 2018, 130, 324–333. [Google Scholar] [CrossRef]
- Chen, Z.-H.; Walker, R.P.; Técsi, L.I.; Lea, P.J.; Leegood, R.C. Phosphoenolpyruvate carboxykinase in cucumber plants is increased both by ammonium and by acidification, and is present in the phloem. Planta 2004, 219, 48–58. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Beard, K.F.M.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not just a circle: flux modes in the plant TCA cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef]
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (GABA) and plant responses to stress. CRC. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Turano, F.J.; Fang, T.K. Characterization of two glutamate decarboxylase cDNA clones from Arabidopsis. Plant Physiol. 1998, 117, 1411–1421. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Xu, B.; Bose, J.; Kaur, S.; Conn, V.; Domingos, P.; Ullah, S.; Wege, S.; Shabala, S.; et al. GABA signalling modulates plant growth by directly regulating the activity of plant-specific anion transporters. Nat. Commun. 2015, 6, 7879. [Google Scholar] [CrossRef] [Green Version]
- Gilliham, M.; Tyerman, S.D. Linking Metabolism to Membrane Signaling: The GABA–Malate Connection. Trends Plant Sci. 2016, 21, 295–301. [Google Scholar] [CrossRef]
- Hachiya, T.; Watanabe, C.K.; Fujimoto, M.; Ishikawa, T.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Nitrate addition alleviates ammonium toxicity without lessening ammonium accumulation, organic acid depletion and inorganic cation depletion in Arabidopsis thaliana shoots. Plant Cell Physiol. 2012, 53, 577–591. [Google Scholar] [CrossRef]
- Solórzano, L. Determination of ammonia in natural waters by the phenolhypochlorite method. Limnol. Oceanogr. 1969, 14, 799–801. [Google Scholar] [CrossRef]
- Yemm, E.W.; Cocking, E.C.; Ricketts, R.E. The determination of amino-acids with ninhydrin. Analyst 1955, 80, 209. [Google Scholar] [CrossRef]
- Walker, R.P.; Chen, Z.-H.; Técsi, L.I.; Famiani, F.; Lea, P.J.; Leegood, R.C. Phosphoenolpyruvate carboxykinase plays a role in interactions of carbon and nitrogen metabolism during grape seed development. Planta 1999, 210, 9–18. [Google Scholar] [CrossRef]
- Srere, P.A. [1] Citrate synthase: [EC 4.1.3.7. Citrate oxaloacetate-lyase (CoA-acetylating)]. Methods Enzymol. 1969, 13, 3–11. [Google Scholar] [CrossRef]
- Glauser, G.; Schweizer, F.; Turlings, T.C.J.; Reymond, P. Rapid profiling of intact glucosinolates in Arabidopsis leaves by UHPLC-QTOFMS using a charged surface hybrid column. Phytochem. Anal. 2012, 23, 520–528. [Google Scholar] [CrossRef]
- Unwin, R.D.; Griffiths, J.R.; Whetton, A.D. Simultaneous analysis of relative protein expression levels across multiple samples using iTRAQ isobaric tags with 2D nano LC–MS/MS. Nat. Protoc. 2010, 5, 1574–1582. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Nitrate | Ammonium |
