Human Brain Injury and miRNAs: An Experimental Study

 ,
,  , , ,
, , , 

Abstract
:1. Introduction
- -
- stroke group (SG): 5 men who died from brain stroke;
- -
- drug group (DG): 5 men who died from drug abuse (cocaine);
- -
- aged group (AG): 5 older men who died from Sudden Cardiac arrest.
2. Results
miRNA Quantitative Real-Time PCR (qRT-PCR)
3. Discussion
4. Material and Methods
4.1. Selected Cases
4.2. miRNA Quantitative Real-Time PCR (qRT-PCR)
4.3. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sessa, F.; Anna, V.; Messina, G.; Cibelli, G.; Monda, V.; Marsala, G.; Ruberto, M.; Biondi, A.; Cascio, O.; Bertozzi, G.; et al. Heart rate variability as predictive factor for sudden cardiac death. Aging Albany NY 2018, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Sessa, F.; Messina, G.; Valenzano, A.; Messina, A.; Salerno, M.; Marsala, G.; Bertozzi, G.; Daniele, A.; Monda, V.; Russo, R. Sports training and adaptive changes. Sport Sci. Health 2018, 14, 705–708. [Google Scholar] [CrossRef]
- Bertozzi, G.; Sessa, F.; Albano, G.D.; Sani, G.; Maglietta, F.; Roshan, M.H.K.; Volti, G.L.; Bernardini, R.; Avola, R.; Pomara, C.; et al. The role of anabolic androgenic steroids in disruption of the physiological function in discrete areas of the central nervous system. Mol. Neurobiol. 2018, 55, 5548–5556. [Google Scholar] [CrossRef]
- Albano, G.D.; Sessa, F.; Messina, A.; Monda, V.; Bertozzi, G.; Maglietta, F.; Giugliano, P.; Vacchiano, G.; Gabriella, M.; Salerno, M. AAS and organs damage: A focus on Nandrolone effects. Acta Medica Mediterr. 2017, 33, 939–946. [Google Scholar]
- Monda, V.; Salerno, M.; Sessa, F.; Bernardini, R.; Valenzano, A.; Marsala, G.; Zammit, C.; Avola, R.; Carotenuto, M.; Messina, G.; et al. Functional changes of orexinergic reaction to psychoactive substances. Mol. Neurobiol. 2018, 55, 6362–6368. [Google Scholar] [CrossRef]
- Messina, A.; Monda, V.; Sessa, F.; Valenzano, A.; Salerno, M.; Bitetti, I.; Precenzano, F.; Marotta, R.; Lavano, F.; Lavano, S.M.; et al. Sympathetic, metabolic adaptations, and oxidative stress in autism spectrum disorders: How far from physiology? Front. Physiol. 2018, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Sessa, F.; Salerno, M.; Di Mizio, G.; Bertozzi, G.; Messina, G.; Tomaiuolo, B.; Pisanelli, D.; Maglietta, F.; Ricci, P.; Pomara, C. Anabolic androgenic steroids: Searching new molecular biomarkers. Front. Pharmacol. 2018, 9, 1321. [Google Scholar] [CrossRef]
- Bafunno, V.; Santacroce, R.; Chetta, M.; Peyvandi, F.; Sessa, F.; Chinni, E.; Longo, V.; Margaglione, M. Polymorphic miRNA-mediated gene contribution to inhibitor development in haemophilia A. Haemophilia 2012, 18, 1003–1007. [Google Scholar] [CrossRef]
