Protein S is Protective in Acute Lung Injury by Inhibiting Cell Apoptosis

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
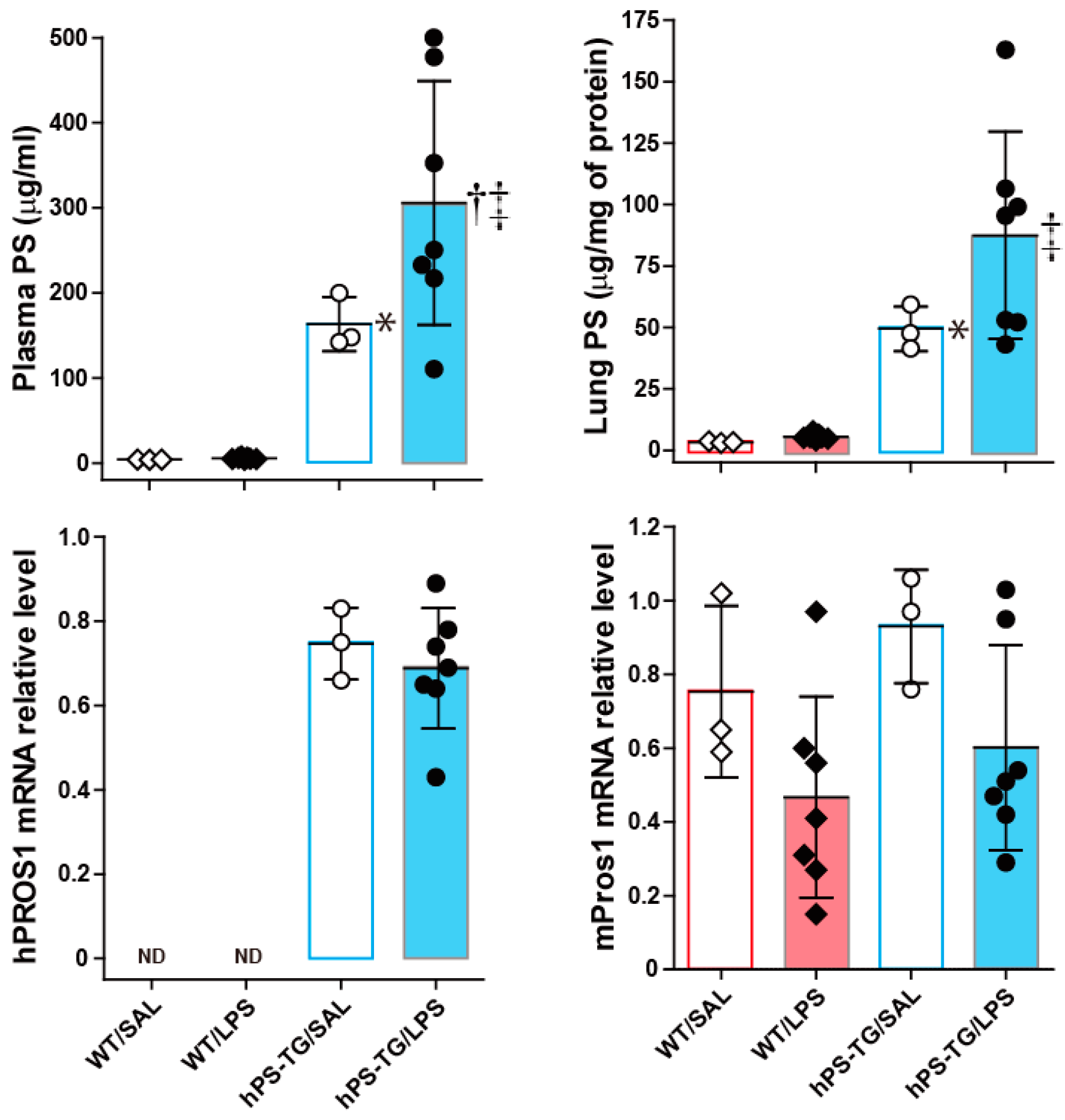
2.1. Systemic and Lung Upregulation of Protein S (PS) During Lipopolysaccharide (LPS)-Induced ALI
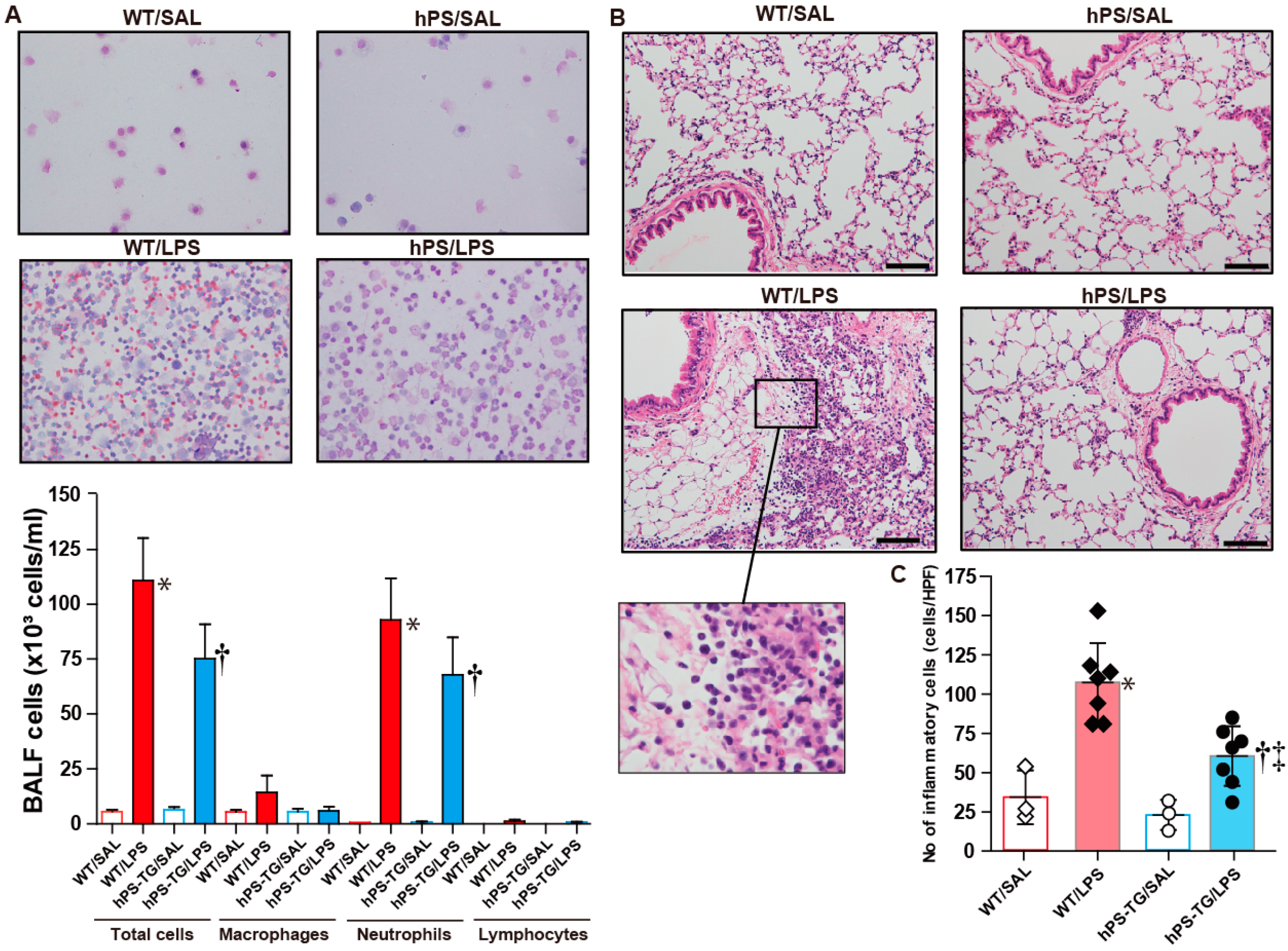
2.2. Less Lung Cell Infiltration in Mice Overexpressing Human Protein S (hPS)
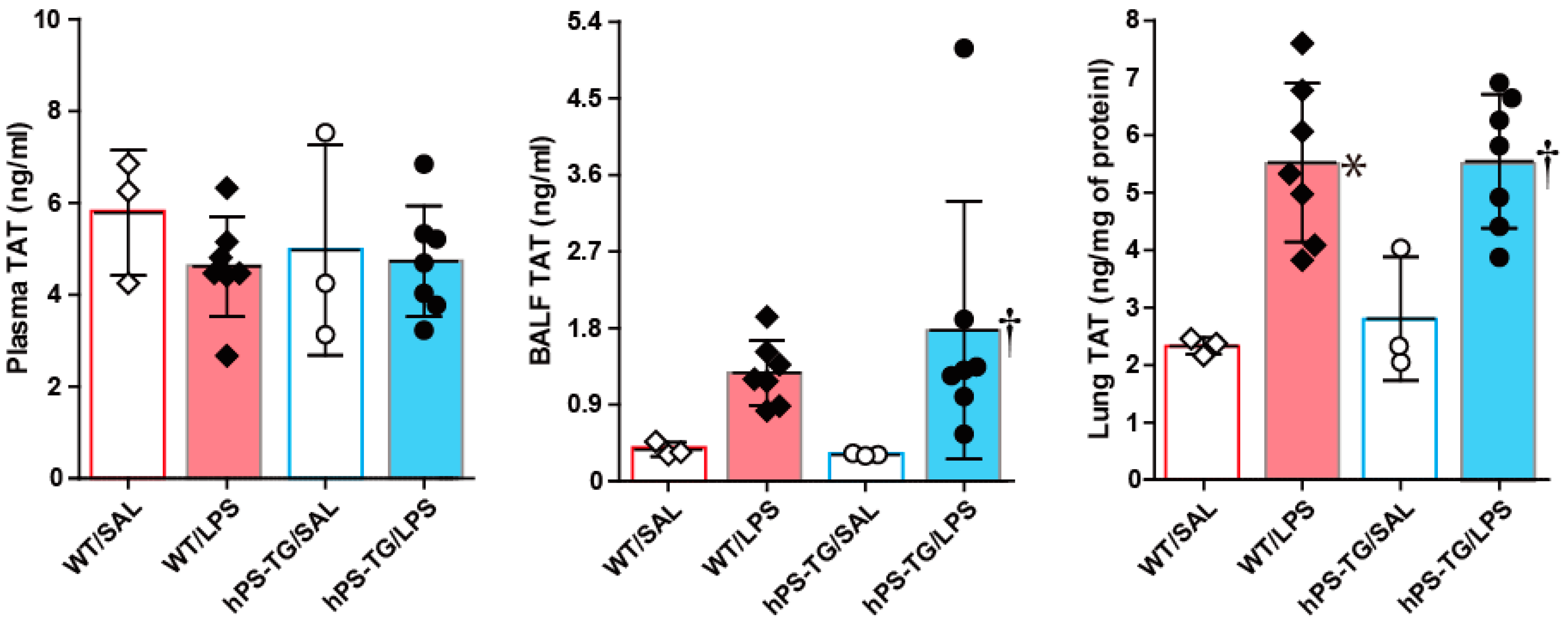
2.3. The Coagulation System Was Not Affected by hPS Overexpression
2.4. Suppression of Pro-Inflammatory Markers by hPS Overexpression
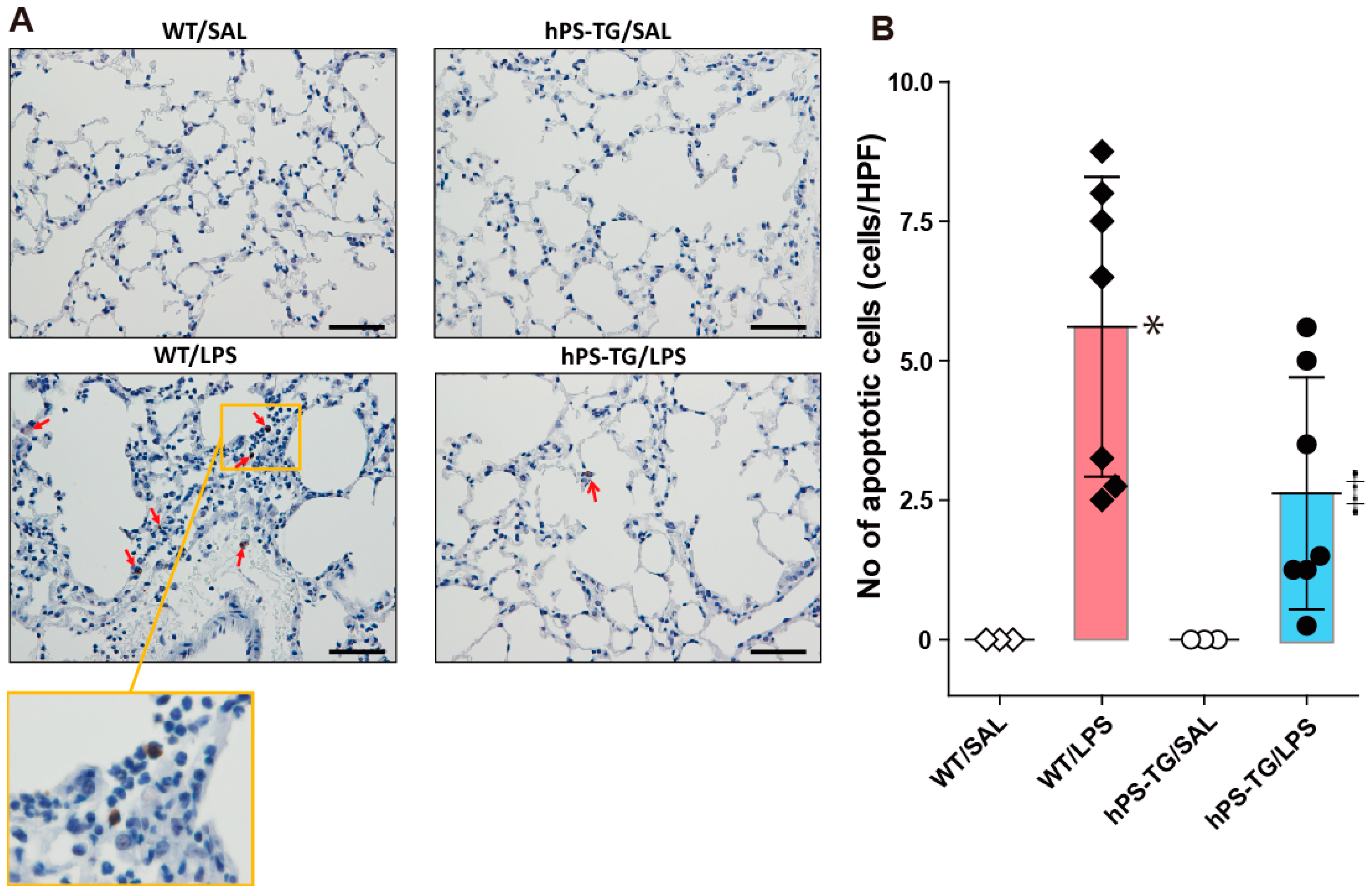
2.5. Decreased Lung Apoptotic Cells in Mice Overexpressing hPS
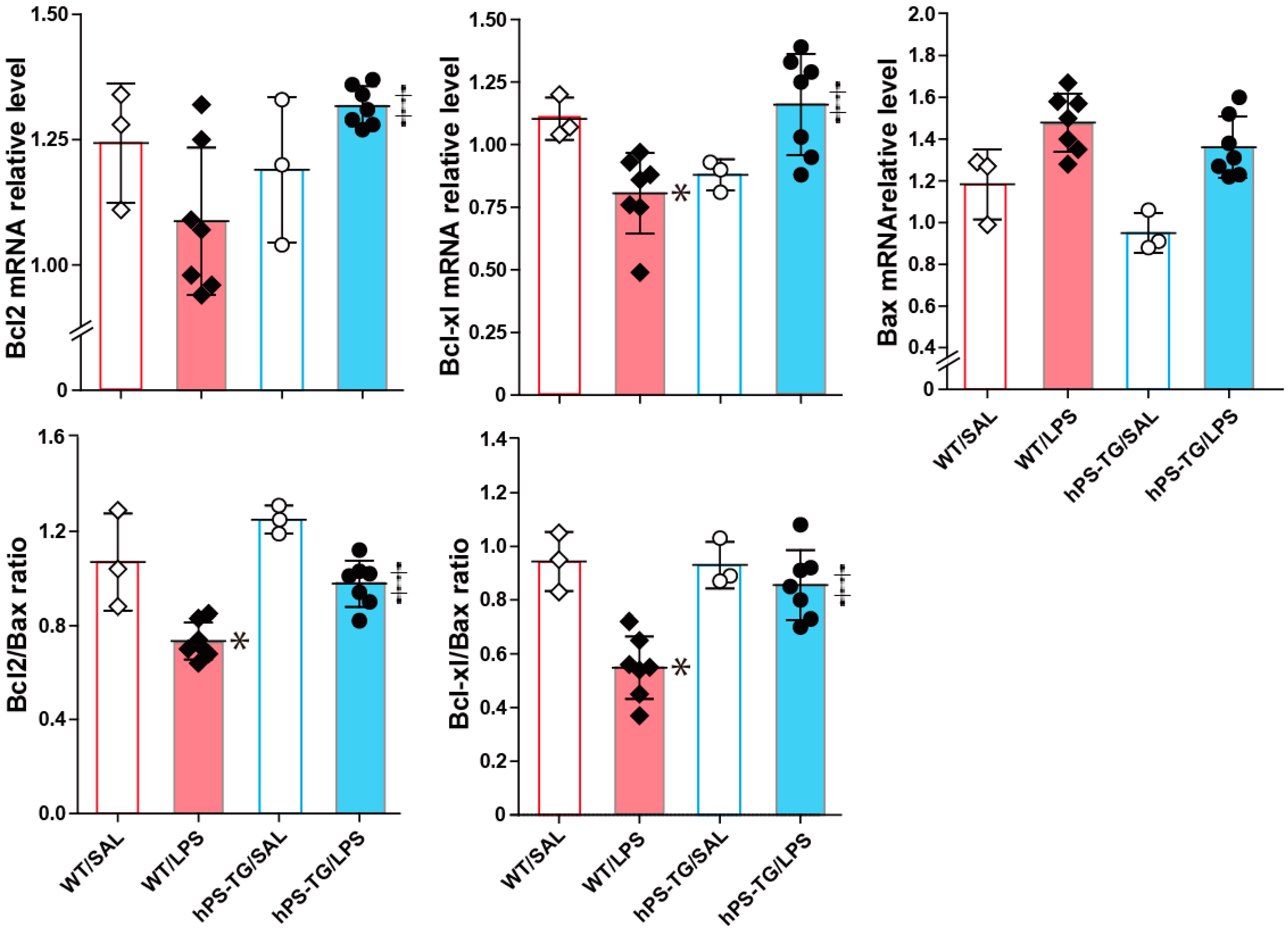
2.6. The Expression of Apoptotic Factors Is Regulated by hPS
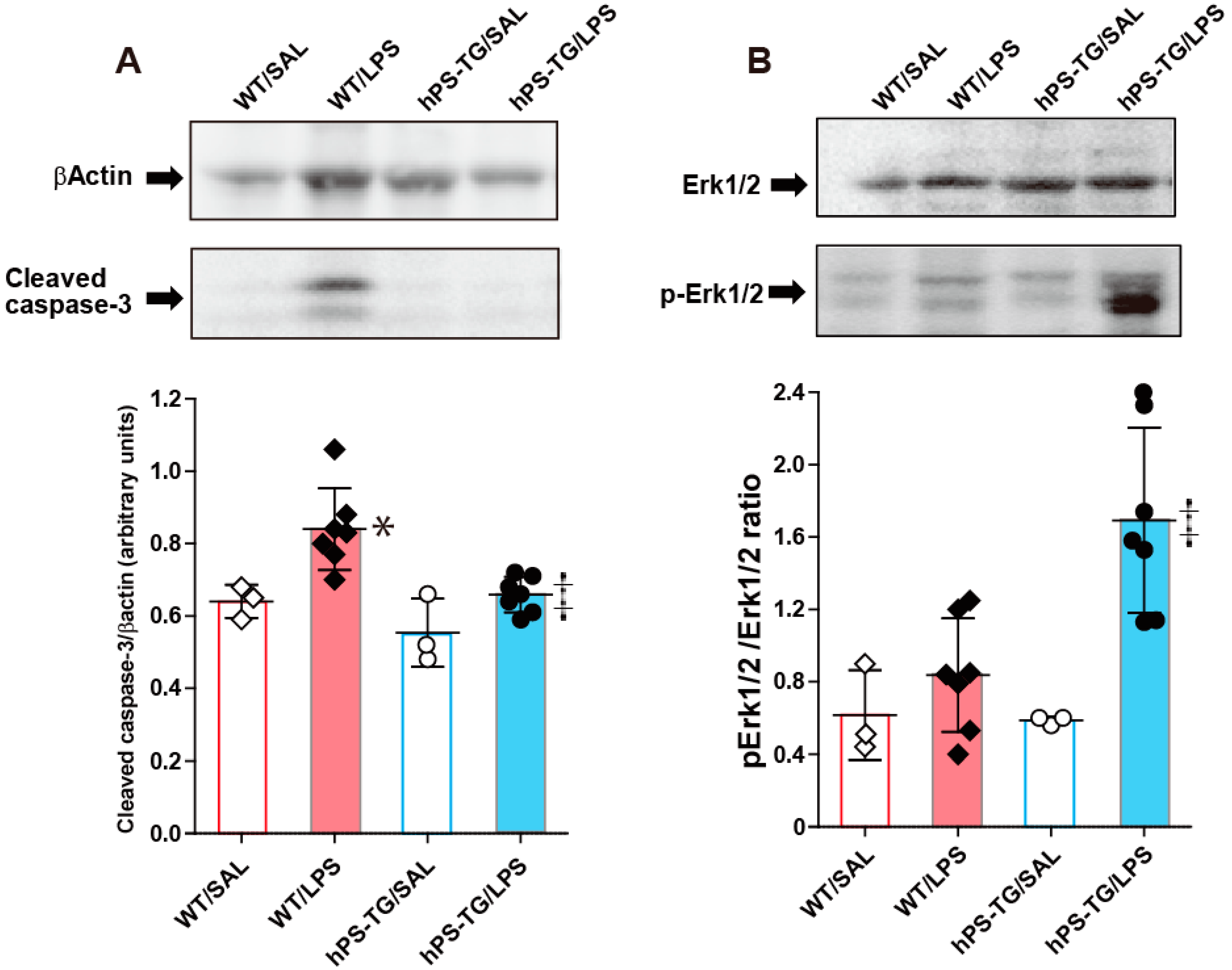
2.7. Decreased Activation of Caspase-3 in Mice Overexpressing hPS
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Acute Lung Injury (ALI) Induction
4.3. Collection of Samples
4.4. Histological Examination
4.5. Western Blotting
4.6. Immunohistochemistry
4.7. Biochemical Analysis