|---|---|---|
| Total plant biomass (mg·FW·plant−1) | 24.88 ± 1.48 | 24.41 ± 2.79 |
| Shoot biomass (mg·FW·plant−1) | 16.61 ± 0.69 | 14.35 ± 0.31 |
| Root biomass (mg·FW·plant−1) | 8.27 ± 0.82 | 10.06 ± 1.89 |
| Root NH4+ content (nmol·mg−1 FW) | 0.41 ± 0.03 | 0.57 ± 0.05 |
| Root total free amino acids (nmol·Gln·mg−1·FW) | 4.02 ± 1.5 | 11.57 ± 0.89 |
| Root total soluble protein (µg·mg−1·FW) | 4.1 ± 0.39 | 9.13 ± 1.19 |
| Protein Description | TAIR ID | Uniprot ID | p-Value | Fold Change NO3−/NH4+ |
|---|---|---|---|---|
| Organic acid transformation | ||||
| Dihydrolipoyllysine-residue acetyltransferase component 5 of pyruvate dehydrogenase complex | AT1G34430 | Q9C8P0 | 0.010 | 5.27 |
| Dihydrolipoyl dehydrogenase 1 | AT1G48030 | Q9M5K3 | 0.019 | 0.30 |
| ATP-citrate synthase alpha chain protein 3 | AT1G09430 | O80526 | 0.001 | 0.11 |
| Photorespiration | ||||
| Serine hydroxymethyltransferase 3, chloroplastic | AT4G32520 | Q94JQ3 | 0.011 | 0.19 |
| Carbohydrate metabolism | ||||
| ADP-glucose pyrophosphorylase family protein | AT1G74910 | F4HXD1 | 0.008 | 0.13 |
| Glycolysis | ||||
| Pyruvate kinase | AT5G56350 | Q9FM97 | 0.009 | 0.21 |
| Gluconeogenese/Glyoxylate cycle | ||||
| Phosphoenolpyruvate carboxykinase (ATP) | AT4G37870 | Q9T074 | 0.010 | 0.14 |
| Mitochondrial electron transport | ||||
| Gamma carbonic anhydrase-like 1 | AT5G63510 | F4KAG8 | 0.003 | 0.17 |
| Amino acid metabolism | ||||
| Glutamate decarboxylase 2 | AT1G65960 | Q42472 | 0.027 | 0.24 |
| Aspartate semialdehyde dehydrogenase | AT1G14810 | Q8VYI4 | 0.040 | 5.29 |
| Methylmalonate-semialdehyde dehydrogenase (acylating) | AT2G14170 | A8MQR6 | 0.048 | 4.36 |
| Secondary metabolism | ||||
| 3-isopropylmalate dehydratase large subunit 1 | AT4G13430 | Q94AR8 | 0.003 | 5.48 |
| Methylthioalkylmalate synthase 3 | AT5G23020 | Q9FN52 | 0.003 | 3.25 |
| Acetyl-CoA acetyltransferase | AT5G48230 | Q854Y1-2 | 0.049 | 0.30 |
| Betaine aldehyde dehydrogenase 1 | AT1G74920 | F4HXD2 | 0.037 | 0.26 |
| Chalcone synthase | AT5G13930 | P13114 | 0.039 | 0.26 |
| Hormone metabolism | ||||
| Delta(24)-sterol reductase | AT3G19820 | Q39085 | 0.037 | 0.31 |
| Co-factor and vitamin metabolism | ||||
| Nicotinate-nucleotide pyrophosphorylase (carboxylating) | AT2G01350 | F41NA0 | 0.015 | 0.10 |
| Cell wall synthesis | ||||
| UDP-glucuronic acid decarboxylase 3 | AT5G59290 | F4KHU8 | 0.029 | 2.93 |
| Tetrapyrrole synthesis | ||||
| Protoporphyrinogen oxidase 2 | AT5G14220 | Q8S9J1-2 | 0.003 | 0.19 |
| Abiotic stress | ||||
| Endoplasmin homolog | AT4G24190 | F4JQ55 | 0.031 | 3.16 |
| DnaJ protein ERDJ38 | AT3G62600 | Q9LZK5 | 0.002 | 6.67 |
| Probable methyltransferase PMt24 | AT1G29470 | Q6NPR7 | 0.004 | 5.51 |
| Germin-like protein subfamily T member 1 | AT1G18970 | P92995 | 0.031 | 3.55 |