- Lecellier, C.H.; Dunoyer, P.; Arar, K.; Lehmann-Che, J.; Eyquem, S.; Himber, C.; Saïb, A.; Voinnet, O. A cellular microRNA mediates antiviral defense in human cells. Science 2005, 308, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.L.; Chen, H.Y.; Chang, G.C.; Chen, C.Y.; Chen, H.W.; Singh, S.; Cheng, C.L.; Yu, C.J.; Lee, Y.C.; Chen, H.S.; et al. MicroRNA signature predicts survival and relapse in lung cancer. Cancer Cell 2008, 13, 48–57. [Google Scholar] [CrossRef]
- Segura, M.F.; Belitskaya-Lévy, I.; Rose, A.E.; Zakrzewski, J.; Gaziel, A.; Hanniford, D.; Darvishian, F.; Berman, R.S.; Shapiro, R.L.; Pavlick, A.C.; et al. Melanoma microRNA signature predicts post-recurrence survival. Clin. Cancer Res. 2010, 16, 1577–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salerno, M.; Cascio, O.; Bertozzi, G.; Sessa, F.; Messina, A.; Monda, V.; Cipolloni, L.; Biondi, A.; Daniele, A.; Pomara, C. Anabolic androgenic steroids and carcinogenicity focusing on Leydig cell: A literature review. Oncotarget 2018, 9, 19415. [Google Scholar] [CrossRef]
- Van Rooij, E.; Sutherland, L.B.; Liu, N.; Williams, A.H.; McAnally, J.; Gerard, R.D.; Richardson, J.A.; Olson, E.N. A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure. Proc. Natl. Acad. Sci. USA 2006, 103, 18255–18260. [Google Scholar] [CrossRef] [Green Version]
- Hébert, S.S.; De Strooper, B. Alterations of the microRNA network cause neurodegenerative disease. Trends Neurosci. 2009, 32, 199–206. [Google Scholar] [CrossRef]
- Ai, J.; Zhang, R.; Li, Y.; Pu, J.; Lu, Y.; Jiao, J.; Li, K.; Yu, B.; Li, Z.; Wang, R.; et al. Circulating microRNA-1 as a potential novel biomarker for acute myocardial infarction. Biochem. Biophys. Res. Commun. 2010, 391, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-K.; Zhu, J.-Q.; Zhang, J.-T.; Li, Q.; Li, Y.; He, J.; Qin, Y.-W.; Jing, Q. Circulating microRNA: A novel potential biomarker for early diagnosis of acute myocardial infarction in humans. Eur. Heart J. 2010, 31, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, I.; Eran, A.; Nishino, I.; Moggio, M.; Lamperti, C.; Amato, A.A.; Lidov, H.G.; Kang, P.B.; North, K.N.; Mitrani-Rosenbaum, S.; et al. Distinctive patterns of microRNA expression in primary muscular disorders. Proc. Natl. Acad. Sci. USA 2007, 104, 17016–17021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, L.B.; Wang, C.; Sørensen, K.; Bang-Berthelsen, C.H.; Hansen, L.; Andersen, M.-L.M.; Hougaard, P.; Juul, A.; Zhang, C.-Y.; Pociot, F.; et al. Circulating levels of MicroRNA from children with newly diagnosed type 1 diabetes and healthy controls: Evidence that miR-25 associates to residual beta-cell function and glycaemic control during disease progression. Exp. Diabetes Res. 2012, 2012, 896362. [Google Scholar] [CrossRef]
