Structural Determinants of Substrate Specificity of Omega-3 Desaturases from Mortierella alpina and Rhizophagus irregularis by Domain-Swapping and Molecular Docking

Abstract
:1. Introduction
2. Results
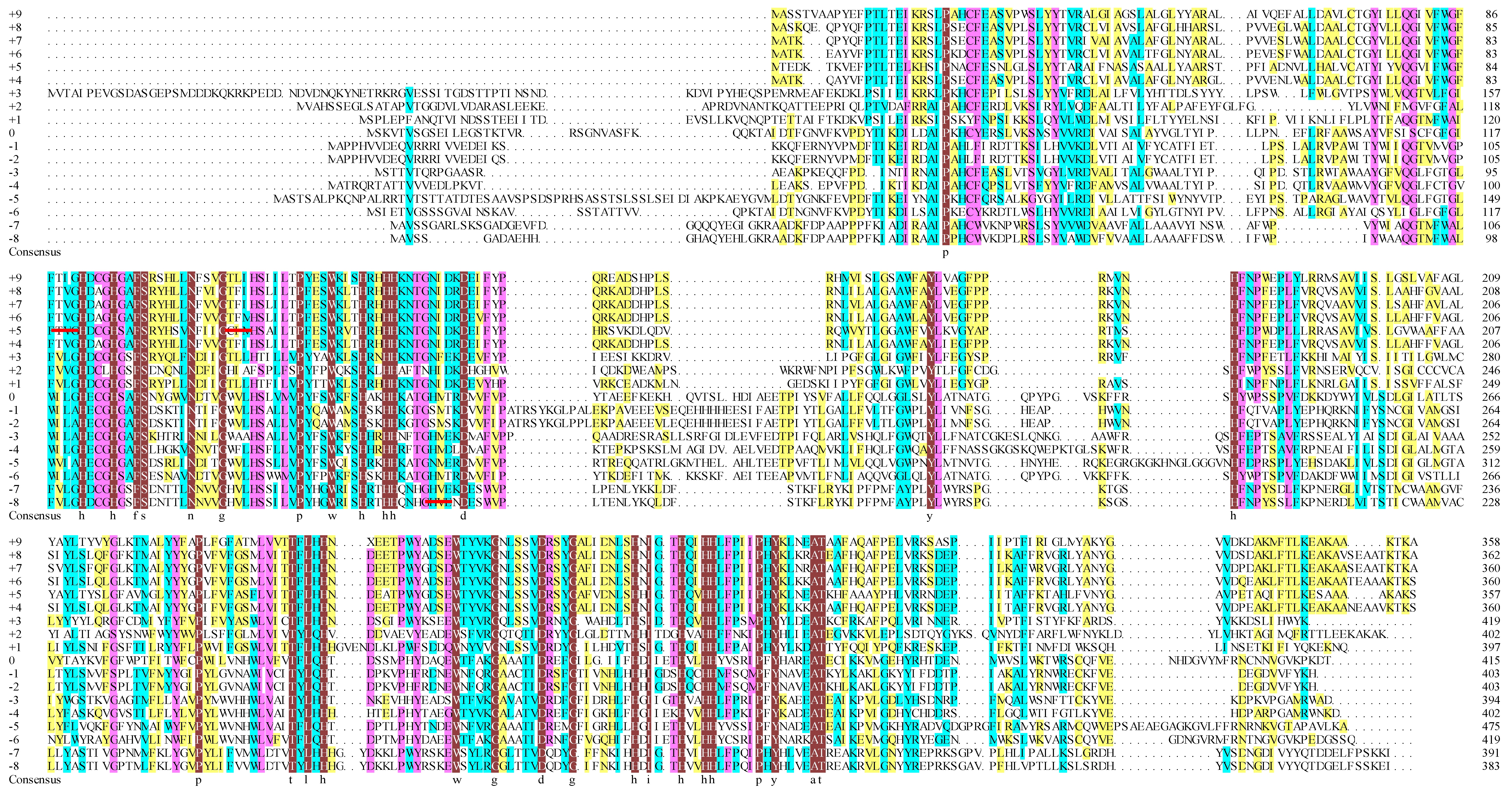
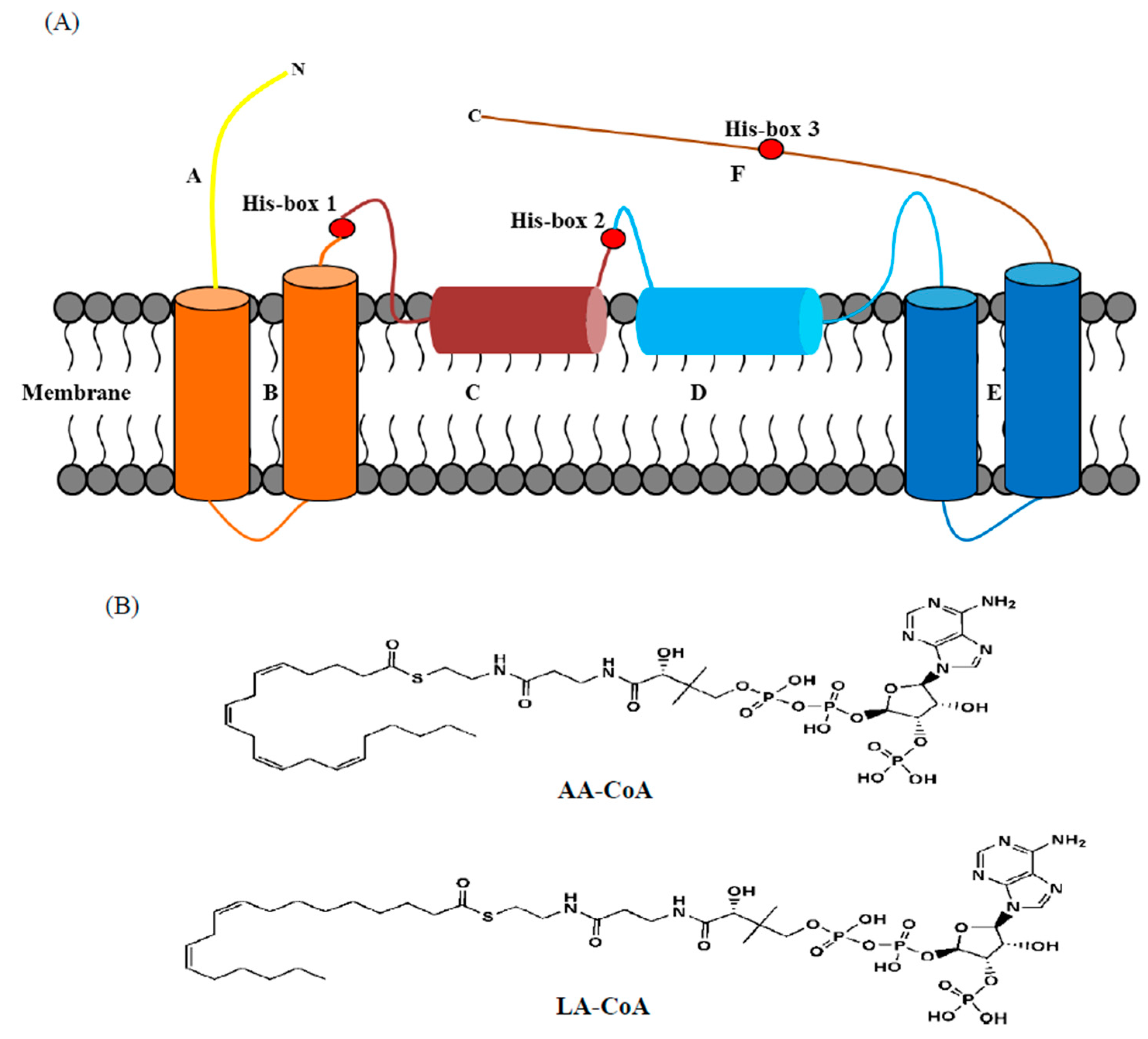
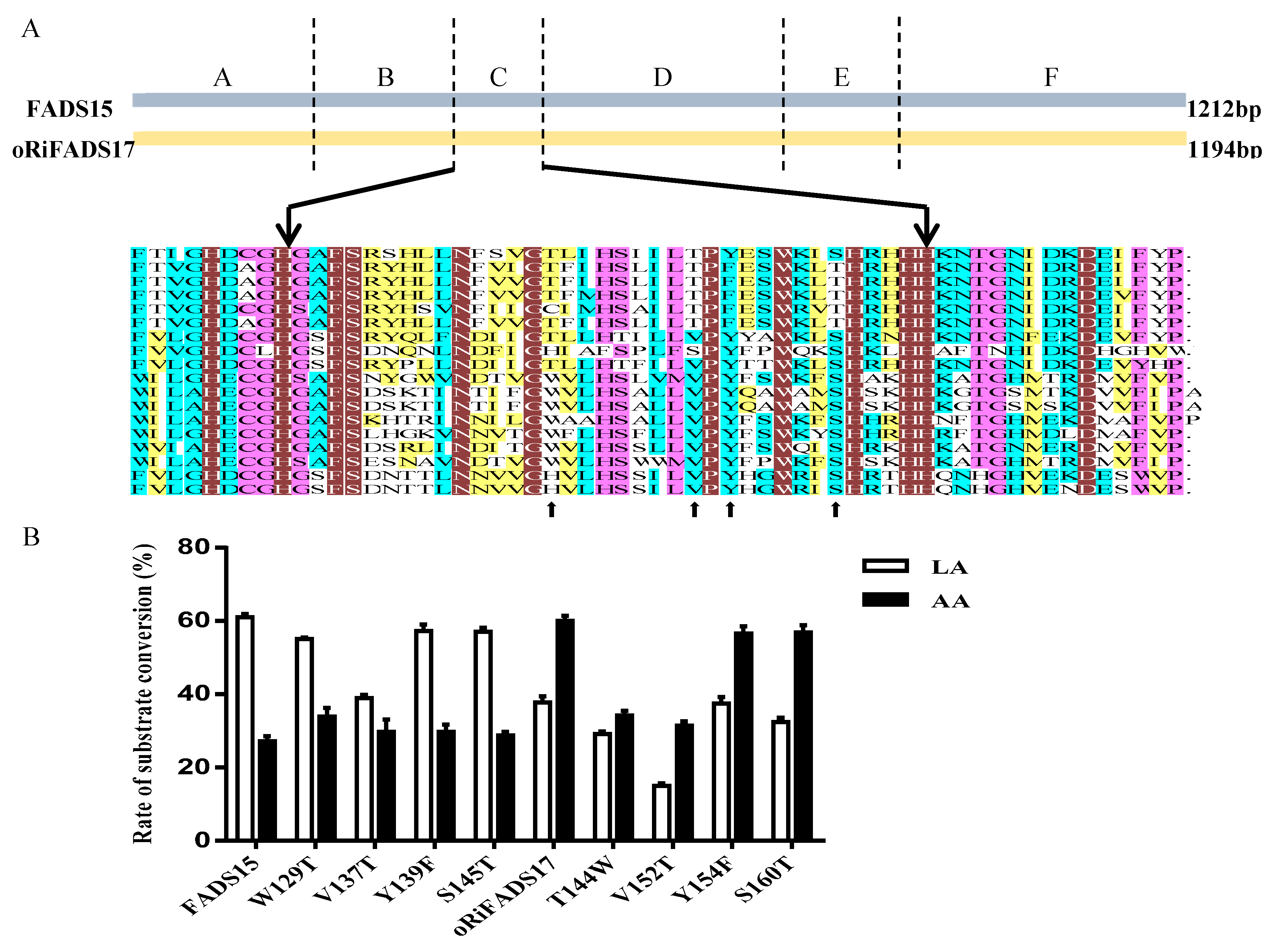
2.1. Similar Sequences and Topological Structures of FADS15 and oRiFADS17
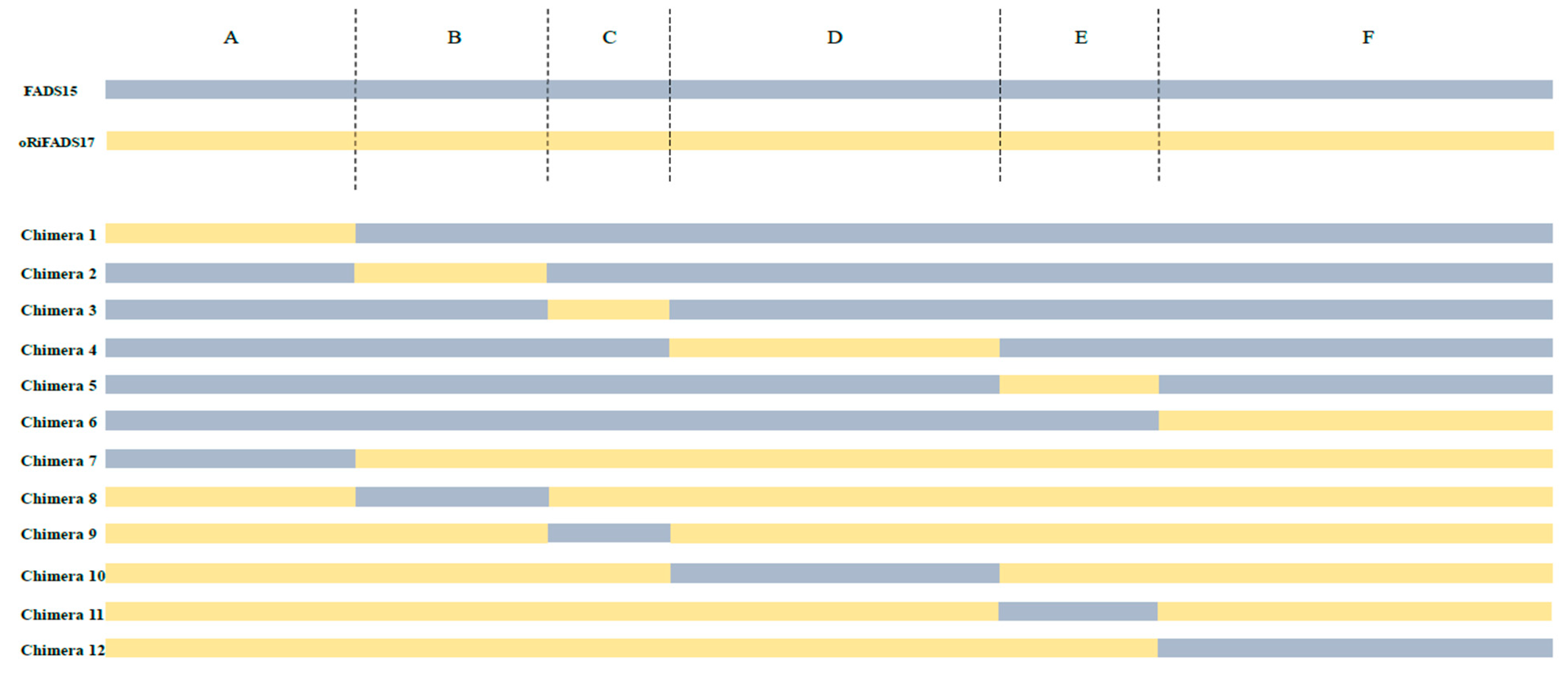
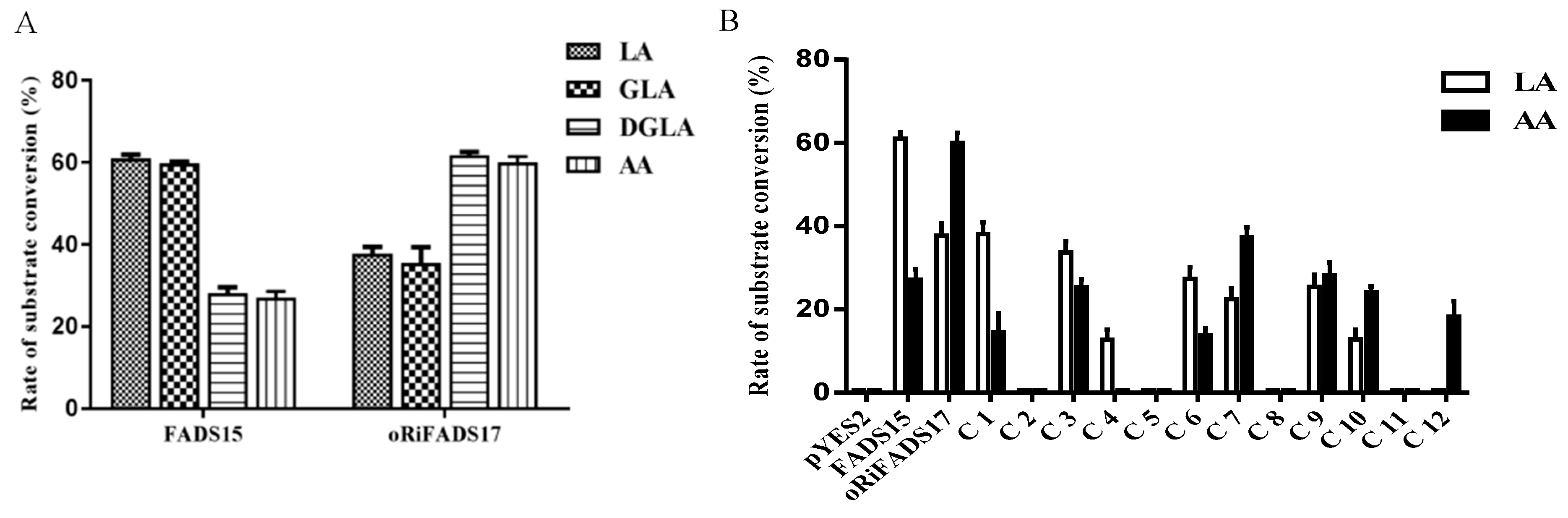
2.2. Identification of Determinants of Substrate Specificity in 12 Chimeras
2.3. Mutation Study of Amino Acids Responsible for Substrate Specificity
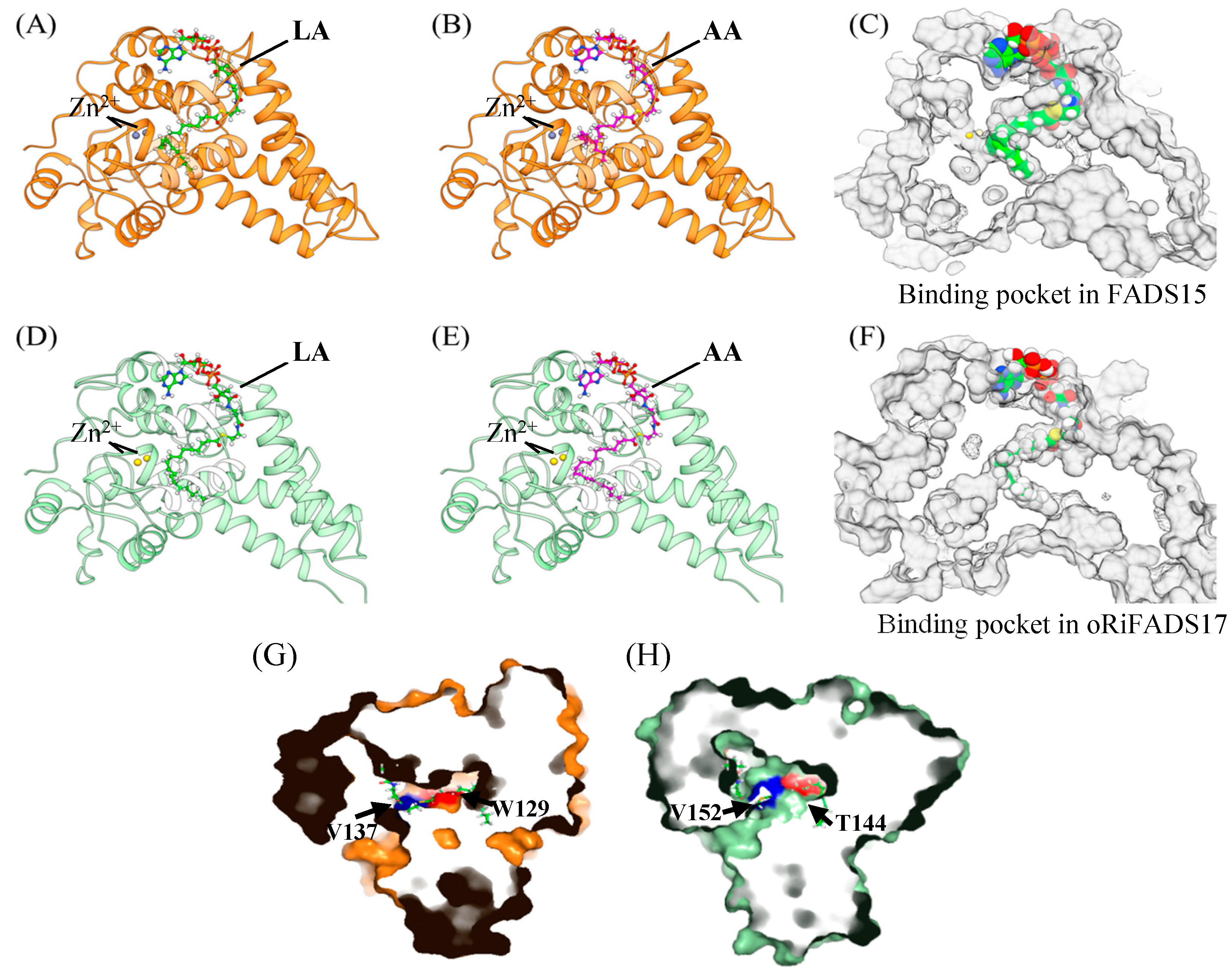
2.4. Homology Modelling and Molecular Docking Analysis of FADS15 and oRiFADS17
3. Discussion
4. Materials and Methods
4.1. Strains and Plasmids
4.2. Materials
4.3. Sequence Comparison and Topology Prediction
4.4. PCR Amplifications and Plasmid Constructions of Chimeric Enzymes and Mutants
4.5. Yeast Transformation and Heterologous Expression in S. cerevisiae
4.6. Lipid Extraction and Fatty Acid Analysis
4.7. Homology Modelling of 3-Dimensional Structures and Molecular Docking of FADS15 and oRiFADS17
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kikukawa, H.; Sakuradani, E.; Kishino, S.; Park, S.B.; Ando, A.; Shima, A.J.; Ochiai, M.; Shimizu, S.; Ogawa, J. Characterization of a trifunctional fatty acid desaturase from oleaginous filamentous fungus Mortierella alpina 1S-4 using a yeast expression system. J. Biosci. Bioeng. 2013, 116, 672–676. [Google Scholar] [CrossRef]