4.8. Gene Expression Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fan, E.; Brodie, D.; Slutsky, A.S. Acute Respiratory Distress Syndrome: Advances in Diagnosis and Treatment. JAMA 2018, 319, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Umbrello, M.; Formenti, P.; Bolgiaghi, L.; Chiumello, D. Current Concepts of ARDS: A Narrative Review. Int. J. Mol. Sci. 2016, 18, 64. [Google Scholar] [CrossRef] [PubMed]
- Laffey, J.G.; Madotto, F.; Bellani, G.; Pham, T.; Fan, E.; Brochard, L.; Amin, P.; Arabi, Y.; Bajwa, E.K.; Bruhn, A.; et al. Geo-economic variations in epidemiology, patterns of care, and outcomes in patients with acute respiratory distress syndrome: Insights from the LUNG SAFE prospective cohort study. Lancet Respir. Med. 2017, 5, 627–638. [Google Scholar] [CrossRef]
- Mutlu, G.M.; Budinger, G.R. Incidence and outcomes of acute lung injury. N. Engl. J. Med. 2006, 354, 416–417; author reply 416–417. [Google Scholar] [PubMed]
- Bellani, G.; Laffey, J.G.; Pham, T.; Fan, E.; Brochard, L.; Esteban, A.; Gattinoni, L.; van Haren, F.; Larsson, A.; McAuley, D.F.; et al. Epidemiology, Patterns of Care, and Mortality for Patients With Acute Respiratory Distress Syndrome in Intensive Care Units in 50 Countries. JAMA 2016, 315, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Dias-Freitas, F.; Metelo-Coimbra, C.; Roncon-Albuquerque, R., Jr. Molecular