- Sun, Y.; Gui, H.; Li, Q.; Luo, Z.M.; Zheng, M.J.; Duan, J.L.; Liu, X. MicroRNA-124 protects neurons against apoptosis in cerebral ischemic stroke. CNS Neurosci. Ther. 2013, 19, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.-T.; Lei, P.; Wang, H.-C.; Zhang, A.-L.; Han, Z.-L.; Chen, X.; Li, S.-H.; Jiang, R.-C.; Kang, C.-S.; Zhang, J.-N. MiR-21 improves the neurological outcome after traumatic brain injury in rats. Sci. Rep. 2015, 4, 6718. [Google Scholar] [CrossRef] [PubMed]
- Neri, M.; Frati, A.; Turillazzi, E.; Cantatore, S.; Cipolloni, L.; Di Paolo, M.; Frati, P.; La Russa, R.; Maiese, A.; Scopetti, M.; et al. Immunohistochemical evaluation of aquaporin-4 and its correlation with CD68, IBA-1, HIF-1α, GFAP, and CD15 expressions in fatal traumatic brain injury. Int. J. Mol. Sci. 2018, 19, 3544. [Google Scholar] [CrossRef]
- Petito, A.; Altamura, M.; Iuso, S.; Padalino, F.A.; Sessa, F.; D’Andrea, G.; Margaglione, M.; Bellomo, A. The relationship between personality traits, the 5HTT polymorphisms, and the occurrence of anxiety and depressive symptoms in elite athletes. PLoS ONE 2016, 11, e0156601. [Google Scholar] [CrossRef] [PubMed]
- Zaninotto, A.L.; Vicentini, J.E.; Fregni, F.; Rodrigues, P.A.; Botelho, C.; de Lucia, M.C.S.; Paiva, W.S. Updates and current perspectives of psychiatric assessments after traumatic brain injury: A systematic review. Front. Psychiatry 2016, 7, 95. [Google Scholar] [CrossRef]
- Pinchi, E.; Frati, A.; Cipolloni, L.; Aromatario, M.; Gatto, V.; La Russa, R.; Pesce, A.; Santurro, A.; Fraschetti, F.; Frati, P.; et al. Clinical-pathological study on β-APP, IL-1β, GFAP, NFL, Spectrin II, 8OHdG, TUNEL, MIR-21, MIR-16, MIR-92 expressions to verify DAI-diagnosis, grade and prognosis. Sci. Rep. 2018, 8, 2387. [Google Scholar] [CrossRef]
- Liu, W.; Chen, X.; Zhang, Y. Effects of microRNA-21 and microRNA-24 inhibitors on neuronal apoptosis in ischemic stroke. Am. J. Transl. Res. 2016, 8, 3179. [Google Scholar]
- Han, Z.; Chen, F.; Ge, X.; Tan, J.; Lei, P.; Zhang, J. MiR-21 alleviated apoptosis of cortical neurons through promoting PTEN-Akt signaling pathway in vitro after experimental traumatic brain injury. Brain Res. 2014, 1582, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.C.; Liao, Y.C.; Wang, Y.S.; Lin, H.F.; Lin, R.T.; Juo, S.H.H. Serum microrna-21 and microrna-221 as potential biomarkers for cerebrovascular disease. J. Vasc. Res. 2013, 50, 346–354. [Google Scholar] [CrossRef]