- Hirahashi, J.; Kawahata, K.; Arita, M.; Iwamoto, R.; Hishikawa, K.; Honda, M.; Hamasaki, Y.; Tanaka, M.; Okubo, K.; Kurosawa, M. Immunomodulation with eicosapentaenoic acid supports the treatment of autoimmune small-vessel vasculitis. Sci. Rep. 2014, 4, 6406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maehre, H.K.; Ida-Johanne, J.; Elvevoll, E.O.; Karl-Erik, E. ω-3 fatty acids and cardiovascular diseases: Effects, mechanisms and dietary relevance. Int. J. Mol. Sci. 2015, 16, 22636–22661. [Google Scholar] [CrossRef]
- Miyata, J.; Arita, M. Role of omega-3 fatty acids and their metabolites in asthma and allergic diseases. Allergol. Int. 2015, 64, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, S.L.; Leonard, A.E.; Pradip, M. Recent advances in the study of fatty acid desaturases from animals and lower eukaryotes. Prostag. Leukotr. Ess. Fatty Acids 2003, 68, 97–106. [Google Scholar] [CrossRef]
- Venegas, C.M.; Sayanova, O.; Napier, J.A. An alternative to fish oils: Metabolic engineering of oil-seed crops to produce omega-3 long chain polyunsaturated fatty acids. Prog. Lipid Res. 2010, 49, 108–119. [Google Scholar] [CrossRef]
- Oura, T.; Kajiwara, S. Saccharomyces kluyveri FAD3 encodes an omega3 fatty acid desaturase. Microbiology 2004, 150, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Oura, T.; Kajiwara, S. Substrate specificity and regioselectivity of Δ12 and ω3 fatty acid desaturases from Saccharomyces kluyveri. Biosci. Biotech. Bioch. 2008, 72, 3174–3179. [Google Scholar] [CrossRef]
- Damude, H.G.; Zhang, H.; Farrall, L.; Ripp, K.G.; Tomb, J.F.; Hollerbach, D.; Yadav, N.S. Identification of bifunctional Δ12/ω3 fatty acid desaturases for improving the ratio of ω3 to ω6 fatty acids in microbes and plants. Proc. Natl. Acad. Sci. USA 2006, 103, 