Expression and Activity of TRPA1 and TRPV1 in the Intervertebral Disc: Association with Inflammation and Matrix Remodeling

 , , and
, , and
Abstract
:
1. Introduction
2. Results
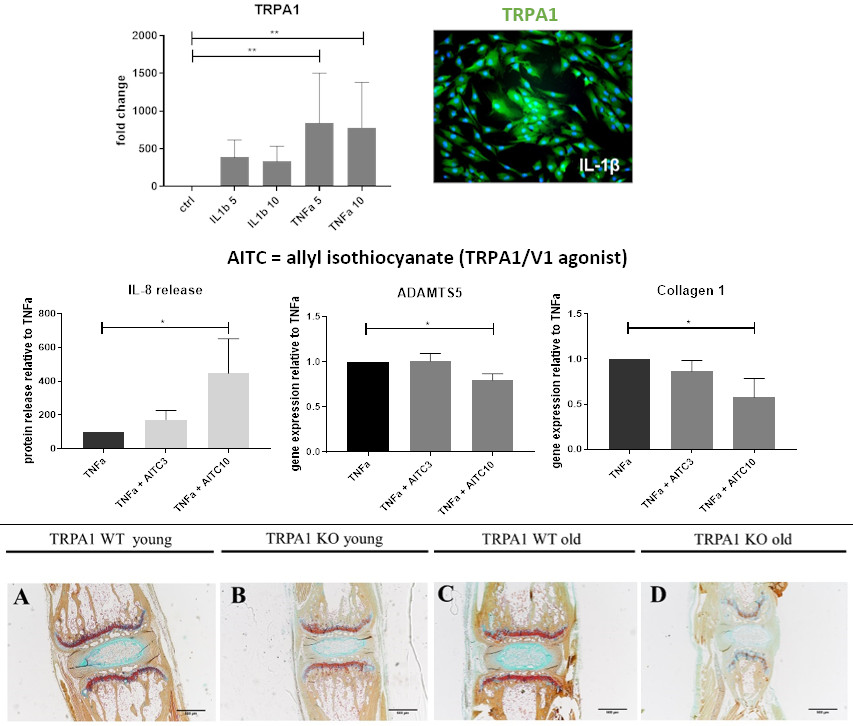
2.1. Gene Expression of TRPA1 in Human IVD Tissue
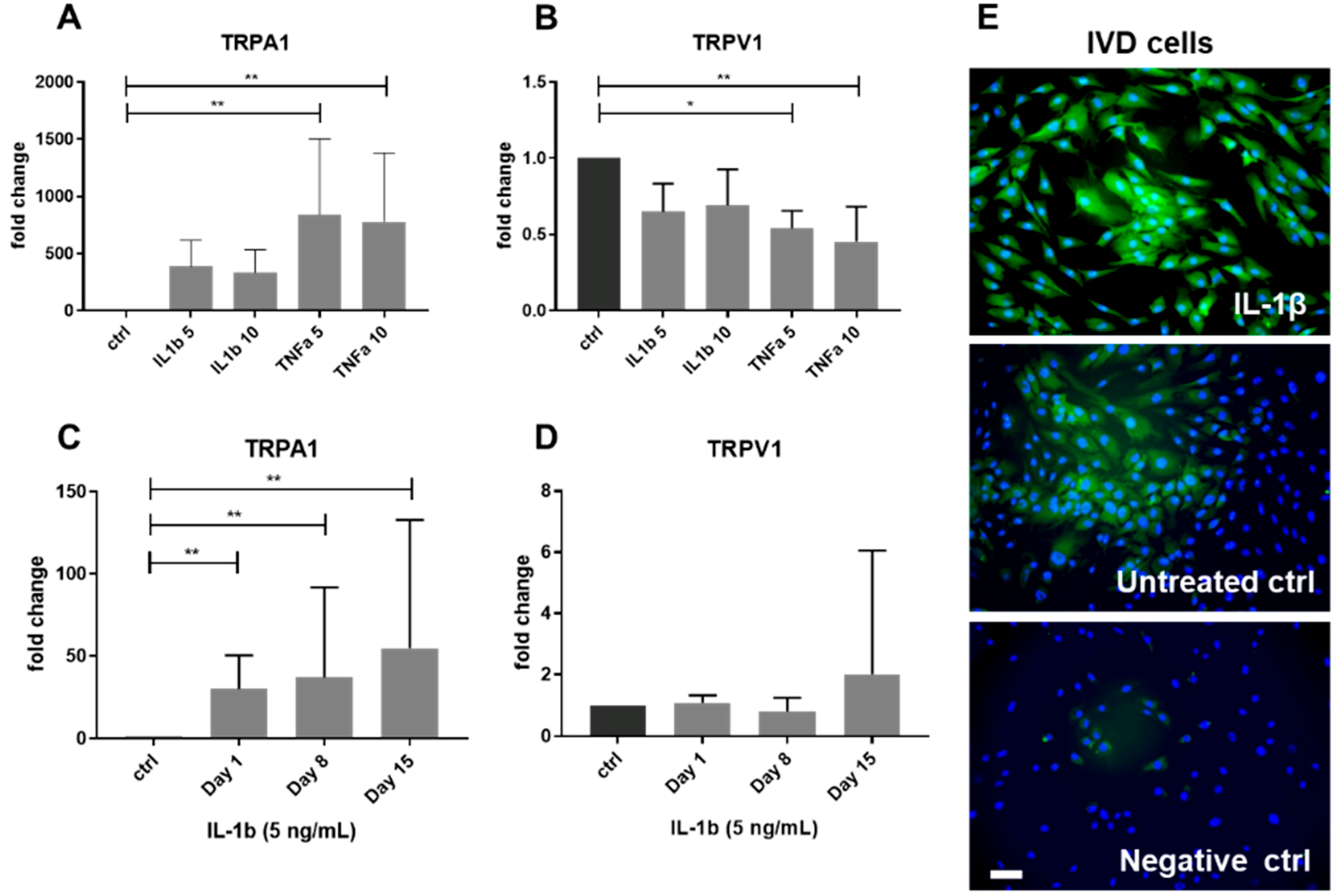
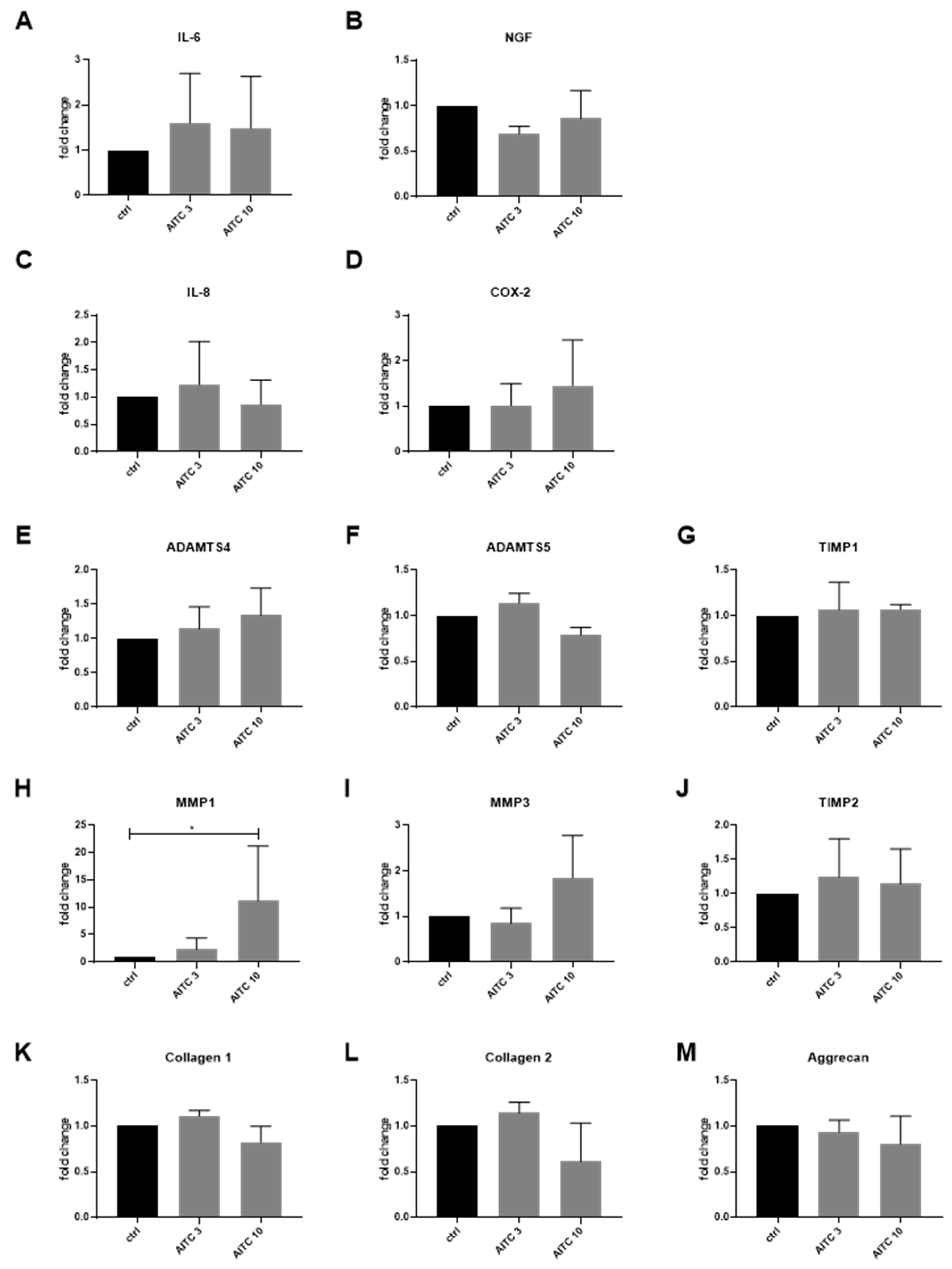
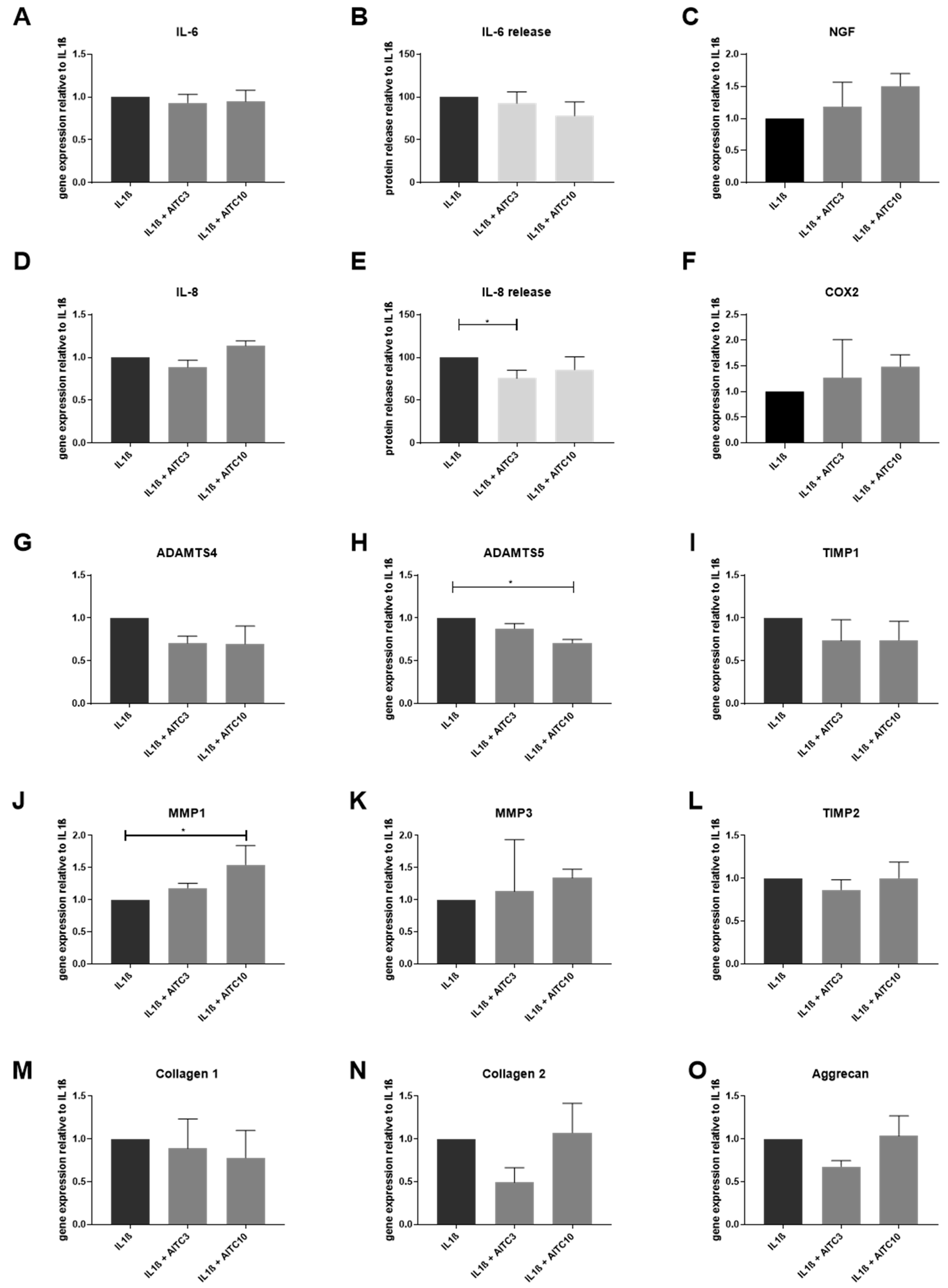
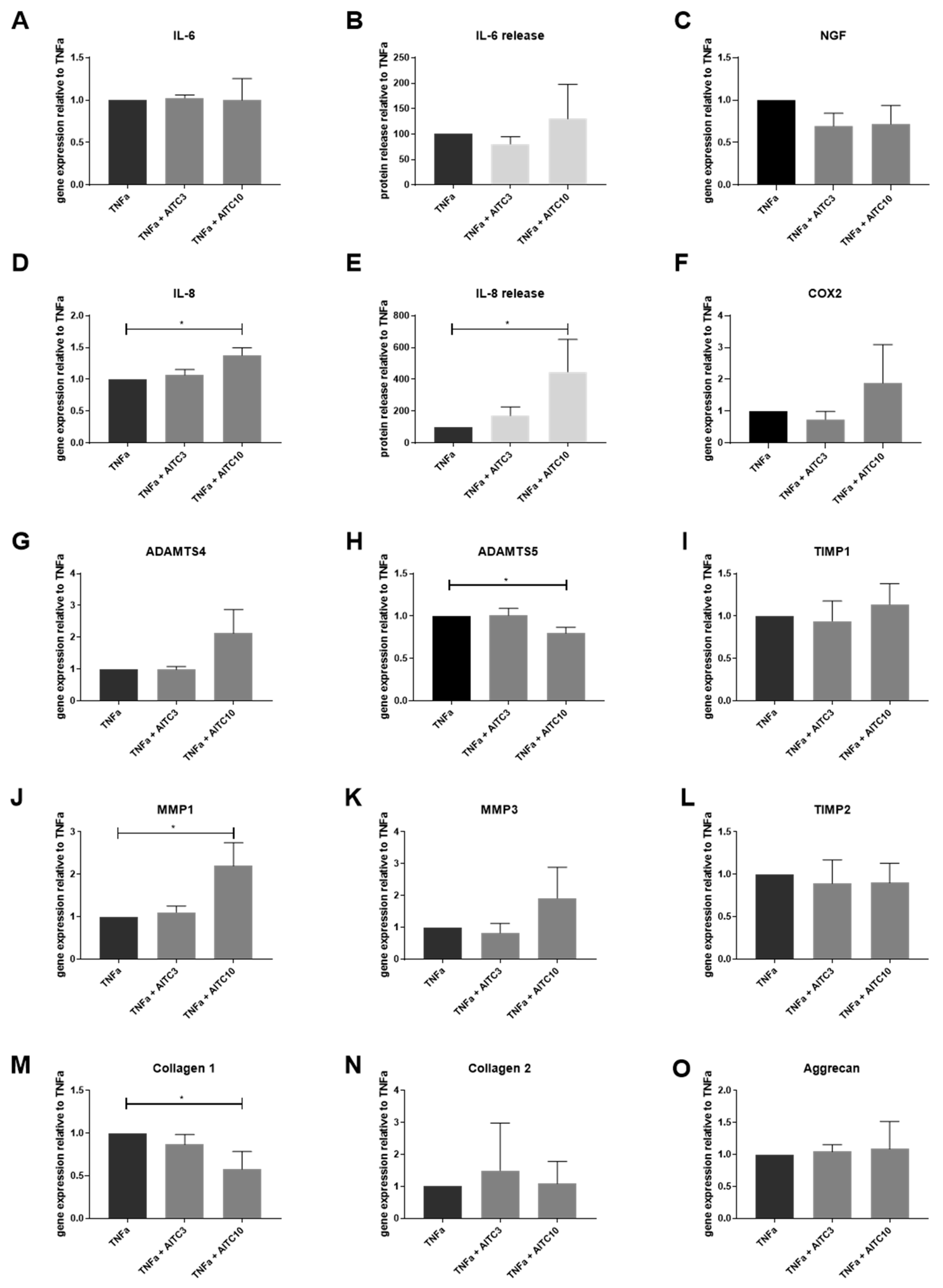