- Buller, B.; Liu, X.; Wang, X.; Zhang, R.L.; Zhang, L.; Hozeska-Solgot, A.; Chopp, M.; Zhang, Z.G. MicroRNA-21 protects neurons from ischemic death. FEBS J. 2010, 277, 4299–4307. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, A.D.; Fonken, L.K.; Watkins, L.R.; Nelson, R.J.; Popovich, P.G. MicroRNAs: Roles in regulating neuroinflammation. Neuroscientist 2018, 24, 221–245. [Google Scholar] [CrossRef]
- Jeyaseelan, K.; Lim, K.; Armugam, A. Neuroprotectants in stroke therapy. Expert Opin. Pharmacother. 2008, 9, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.H.; Chen, J.; Clark, R.S. Bcl-2 family gene products in cerebral ischemia and traumatic brain injury. J.Neurotrauma 2000, 17, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, V.; Dreyer, J.L. MicroRNAs miR-124, let-7d and miR-181a regulate cocaine-induced plasticity. Mol. Cell. Neurosci. 2009, 42, 350–362. [Google Scholar] [CrossRef]
- Sun, Y.; Luo, Z.-M.; Guo, X.-M.; Su, D.-F.; Liu, X. An updated role of microRNA-124 in central nervous system disorders: A review. Front. Cell. Neurosci. 2015, 9, 193. [Google Scholar] [CrossRef]
- Rajasethupathy, P.; Fiumara, F.; Sheridan, R.; Betel, D.; Puthanveettil, S.V.; Russo, J.J.; Sander, C.; Tuschl, T.; Kandel, E. Characterization of small RNAs in aplysia reveals a role for miR-124 in constraining synaptic plasticity through CREB. Neuron 2009, 63, 803–817. [Google Scholar] [CrossRef]
- Khanna, A.; Muthusamy, S.; Liang, R.; Sarojini, H.; Wang, E. Gain of survival signaling by down-regulation of three key miRNAs in brain of calorie-restricted mice. Aging Albany NY 2011, 3, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Zovoilis, A.; Agbemenyah, H.Y.; Agis-Balboa, R.C.; Stilling, R.M.; Edbauer, D.; Rao, P.; Farinelli, L.; Delalle, I.; Schmitt, A.; Falkai, P.; et al. MicroRNA-34c is a novel target to treat dementias. EMBO J. 2011, 30, 4299–4308. [Google Scholar] [CrossRef] [PubMed]
- Karr, J.; Vagin, V.; Chen, K.; Ganesan, S.; Olenkina, O.; Gvozdev, V.; Featherstone, D.E. Regulation of glutamate receptor subunit availability by microRNAs. J. Cell Biol. 2009, 185, 685–697. [Google Scholar] [CrossRef] [Green Version]
- Nudelman, A.S.; Dirocco, D.P.; Lambert, T.J.; Garelick, M.G.; Le, J.; Nathanson, N.M.; Storm, D.R. Neuronal activity rapidly induces transcription of the CREB-regulated microRNA-132, in vivo. Hippocampus 2010, 20, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Luikart, B.W.; Bensen, A.S.L.; Washburn, E.K.; Perederiy, J.V.; Su, K.G.; Li, Y.; Kernie, S.G.; Parada, L.F.; Westbrook, G.L. MiR-132 mediates the integration of newborn neurons into the adult dentate gyrus. PLoS ONE 2011, 6, e19077. [Google Scholar] [CrossRef]