9446–9451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, C.; Chen, H.; Wang, M.; Gu, Z.; Zhao, J.; Zhang, H.; Chen, W.; Chen, Y. Molecular mechanism of substrate preference for ω-3 fatty acid desaturase from Mortierella alpina by mutational analysis and molecular docking. Appl. Microbiol. Biotechnol. 2018, 102, 9679–9689. [Google Scholar] [CrossRef] [PubMed]
- Eiji, S.; Takahiro, A.; Keita, I.; Sakayu, S. A novel fungal ω3-desaturase with wide substrate specificity from arachidonic acid-producing Mortierella alpina 1S-4. Appl. Microbiol. Biot. 2005, 66, 648–654. [Google Scholar]
- Xue, Y.; Chen, B.; Win, A.N.; Fu, C.; Lian, J.; Liu, X.; Wang, R.; Zhang, X.; Chai, Y. ω-3 fatty acid desaturase gene family from two ω-3 sources, Salvia hispanica and Perilla frutescens: Cloning, characterization and expression. PLoS ONE 2018, 13, e0191432. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, M.; Gou, K. SDD17 desaturase can convert arachidonic acid to eicosapentaenoic acid in mammalian cells. Biochem. Biophys. Res. Commun. 2010, 394, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Fan, X.; Li, X.; Wang, H.; Chen, H. The desaturase OPIN17 from Phytophthora infestans converts arachidonic acid to eicosapentaenoic acid in CHO Cells. Appl. Biochem. Biotech. 2013, 171, 975–988. [Google Scholar] [CrossRef]
- Tang, X.; Chen, H.; Mei, T.; Ge, C.; Gu, Z.; Zhang, H.; Chen, Y.; Chen, W. Characterization of an ω-3 desaturase from Phytophthora parasitica and application for eicosapentaenoic acid production in Mortierella alpina. Front. Microbiol. 2018, 14, 1878. [Google Scholar] [CrossRef]