mechanisms underlying hyperoxia acute lung injury. Respir. Med. 2016, 119, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Choi, Y.H.; Kim, Y.S.; Baik, S.H.; Oh, Y.J.; Sheen, S.S.; Park, J.H.; Hwang, S.C.; Park, K.J. Evaluation of bronchoalveolar lavage fluid from ARDS patients with regard to apoptosis. Respir. Med. 2008, 102, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Q.; Sun, Z.; Yang, Z.; Zhang, W.; Ren, Y.; Chen, W.; Yao, M.; Nie, S. Protective effect of ginsenoside Rg1 on LPS-induced apoptosis of lung epithelial cells. Mol. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Gao, F.; Hao, J.; Liu, Z. microRNA-1246 mediates lipopolysaccharide-induced pulmonary endothelial cell apoptosis and acute lung injury by targeting angiotensin-converting enzyme 2. Am. J. Transl. Res. 2017, 9, 1287–1296. [Google Scholar] [PubMed]
- Lin, W.C.; Chen, C.W.; Huang, Y.W.; Chao, L.; Chao, J.; Lin, Y.S.; Lin, C.F. Kallistatin protects against sepsis-related acute lung injury via inhibiting inflammation and apoptosis. Sci. Rep. 2015, 5, 12463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albertine, K.H.; Soulier, M.F.; Wang, Z.; Ishizaka, A.; Hashimoto, S.; Zimmerman, G.A.; Matthay, M.A.; Ware, L.B. Fas