2.2. Gene Expression of TRPA1 and TRPV1 in Human IVD Cells Treated with Pro-Inflammatory Cytokines
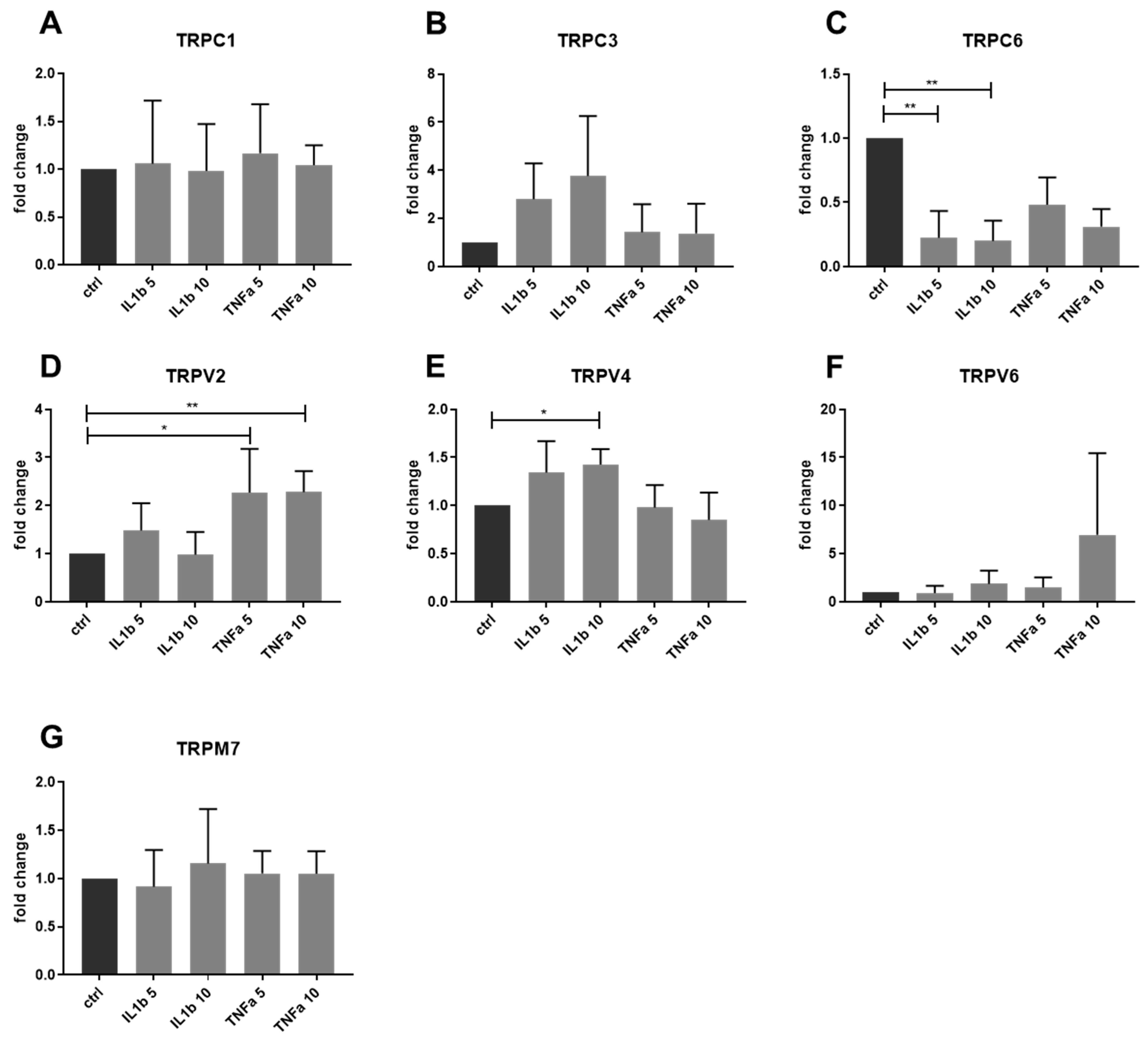
2.3. Gene Expression of Other TRP Channels in Human IVD Cells Treated with Pro-Inflammatory Cytokines
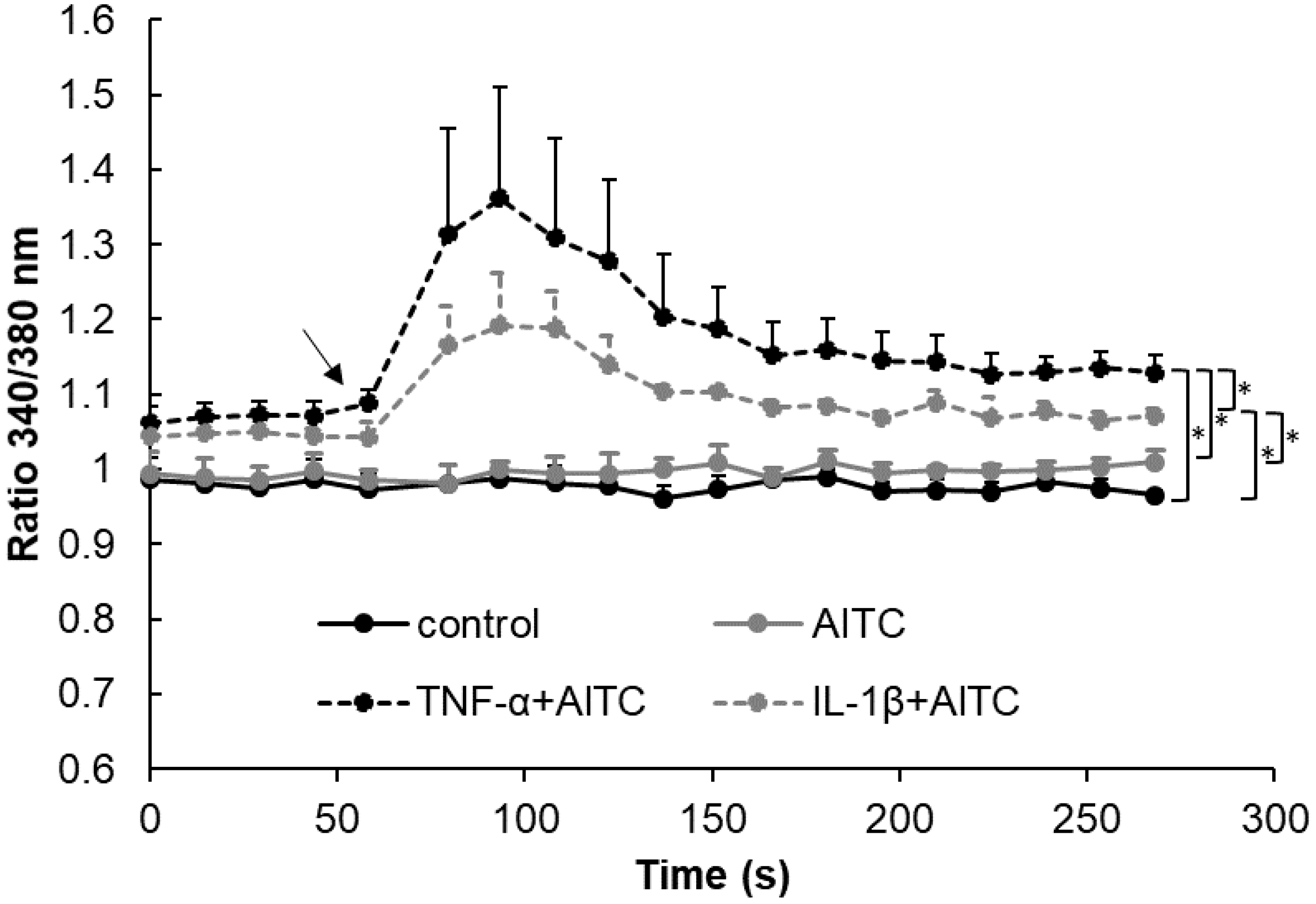
2.4. Functional Analysis of TRPA1 and TRPV1 in IVD Cells
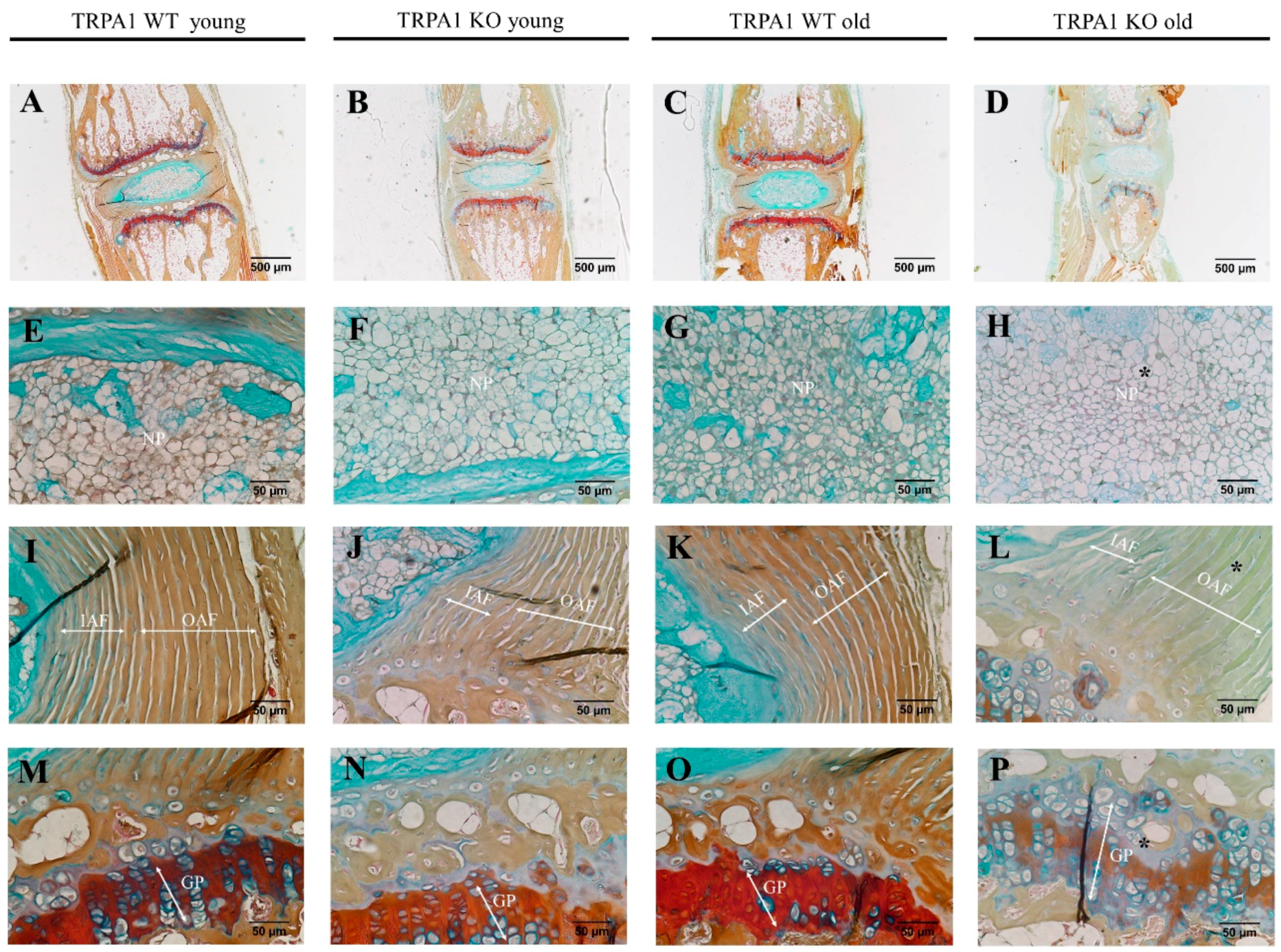
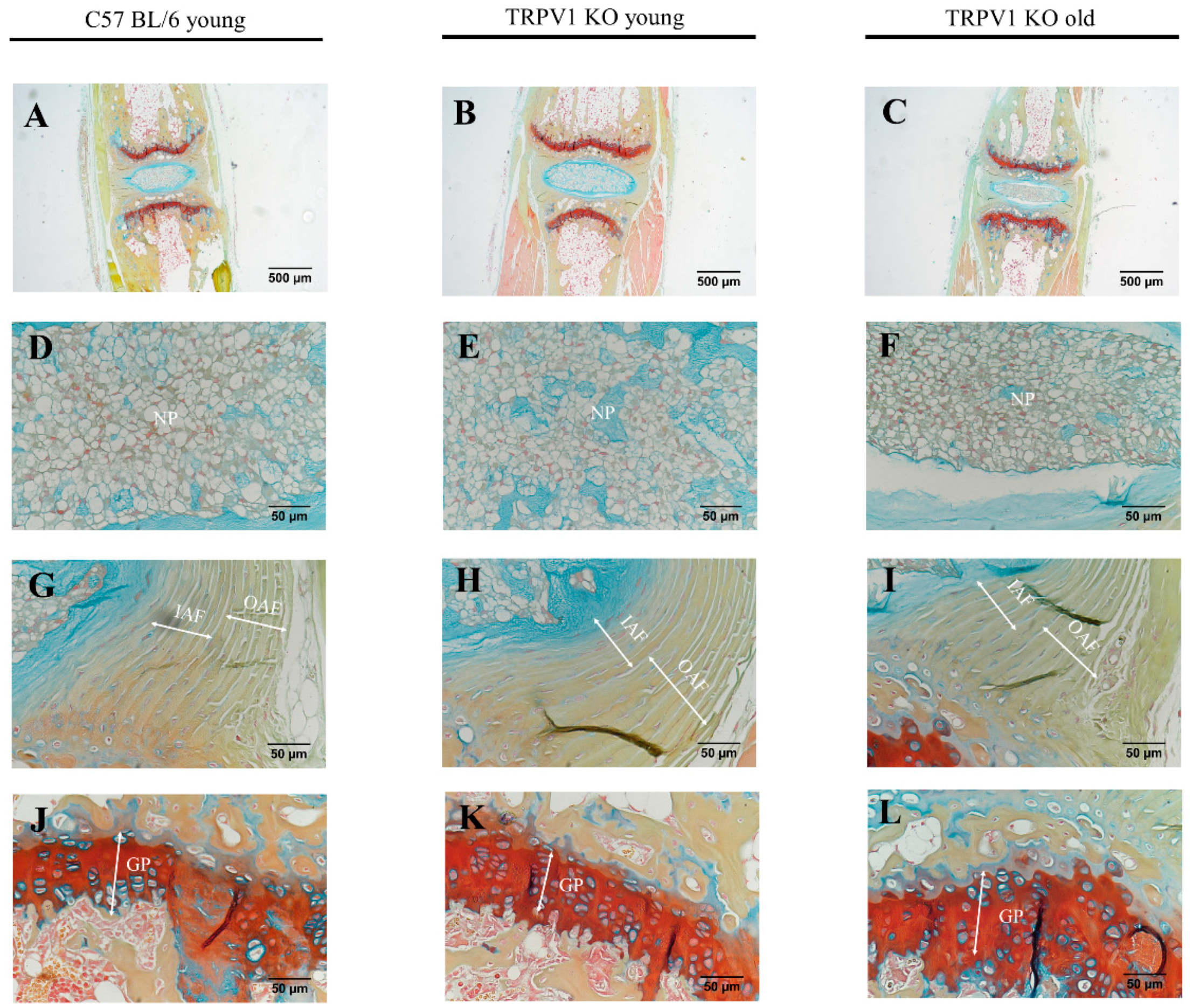
2.5. Motion Segments of TRPA1 and TRPV1-Deficient Mice