- Xue, Z.; He, H.; Hollerbach, D.; Macool, D.J.; Yadav, N.S.; Zhang, H.; Szostek, B.; Zhu, Q. Identification and characterization of new Δ-17 fatty acid desaturases. Appl. Microbiol. Biot. 2013, 97, 1973–1985. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hornung, E.; Busch, S.; Kassner, N.; Ternes, P.; Braus, S.; Kassner, N.; Ternes, P.; Braus, G.; Feussner, I. A small membrane-peripheral region close to the active centre determines regioselectivity of membrane-bound fatty acid desaturases from Aspergillus nidulans. J. Biol. Chem. 2007, 282, 26666–26674. [Google Scholar] [CrossRef]
- Shi, H.; Chen, H.; Gu, Z.; Song, Y.; Zhang, H.; Chen, W.; Chen, Y.Q. Molecular mechanism of substrate specificity for Δ 6 desaturase from Mortierella alpina and Micromonas pusilla. J. Lipid Res. 2015, 56, 2309–2321. [Google Scholar] [CrossRef]
- Lim, Z.L.; Senger, T.; Vrinten, P. Four amino acid residues influence the substrate chain-length and regioselectivity of Siganus canaliculatus Δ4 and Δ5/6 desaturases. Lipids 2014, 49, 357–367. [Google Scholar] [CrossRef]
- Song, L.; Zhang, Y.; Li, S.; Hu, J.; Yin, W.; Chen, Y.; Hao, S.; Wang, B.; Richard, R.C.; Hu, Z. Identification of the substrate recognition region in the delta(6)-fatty acid and delta(8)-sphingolipid desaturase by fusion mutagenesis. Planta 2014, 239, 753–763. [Google Scholar] [CrossRef]
- Vanhercke, T.; Shrestha, P.; Green, A.G.; Singh, S.P. Mechanistic and structural insights into the regioselectivity of an acyl-coa fatty acid desaturase via directed molecular evolution. J. Biol. Chem. 2011, 286, 12860–12869. [Google Scholar] [CrossRef]
- Meesapyodsuk, D.; Qiu, X. Structure determinants for the substrate specificity of acyl-CoA Δ9 desaturases from a marine copepod. ACS Chem. Biol. 2014, 9, 922. [Google Scholar] [CrossRef]