and fas ligand are up-regulated in pulmonary edema fluid and lung tissue of patients with acute lung injury and the acute respiratory distress syndrome. Am. J. Pathol. 2002, 161, 1783–1796. [Google Scholar] [CrossRef]
- Kawasaki, M.; Kuwano, K.; Hagimoto, N.; Matsuba, T.; Kunitake, R.; Tanaka, T.; Maeyama, T.; Hara, N. Protection from lethal apoptosis in lipopolysaccharide-induced acute lung injury in mice by a caspase inhibitor. Am. J. Pathol. 2000, 157, 597–603. [Google Scholar] [CrossRef]
- Ma, X.; Xu, D.; Ai, Y.; Ming, G.; Zhao, S. Fas inhibition attenuates lipopolysaccharide-induced apoptosis and cytokine release of rat type II alveolar epithelial cells. Mol. Biol. Rep. 2010, 37, 3051–3056. [Google Scholar] [CrossRef] [PubMed]
- Dahlback, B. Vitamin K-Dependent Protein S: Beyond the Protein C Pathway. Semin. Thromb. Hemost. 2018, 44, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, J.H.; van der Poll, T.; van’t Veer, C. TAM receptors, Gas6, and protein S: Roles in inflammation and hemostasis. Blood 2014, 123, 2460–2469. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Taguchi, O.; Aoki, S.; Toda, M.; Yamaguchi, A.; Fujimoto, H.; Boveda-Ruiz, D.; Gil-Bernabe, P.; Ramirez, A.Y.; Naito, M.; et al. Direct effects of protein S in ameliorating acute lung injury. J. Thromb. Haemost. 2009, 7, 2053–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urawa, M.; Kobayashi, T.; D’Alessandro-Gabazza, C.N.; Fujimoto, H.; Toda, M.; Roeen, Z.; Hinneh, J.A.; Yasuma, T.; Takei, Y.; Taguchi, O.; et al. Protein S is protective in pulmonary fibrosis. J. Thromb. Haemost. 2016, 14, 1588–1599. [Google Scholar] [CrossRef] [PubMed]
- Yasuma, T.; Yano, Y.; D’Alessandro-Gabazza, C.N.; Toda, M.; Gil-Bernabe, P.; Kobayashi, T.; Nishihama, K.; Hinneh, J.A.; Mifuji-Moroka, R.; Roeen, Z.; et al. Amelioration of Diabetes by Protein S. Diabetes 2016, 65, 1940–1951. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot-Govindalayathil, A.L.; Mifuji-Moroka, R.; D’Alessandro-Gabazza, C.N.; Toda, M.; Matsuda, Y.; Gil-Bernabe, P.; Roeen, Z.; Yasuma, T.; Yano, Y.; Gabazza, E.C.; et al. Protein S exacerbates alcoholic hepatitis by stimulating liver natural killer T cells. J. Thromb. Haemost. 2015, 13, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Totoki, T.; D’Alessandro-Gabazza, C.N.; Toda, M.; Tonto, P.B.; Takeshita, A.; Yasuma, T.; Nishihama, K.; Iwasa, M.; Horiki, N.; Takei, Y.; et al. Protein S Exacerbates Chronic Liver Injury and Fibrosis. Am. J. Pathol. 