- Hadar, A.; Milanesi, E.; Walczak, M.; Puzianowska-Kuźnicka, M.; Kuźnicki, J.; Squassina, A.; Niola, P.; Chillotti, C.; Attems, J.; Gozes, I.; et al. SIRT1, miR-132 and miR-212 link human longevity to Alzheimer’s Disease. Sci. Rep. 2018, 8, 8465. [Google Scholar] [CrossRef]
- Vo, N.; Klein, M.E.; Varlamova, O.; Keller, D.M.; Yamamoto, T.; Goodman, R.H.; Impey, S. From The Cover: A cAMP-response element binding protein-induced microRNA regulates neuronal morphogenesis. Proc. Natl. Acad. Sci. USA 2005, 102, 16426–16431. [Google Scholar] [CrossRef] [PubMed]
- Wayman, G.A.; Davare, M.; Ando, H.; Fortin, D.; Varlamova, O.; Cheng, H.-Y.M.; Marks, D.; Obrietan, K.; Soderling, T.R.; Goodman, R.H.; et al. An activity-regulated microRNA controls dendritic plasticity by down-regulating p250GAP. Proc. Natl. Acad. Sci. USA 2008, 105, 9093–9098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanet, A.; Tacheny, A.; Arnould, T.; Renard, P. MiR-212/132 expression and functions: Within and beyond the neuronal compartment. Nucleic Acids Res. 2012, 40, 4742–4753. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J. Boosting the brain’s ability to block inflammation via MICRORNA-132. Immunity 2009, 31, 854–855. [Google Scholar] [CrossRef] [PubMed]
- Shaked, I.; Meerson, A.; Wolf, Y.; Avni, R.; Greenberg, D.; Gilboa-Geffen, A.; Soreq, H. MicroRNA-132 potentiates cholinergic anti-inflammatory signaling by targeting acetylcholinesterase. Immunity 2009, 31, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Hollander, J.A.; Im, H.I.; Amelio, A.L.; Kocerha, J.; Bali, P.; Lu, Q.; Willoughby, D.; Wahlestedt, C.; Conkright, M.D.; Kenny, P.J. Striatal microRNA controls cocaine intake through CREB signalling. Nature 2010, 466, 197–202. [Google Scholar] [CrossRef]
- Sadakierska-Chudy, A.; Frankowska, M.; Miszkiel, J.; Wydra, K.; Jastrzębska, J.; Filip, M. Prolonged induction of miR-212/132 and REST expression in rat striatum following cocaine self-administration. Mol. Neurobiol. 2017, 54, 2241–2254. [Google Scholar] [CrossRef] [PubMed]
- Buller, B.; Chopp, M.; Ueno, Y.; Zhang, L.; Zhang, R.L.; Morris, D.; Zhang, Y.; Zhang, Z.G. Regulation of serum response factor by miRNA-200 and miRNA-9 modulates oligodendrocyte progenitor cell differentiation. Glia 2012, 60, 1906–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadhav, S.P.; Kamath, S.P.; Choolani, M.; Lu, J.; Dheen, S.T. MicroRNA-200b modulates microglia-mediated neuroinflammation via the cJun/MAPK pathway. J. Neurochem. 2014, 130, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Ciesielski-Treska, J.; Grant, N.J.; Ulrich, G.; Corrotte, M.; Bailly, Y.; Haeberle, A.M.; Chasserot-Golaz, S.; Bader, M.F. Fibrillar prion peptide (106–126) and scrapie prion protein hamper phagocytosis in microglia. Glia 2004, 46, 101–115. [Google Scholar] [CrossRef]