- Sakamoto, T.; Los, D.A.; Higashi, S.; Wada, H.; Nishida, I.; Ohmori, M.; Murata, N. Cloning of ω3 desaturase from Cyanobacteria and its use in altering the degree of membrane-lipid unsaturation. Plant. Mol. Biol. 1994, 26, 249–263. [Google Scholar] [CrossRef]
- Libisch, B.; Michaelson, L.V.; Lewis, M.J.P.; Shewry, R.; Napier, J.A. Chimeras of D6-fatty acid and D8-sphingolipid desaturases. Biochem. Biophys. Res. Commun. 2000, 279, 779–785. [Google Scholar] [CrossRef]
- Watanabe, K.; Ohno, M.; Taguchi, M.; Kawamoto, S.; Ono, K.; Aki, T. Identification of amino acid residues that determine the substrate specificity of mammalian membrane-bound front-end fatty acid desaturases. J. Lipid Res. 2015, 57, 89. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Wang, L.; Chen, W.; Feng, Y.; Ren, Y.; Gu, Z.; Chen, H.; Wang, H.; Thomas, M.J.; Zhang, B.; Berquin, I.M. Genome characterization of the oleaginous fungus Mortierella alpina. PLoS ONE. 2011, 6, e28319. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Mccoy, J.G.; Levin, E.J.; Zhou, M. X-ray structure of a mammalian stearoyl-coa desaturase-1. Nature 2015, 108, 534a. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Comput. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transformants | pYES2 (Control) | pYES2-FADS15 | pYES2-oRiFADS17 | ||||
|---|---|---|---|---|---|---|---|
| Fatty acid | + LA | + AA | + LA | + AA | + LA | + AA | |
| 16:0 (PA) | 25.4 ± 1.4 a | 25.6 ± 1.1 a | 23.3 ± 1.3 a | 24.3 ± 1.5 a | 25.8 ± 2.0 a | 22.9 ± 2.6 a | |
| 16:1 (PA) | 33.4 ± 1.7 a | 33.9 ± 3.1 a | 32.2 ± 1.8 a | 34.0 ± 1.8 a | 30.1 ± 1.6 a | 32.2 ± 3.2 a | |
| 18:0 (SA) | 4.5 ± 0.4 a | 4.7 ± 0.5 a | 4.6 ± 0.4 a | 4.3 ± 0.4 a | 5.1 ± 0.3 a | 5.0 ± 0.6 a | |