2018, 188, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Barlos, D.; Deitch, E.A.; Watkins, A.C.; Caputo, F.J.; Lu, Q.; Abungu, B.; Colorado, I.; Xu, D.Z.; Feinman, R. Trauma-hemorrhagic shock-induced pulmonary epithelial and endothelial cell injury utilizes different programmed cell death signaling pathways. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L404–L417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, M.; Lomas-Neira, J.; Chung, C.S.; Ayala, A. Epithelial cell apoptosis and neutrophil recruitment in acute lung injury—A unifying hypothesis? What we have learned from small interfering RNAs. Mol. Med. 2008, 14, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Dai, Q.; Huang, X. Neutrophils in acute lung injury. Front. Biosci. 2012, 17, 2278–2283. [Google Scholar] [CrossRef]
- Robb, C.T.; Regan, K.H.; Dorward, D.A.; Rossi, A.G. Key mechanisms governing resolution of lung inflammation. Semin. Immunopathol. 2016, 38, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.E.; Krodel, M.; Pazos, M.; Lai, C.; Prieto, A.L. Cross-phosphorylation, signaling and proliferative functions of the Tyro3 and Axl receptors in Rat2 cells. PLoS ONE 2012, 7, e36800. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.J.; Stuart, K.; Gilley, R.; Sale, M.J. Control of cell death and mitochondrial fission by ERK1/2 MAP kinase signalling. FEBS J. 2017, 284, 4177–4195. [Google Scholar] [CrossRef] [PubMed]
- Fraineau, S.; Monvoisin, A.; Clarhaut, J.; Talbot, J.; Simonneau, C.; Kanthou, C.; Kanse, S.M.; Philippe, M.; Benzakour, O. The vitamin K-dependent anticoagulant factor, protein S, inhibits multiple VEGF-A-induced angiogenesis events in a Mer- and SHP2-dependent manner. Blood 2012, 120, 5073–5083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Zhang, Z.; Miao, L.; Yang, Z.; Yang, J.; Wang, Y.; Qian, D.; Cai, H.; Wang, Y. Anexelekto (AXL) Increases Resistance to EGFR-TKI and Activation of AKT and ERK1/2 in Non-Small Cell Lung Cancer Cells. Oncol. Res. 2016, 24, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, L.; Négrier, C.; Boukerche, H. Protein S: A multifunctional anticoagulant vitamin K-dependent protein at the crossroads of coagulation, inflammation, angiogenesis, and cancer. Crit. Rev. Oncol. Hematol. 2013, 88, 637–654. [Google Scholar] [CrossRef] [PubMed]