- Park, J.Y.; Paik, S.R.; Jou, I.; Park, S.M. Microglial phagocytosis is enhanced by monomeric α-synuclein, not aggregated α-synuclein: Implications for Parkinson’s disease. Glia 2008, 56, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T. The biphasic role of microglia in Alzheimer’s disease. Int. J. Alzheimers. Dis. 2012, 2012, 737846. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M.E.; Barres, B.A. Why is wallerian degeneration in the CNS so slow? Annu. Rev. Neurosci. 2007, 30, 153–179. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Expression Levels (Endogenous Control: miR-186) | ||||||
|---|---|---|---|---|---|---|
| Groups | miR-132 | miR-200b | miR-21 | miR-34 | miR-124 | |
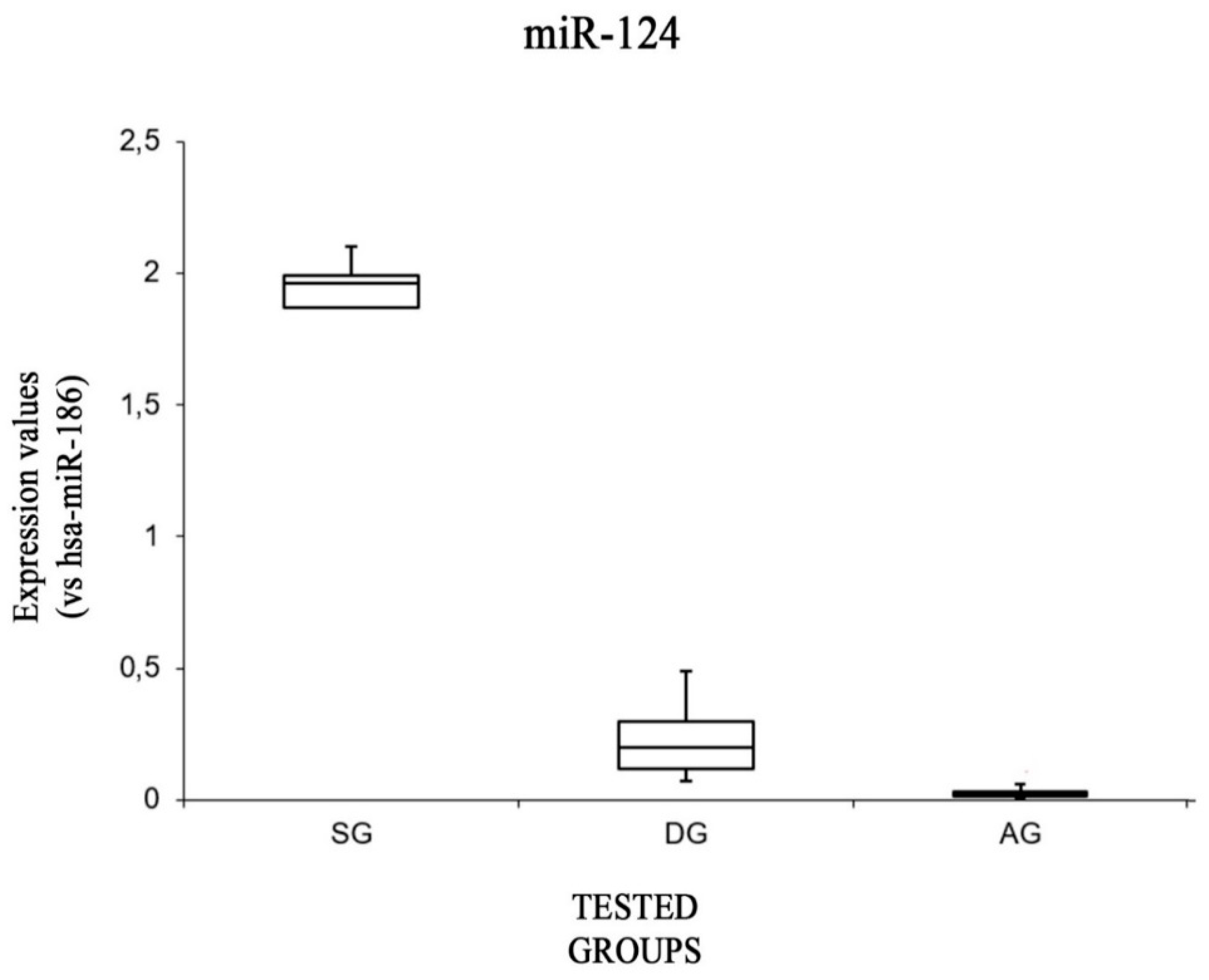
| SG | ∆CT (Mean Values) | 3.93 ± 1.98 | −0.52 ± 0.40 | −2.06 ± 0.20 | 3.81 ± 1.40 | −1.40 ± 0.07 |
| ∆∆CT (Mean Values) | 1.47 ± 1.98 | −0.06± 0.40 | −0.74 ± 0.20 | −0.84 ± 1.40 | −0.96 ± 0.07 | |
| Fold Difference = 2−∆∆CT (Mean Values) | 0.62 ± 0.53 | 1.07 ± 0.31 | 1.69 ± 0.23 | 2.47 ± 1.91 | 1.95 ± 0.09 | |
| DG | ∆CT (Mean Values) | 2.17 ± 0.44 | 0.81 ± 0.91 | −0.92 ± 0.13 | 0.95 ± 0.19 | 1.91 ± 1.05 |
| ∆∆CT (Mean Values) | −3.41 ± 0.44 | 1.27 ± 0.91 | 0.38 ± 0.13 | −3.69 ± 0.19 | 2.35 ± 1.05 | |
| Fold Difference = 2−∆∆CT (Mean Values) | 11.042 ± 2.91 | 0.47 ± 0.27 | 0.76 ± 0.06 | 13.06 ± 1.65 | 0.23 ± 0.16 | |
| AG | ∆CT (Mean Values) | 2.16 ± 0.72 | −0.49 ± 0.26 | −2.16 ± 0.48 | 3.63 ± 0.33 | 5.42 ± 0.73 |
| ∆∆CT (Mean Values) | −0.29 ± 0.72 | −0.02 ± 0.26 | −0.95 ± 0.48 | −0.76 ± 0.33 | 5.86 ± 0.73 | |
| Fold Difference = 2−∆∆CT (Mean Values) | 1.33 ± 0.53 | 1.02 ± 0.18 | 1.91 ± 0.61 | 2.07 ± 0.5 | 0.04 ± 0.05 | |
| Groups | Age | Cause of Death | Macroscopic Brain Findings | Microscopic Brain Findings |
|---|---|---|---|---|
| SG | ||||
| 1 | 80 | ischemic stroke | brain edema; reddish punctiform areas of the white matter | perineuronal and perivasal edema, red neurons, malacic areas |
| 2 | 79 | ischemic stroke from a vertebrobasilar embolism | brain edema; reddish punctiform areas of the white matter | perineuronal edema, red neurons |
| 3 | 34 | ischemic stroke in a subject with atheromatous formation at the Willis polygon | atherosclerotic alterations in the Willis polygon. | vasogenic edema, red neurons |
| 4 | 24 | ischemic stroke from a systemic massive bleeding | post-hypoxic malacic areas of the white matter | perivasal edema, red neurons, hemorrhages of Duret. |
| 5 | 72 | ischemic stroke | brain edema; reddish punctiform areas of the white matter | vasogenic edema, ischemic areas, “red neurons” |
| DG | ||||
| 1 | 29 | sudden cardiac death secondary to cocaine intake | brain edema | cortical edema |