3. Discussion
4. Materials and Methods
4.1. Subjects
4.1.1. Non-Degenerated Human IVD Tissue
4.1.2. Human IVD Tissue
4.1.3. Human Fetal IVD Cells
4.1.4. Knock-Out Mice
4.2. Cell Culture
4.2.1. 2D Cell Culture
4.2.2. 3D Cell Culture
4.2.3. Cell Viability of 3D Cell Culture
4.3. Treatments
4.3.1. 2D Cell Cultures
4.3.2. 3D Cell Cultures
4.4. Analyses
4.4.1. Gene Expression Analysis of IVD Tissue
4.4.2. FAST Staining
4.4.3. Gene Expression Analysis of IVD Cells
4.4.4. Immunofluorescence
4.4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.4.6. [Ca2+]i Imaging
4.4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pai, S.; Sundaram, L.J. Low back pain: An economic assessment in the United States. Orthop. Clin. N. Am. 2004, 35, 1–5. [Google Scholar] [CrossRef]
- Ito, K.; Creemers, L. Mechanisms of intervertebral disk degeneration/injury and pain: A review. Glob. Spine J. 2013, 3, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, K.; Vo, N.; Kletsas, D.; Boos, N. Inflammatory and catabolic signalling in intervertebral discs: The roles of NF-kappaB and MAP kinases. Eur. Cell. Mater. 2012, 23, 103–119, discussion 119–120. [Google Scholar] [CrossRef] [PubMed]
- Vo, N.V.; Hartman, R.A.; Patil, P.R.; Risbud, M.V.; Kletsas, D.; Iatridis, J.C.; Hoyland, J.A.; Le Maitre, C.L.; Sowa, G.A.; Kang, J.D. Molecular mechanisms of biological aging in intervertebral