| 18:1 (OA) | 26.0 ± 2.1 a | 25.1 ± 1.1 a | 26.6 ± 1.1 a | 27.2 ± 2.6 a | 26.7 ± 1.5 a | 28.6 ± 3.5 a | |
| 18:2 (LA, ω-6) | 8.6 ± 0.5 a | ND | 3.2 ± 0.1 c | ND | 4.7 ± 0.3 b | ND | |
| 18:3 (GLA, ω-3) | ND | ND | 4.9 ± 0.3 a | ND | 2.9 ± 0.3 b | ND | |
| 20:4 (AA, ω-6) | ND | 5.8 ± 0.2 a | ND | 4.3 ± 0.3 b | ND | 2.0 ± 0.1 c | |
| 20:5 (EPA, ω-3) | ND | ND | ND | 1.6 ± 0.1 b | ND | 3.0 ± 0.3 a | |
| LA conversion rate | 0.0 ± 0.0 c | - | 61.0 ± 1.6 a | - | 37.8 ± 2.9 b | - | |
| AA conversion rate | - | 0.0 ± 0.0 c | - | 27.2 ± 2.6 b | - | 60.1 ± 2.4 a | |
| Fatty Acid | FADS15 Chimeras | oRiFADS17 Chimeras | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | C7 | C8 | C9 | C10 | C11 | C12 | |
| 16:0 (PA) | 25.3 ± 1.4 a | 21.8 ± 2.1 a | 24.4 ± 1.0 a | 24.6 ± 1.6 a | 23.6 ± 1.2 a | 20.2 ± 1.1 b | 23.7 ± 2.2 a | 23.0 ± 2.4 a | 24.7 ± 1.1 a | 22.1 ± 2.6 a | 22.1 ± 0.8 a | 22.1 ± 2.7 a |
| 16:1 (PA) | 30.4 ± 2.2 a | 31.6 ± 1.5 a | 30.0 ± 1.7 a | 31.2 ± 1.2 a | 32.4 ± 2.4 a | 32.1 ± 1.9 a | 30.4 ± 0.8 a | 31.4 ± 1.3 a | 30.8 ± 3.4 a | 33.4 ± 2.4 a | 30.7 ± 3.2 a | 29.7 ± 1.4 a |
| 18:0 (SA) | 4.0 ± 0.4 b | 4.8 ± 0.3 a | 4.7 ± 0.4 a,b | 4.0 ± 0.5 b | 5.0 ± 0.4 a,b | 4.9 ± 0.3 a,b | 5.3 ± 0.3 a,b | 5.3 ± 1.0 a,b | 4.8 ± 0.6 a,b | 4.6 ± 0.4 b | 5.7 ± 0.3 a | 5.2 ± 0.3 a,b |
| 18:1 (OA) | 23.1 ± 1.7 a | 23.6 ± 1.4 a | 22.3 ± 1.6 a,b | 22.2 ± 1.1 a,b | 21.6 ± 0.9 a,b | 24.4 ± 1.8 a | 21.6 ± 2.3 a,b | 20.9 ± 1.8 a,b | 18.6 ± 2.2 b | 23.4 ± 0.9 a | 21.3 ± 2.3 a,b | 22.8 ± 1.7 a |
| 18:2 (LA, ω-6) | 4.6 ± 0.3 c | 8.4 ± 1.1 a | 5.2 ± 0.3 c | 7.2 ± 0.3 b | 7.7 ± 0.5 a,b | 5.6 ± 0.5 c | 5.4 ± 0.5 c | 7.9 ± 0.4 a,b | 6.5 ± 0.1 b,c | 6.9 ± 0.1 b | 8.5 ± 0.4 a | 8.4 ± 0.4 a |