| Redox response | ||||
| Protein disulfide isomerase-like 1-2 | AT1G21750 | F4HZN9 | 0.036 | 4.41 |
| Protein disulfide-isomerase | AT1G77510 | Q9SRG3 | 0.002 | 8.12 |
| Protein disulfide isomerase-like 1-6 | AT3G16110 | Q66GQ3 | 0.025 | 0.21 |
| Thioredoxin reductase 2 | AT2G17420 | Q39242 | 0.013 | 0.28 |
| Nucleotide metabolism | ||||
| Nudix hydrolase 3 | AT1G79690 | Q8L831 | 0.018 | 0.26 |
| RNA processing | ||||
| Polyadenylate-binding protein 2 | AT4G34110 | P42731 | 0.048 | 0.39 |
| Polyadenylate-binding protein 4 | AT2G23350 | O22173 | 0.006 | 0.18 |
| Reactive intermediate deaminase A | AT3G20390 | Q94JQ4 | 0.002 | 11.07 |
| Protein synthesis | ||||
| 30S ribosomal protein S3 | ATCG00800 | P56798 | 0.043 | 2.82 |
| 40S ribosomal protein S3-3 | AT5G35530 | Q9FJA6 | 0.004 | 7.52 |
| Elongation factor Tu (mitochondrial) | AT4G02930 | Q9ZT91 | 0.034 | 3.43 |
| Elongation factor Tu (chloroplastic) | AT4G20360 | P17745 | 0.026 | 5.32 |
| Protein degradation | ||||
| 26S proteasome non-ATPase regulatory subunit 14 homolog | AT5G23540 | Q9LT08 | 0.012 | 0.27 |
| Proteasome subunit beta type-4 | AT1G56450 | Q7DLR9 | 0.035 | 0.27 |
| Protein targeting | ||||
| Nuclear pore complex protein NUP155 | AT1G14850 | F4HXV6 | 0.033 | 2.99 |
| ADP-ribosylation factor 2-A | AT3G62290 | Q9M1P5 | 0.031 | 4.87 |
| Signaling | ||||
| Rho GDP-dissociation inhibitor 1 | AT3G07880 | Q9SFC6 | 0.021 | 5.59 |
| Dynamin-related protein 1A | AT5G42080 | P42697 | 0.003 | 5.79 |
| GTP-binding nuclear protein Ran-1 | AT5G20010 | P41916 | 0.020 | 4.92 |
| 14-3-3-like protein GF14 chi | AT4G09000 | P42643 | 0.032 | 0.22 |
| Cell vesicle transport | ||||
| Tubulin beta 6-chain (Cell organization) | AT5G12250 | P29514 | 0.020 | 0.28 |
| AP-4 complex subunit epsilon | AT1G31730 | Q8L7A9 | 0.030 | 3.63 |
| Coatomer subunit beta-1 | AT4G31480 | Q95V21 | 0.007 | 8.98 |
| Golgin candidate 5 | AT1G79830 | F4HQB9 | 0.015 | 5.49 |
| COG complex component-related protein | AT5G51430 | Q9FGN0 | 0.023 | 6.23 |
| Transport | ||||
| V-type proton ATPase subunit E3 | AT1G64200 | P0CAN7 | 0.044 | 2.89 |
| ATPase 2, plasma membrane-type | AT4G30190 | P19456 | 0.046 | 3.26 |
| Mitochondrial dicarboxylate/tricarboxylate transporter DTC | AT5G19760 | Q9C5M0 | 0.010 | 3.54 |
| ABC transporter A family member 2 | AT3G47730 | Q84K47 | 0.010 | 3.82 |
| ABC transporter F family member 1 | AT5G60790 | Q9FJH5 | 0.009 | 0.15 |
| Miscellaneous | ||||
| AT3g23600/MDB19_9 | ATG23600 | Q9LUG8 | 0.012 | 4.83 |
| Dolichyl-diphosphooligosaccharide-protein glycosyltransferase 48 kDa subunit | AT5G66680 | Q944K2 | 0.009 | 0.17 |
| Methylesterase 3 | AT2G23610 | O80477 | 0.0369 | 2.93 |
| Glutathione S-transferase L3 | AT5G02790 | Q9LZO6 | 0.0218 | 3.61 |
| Peroxidase 34 | AT3G49120 | Q9SMU8 | 0.0070 | 0.16 |