- Kanji, G.K.; SAGE Research Methods Core. 100 Statistical Tests, 3rd ed.; SAGE Publications, Incorporated Distributor: London, UK; Thousand Oaks, CA, USA, 2006; 256p. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baffour Tonto, P.; Yasuma, T.; Kobayashi, T.; D’Alessandro-Gabazza, C.N.; Toda, M.; Saiki, H.; Fujimoto, H.; Asayama, K.; Fujiwara, K.; Nishihama, K.; et al. Protein S is Protective in Acute Lung Injury by Inhibiting Cell Apoptosis. Int. J. Mol. Sci. 2019, 20, 1082. https://doi.org/10.3390/ijms20051082
Baffour Tonto P, Yasuma T, Kobayashi T, D’Alessandro-Gabazza CN, Toda M, Saiki H, Fujimoto H, Asayama K, Fujiwara K, Nishihama K, et al. Protein S is Protective in Acute Lung Injury by Inhibiting Cell Apoptosis. International Journal of Molecular Sciences. 2019; 20(5):1082. https://doi.org/10.3390/ijms20051082
Chicago/Turabian StyleBaffour Tonto, Prince, Taro Yasuma, Tetsu Kobayashi, Corina N. D’Alessandro-Gabazza, Masaaki Toda, Haruko Saiki, Hajime Fujimoto, Kentaro Asayama, Kentaro Fujiwara, Kota Nishihama, and et al. 2019. "Protein S is Protective in Acute Lung Injury by Inhibiting Cell Apoptosis" International Journal of Molecular Sciences 20, no. 5: 1082. https://doi.org/10.3390/ijms20051082