| 18:3 (GLA, ω-3) | 2.8 ± 0.2 a | ND | 2.7 ± 0.2 a,b | 1.0 ± 0.2 d | ND | 2.1 ± 0.1 b | 1.6 ± 0.1 c | ND | 2.2 ± 0.4 b | 1.0 ± 0.2 d | ND | ND |
| 20:4 (AA, ω-6) | 4.1 ± 0.3 b | 5.6 ± 0.8 a,b | 4.2 ± 0.2 b | 5.0 ± 0.5 b | 5.4 ± 0.3 a,b | 4.7 ± 0.3 b | 4.3 ± 0.6 b | 5.7 ± 0.4 a,b | 4.4 ± 0.4 b | 4.1 ± 0.2 b | 6.3 ± 0.4 a | 5.1 ± 0.8 b |
| 20:5 (EPA, ω-3) | 0.7 ± 0.2 d | ND | 1.4 ± 0.1 b,c | ND | ND | 0.8 ± 0.1 d | 2.6 ± 0.2 a | ND | 1.7 ± 0.3 b | 1.3 ± 0.1 c | ND | 1.2 ± 0.1 c |
| LA conversion rate | 38.1 ± 2.8 a | - | 33.7 ± 2.7 a | 12.7 ± 2.3 c | - | 27.4 ± 2.8 b | 22.5 ± 2.7 b | - | 25.5 ± 2.9 b | 12.9 ± 2.3 c | - | - |
| AA conversion rate | 14.5 ± 4.5 c | - | 25.4 ± 1.9 b | - | - | 13.7 ± 1.9 c | 37.4 ± 2.3 a | - | 28.1 ± 3.1 b | 24.1 ± 1.5 b,c | - | 18.2 ± 3.7 c |
| Fatty Acid | FADS15 Mutants | oRiFADS17 Mutants | ||||||
|---|---|---|---|---|---|---|---|---|
| W129T | V137T | Y139F | S145T | T144W | V152T | Y154F | S160T | |
| 16:0 (PA) | 23.9 ± 0.7 a,b | 23.2 ± 0.7 a,b | 23.4 ± 2.4 a,b | 25.3 ± 1.1 a | 23.9 ± 1.5 a,b | 20.9 ± 2.0 b | 22.7 ± 1.8 a,b | 23.0 ± 1.0 a,b |
| 16:1 (PA) | 30.8 ± 2.2 a | 34.1 ± 2.4 a | 30.7 ± 3.1a | 31.2 ± 2.8a | 32.1 ± 2.1 a | 31.8 ± 2.3 a | 30.1 ± 3.1 a | 31.1 ± 3.9 a |
| 18:0 (SA) | 5.6 ± 0.6 a | 3.7 ± 0.2 b | 4.6 ± 0.9 a,b | 4.6 ± 0.8 a,b | 5.3 ± 0.5 a | 5.3 ± 0.4 a | 4.9 ± 0.4 a,b | 4.8 ± 0.5 a,b |
| 18:1 (OA) | 21.1 ± 2.0 a | 23.3 ± 2.2 a | 22.1 ± 2.0 a | 21.8 ± 1.4 a | 20.9 ± 1.7 a | 23.8 ± 2.2 a | 23.2 ± 2.4 a | 22.2 ± 2.1 a |
| 18:2 (LA, ω-6) | 3.7 ± 0.3 c | 4.3 ± 0.1 b,c | 3.6 ± 0.3 c | 3.5 ± 0.1 c | 5.6 ± 0.2 a | 6.1 ± 0.4 a | 4.9 ± 0.3 b | 5.6 ± 0.4 a |
| 18:3 (GLA, ω-3) | 4.6 ± 0.3 a | 3.2 ± 0.2 b | 4.9 ± 0.2 a | 4.6 ± 0.5 a | 2.3 ± 0.2 c | 1.1 ± 0.1 d | 2.9 ± 0.2 b | 2.7 ± 0.1 b |