| 2 | 32 | fatal ventricular arrhythmia secondary to cocaine intake | brain edema | vasogenic edema |
| 3 | 30 | sudden cardiac death secondary to cocaine intake | brain edema | perineuronal and perivasal edema |
| 4 | 20 | fatal cardiac arrhythmia secondary to cocaine intake | brain edema | cortical edema, arteriolosclerosis |
| 5 | 35 | fatal ventricular arrhythmia secondary to cocaine intake | brain edema, stasis | perineuronal edema, small perivascular hemorrhages |
| AG | ||||
| 1 | 80 | fatal cardiac arrhythmia | brain edema | vasogenic edema, arteriolosclerosis |
| 2 | 83 | ventricular arrhythmia | brain edema | perineuronal and perivasal edema |
| 3 | 77 | cardiac failure | brain edema | vasogenic edema, small periventricular hemorrhages |
| 4 | 65 | sudden cardiac death | brain edema | cortical edema |
| 5 | 74 | fatal cardiac arrhythmia | brain edema | perineuronal and perivasal edema, small perivascular hemorrhages |
| CTR | ||||
| 1 | 27 | acute cardiorespiratory arrest in subjects with aortic lesions | brain edema | perineuronal edema, stasis |
| 2 | 55 | acute cardiorespiratory arrest in subjects with multiple costal fracture and pulmonary contusions | brain edema | perineuronal and perivasal edema, small perivascular hemorrhages |
| Assay Name | Mature miRNA Sequence: | Chromosome Location |
|---|---|---|
| hsa-miR-132-3p | UAACAGUCUACAGCCAUGGUCG | Chr. 17-2049908-2050008 |
| hsa-miR-200b-3p | UAAUACUGCCUGGUAAUGAUGA | Chr.1: 1167104-1167198 |
| hsa-miR-21-5p | UAGCUUAUCAGACUGAUGUUGA | Chr.17: 59841266-59841337 |
| hsa-miR-34a-5p | UGGCAGUGUCUUAGCUGGUUGU | Chr.1: 9151668-9151777 |
| hsa-miR-124-5p | CGUGUUCACAGCGGACCUUGAU | Chr.8: 9903388-9903472 |
| Endogenous Control Genes | ||
| hsa-miR-186-5p | CAAAGAAUUCUCCUUUUGGGCU | Chr.1: 71067631-71067716 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sessa, F.; Maglietta, F.; Bertozzi, G.; Salerno, M.; Di Mizio, G.; Messina, G.; Montana, A.; Ricci, P.; Pomara, C. Human Brain Injury and miRNAs: An Experimental Study. Int. J. Mol. Sci. 2019, 20, 1546. https://doi.org/10.3390/ijms20071546
Sessa F, Maglietta F, Bertozzi G, Salerno M, Di Mizio G, Messina G, Montana A, Ricci P, Pomara C. Human Brain Injury and miRNAs: An Experimental Study. International Journal of Molecular Sciences. 2019; 20(7):1546. https://doi.org/10.3390/ijms20071546
Chicago/Turabian StyleSessa, Francesco, Francesca Maglietta, Giuseppe Bertozzi, Monica Salerno, Giulio Di Mizio, Giovanni Messina, Angelo Montana, Pietrantonio Ricci, and Cristoforo Pomara. 2019. "Human Brain Injury and miRNAs: An Experimental Study" International Journal of Molecular Sciences 20, no. 7: 1546. https://doi.org/10.3390/ijms20071546