| Peroxidase 30 | AT3G21770 | Q9LSY7 | 0.0165 | 4.46 |
| AT4g13180/F17N18-70 | AT4G13180 | Q9SVQ9 | 0.0322 | 0.22 |
| Not assigned ontology | ||||
| NADH dehydrogenase (ubiquinone) iron-sulfur protein 2 | ATMG00510 | P93306 | 0.0167 | 1.78 |
| tRNA (guanine-N(7)-)-methyltransferase non-catalytic subunit | AT1G03110 | Q93WD7 | 0.0376 | 0.31 |
| Pheromone receptor, putative (AR401) | AT1G66680 | Q9C9M1 | 0.0143 | 4.38 |
| WD40 domain-containing protein | AT5G24710 | F4K1H8 | 0.0265 | 0.29 |
| Protein EMBRYO DEFECTIVE 2734 | AT5G19820 | Q93V68 | 0.0208 | 0.20 |
| Calcium-dependent lipid-binding family protein | AT1G48090 | F4HWS2 | 0.0156 | 0.29 |
| Metal-dependent protein hydrolase | AT5G41970 | F4K000 | 0.0349 | 3.09 |
| Aliphatic Glucosinolates | Nitrate | Ammonium |
|---|---|---|
| Glucoraphanin (4MSOB) | 25.61 ± 4.13 | 64.12 ± 8.76 |
| Glucoalyssin (5MSOP) | 3.59 ± 0.32 | 6.04 ± 0.52 |
| Glucoiberin (3MSOP) | 1.88 ± 0.27 | 4.50 ± 0.55 |
| Glucoerucin (4MTB) | 2.20 ± 0.52 | 4.25 ± 0.38 |
| Glucoberteroin (5MTP) | 0.87 ± 0.08 | 1.50 ± 0.08 |
| Glucoibarin (7MSOH) | 54.17 ± 4.76 | 44.56 ± 3.01 |
| Glucohirsutin (8MSOO) | 610.00 ± 57.62 | 452.58 ± 33.60 |
| C6-aliphatic GLS A (C13H24NO9S2) | 0.57 ± 0.07 | 0.80 ± 0.10 |
| 7-Methylthioheptyl-GS (C15H28NO9S3) | 125.12 ± 11.28 | 92.59 ± 4.57 |
| 8-Methylthiooctyl-GS (C16H31NO9S3) | 1090.72 ± 58.47 | 786.08 ± 33.54 |
| Total Aliphatic | 1914.67 ± 125.02 | 1457.04 ± 69.85 |
| Indolic Glucosinolates | Nitrate | Ammonium |
| Glucobrassicin (I3M) | 96.97 ± 8.25 | 97.54 ± 4.64 |
| Neoglucobrassicin (IMOI3M) | 268.83 ± 25.97 | 214.38 ± 16.71 |
| Hydroxyglucobrassicin (4OHI3M) | 12.39 ± 1.00 | 11.73 ± 0.37 |
| Methoxyglucobrassicin (4MOI3M) | 21.90 ± 2.62 | 20.87 ± 2.58 |
| Total Indolic | 400.10 ± 36.42 | 344.52 ± 22.24 |
| Total Glucosinolates | 2337.88 ± 158.74 | 1818.12 ± 90.34 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coleto, I.; Vega-Mas, I.; Glauser, G.; González-Moro, M.B.; Marino, D.; Ariz, I. New Insights on Arabidopsis thaliana Root Adaption to Ammonium Nutrition by the Use of a Quantitative Proteomic Approach. Int. J. Mol. Sci. 2019, 20, 814. https://doi.org/10.3390/ijms20040814
Coleto I, Vega-Mas I, Glauser G, González-Moro MB, Marino D, Ariz I. New Insights on Arabidopsis thaliana Root Adaption to Ammonium Nutrition by the Use of a Quantitative Proteomic Approach. International Journal of Molecular Sciences. 2019; 20(4):814. https://doi.org/10.3390/ijms20040814
Chicago/Turabian StyleColeto, Inmaculada, Izargi Vega-Mas, Gaetan Glauser, María Begoña González-Moro, Daniel Marino, and Idoia Ariz. 2019. "New Insights on Arabidopsis thaliana Root Adaption to Ammonium Nutrition by the Use of a Quantitative Proteomic Approach" International Journal of Molecular Sciences 20, no. 4: 814. https://doi.org/10.3390/ijms20040814