| 20:4 (AA, ω-6) | 3.8 ± 0.2 a | 3.6 ± 0.2 a | 3.8 ± 0.3 a | 3.6 ± 0.4 a | 3.4 ± 0.3 a | 3.9 ± 0.5 a | 2.6 ± 0.1 b | 2.5 ± 0.3b |
| 20:5 (EPA, ω-3) | 2.0 ± 0.4 b | 1.3 ± 0.2 b | 1.6 ± 0.3 b | 1.5 ± 0.1 b | 1.8 ± 0.3 b | 1.8 ± 0.2 b | 3.4 ± 0.4 a | 3.3 ± 0.2 a |
| LA conversion rate | 55.1 ± 0.5 a | 42.4 ± 2.0 b | 57.2 ± 3.0 a | 57.0 ± 2.0 a | 29.2 ± 1.1 c | 15.0 ± 1.2 d | 37.4 ± 3.2 b | 32.5 ± 1.9 c |
| AA conversion rate | 33.9 ± 4.1 b | 28.0 ± 1.2 b | 29.7 ± 3.5 b | 28.7 ± 1.8 b | 34.2 ± 2.1 b | 31.3 ± 2.2 b | 56.5 ± 3.4 a | 56.8 ± 3.4 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rong, C.; Chen, H.; Tang, X.; Gu, Z.; Zhao, J.; Zhang, H.; Chen, Y.; Chen, W. Structural Determinants of Substrate Specificity of Omega-3 Desaturases from Mortierella alpina and Rhizophagus irregularis by Domain-Swapping and Molecular Docking. Int. J. Mol. Sci. 2019, 20, 1603. https://doi.org/10.3390/ijms20071603
Rong C, Chen H, Tang X, Gu Z, Zhao J, Zhang H, Chen Y, Chen W. Structural Determinants of Substrate Specificity of Omega-3 Desaturases from Mortierella alpina and Rhizophagus irregularis by Domain-Swapping and Molecular Docking. International Journal of Molecular Sciences. 2019; 20(7):1603. https://doi.org/10.3390/ijms20071603
Chicago/Turabian StyleRong, Chunchi, Haiqin Chen, Xin Tang, Zhennan Gu, Jianxin Zhao, Hao Zhang, Yongquan Chen, and Wei Chen. 2019. "Structural Determinants of Substrate Specificity of Omega-3 Desaturases from Mortierella alpina and Rhizophagus irregularis by Domain-Swapping and Molecular Docking" International Journal of Molecular Sciences 20, no. 7: 1603. https://doi.org/10.3390/ijms20071603