discs. J. Orthop. Res. 2016, 34, 1289–1306. [Google Scholar] [CrossRef]
- Johnson, Z.I.; Schoepflin, Z.R.; Choi, H.; Shapiro, I.M.; Risbud, M.V. Disc in Flames: Roles of Tnf-alpha AND IL-1 beta in Intervertebral Disc Degeneration. Eur. Cell. Mater. 2015, 30, 104–117. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Kepler, C.K.; Markova, D.Z.; Hilibrand, A.S.; Vaccaro, A.R.; Risbud, M.V.; Albert, T.J.; Anderson, D.G. Substance P stimulates production of inflammatory cytokines in human disc cells. Spine 2013, 38, E1291–E1299. [Google Scholar] [CrossRef] [PubMed]
- Freemont, A.J.; Peacock, T.E.; Goupille, P.; Hoyland, J.A.; OBrien, J.; Jayson, M.I.V. Nerve ingrowth into diseased intervertebral disc in chronic back pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef]
- Stemkowski, P.L.; Noh, M.C.; Chen, Y.S.; Smith, P.A. Increased excitability of medium-sized dorsal root ganglion neurons by prolonged interleukin-1 exposure is K+ channel dependent and reversible. J. Physiol. 2015, 593, 3739–3755. [Google Scholar] [CrossRef]
- Wilkinson, M.F.; Earle, M.L.; Triggle, C.R.; Barnes, S. Interleukin-1beta, tumor necrosis factor-alpha, and LPS enhance calcium channel current in isolated vascular smooth muscle cells of rat tail artery. FASEB J. 1996, 10, 785–791. [Google Scholar] [CrossRef]
- Gavenis, K.; Schumacher, C.; Schneider, U.; Eisfeld, J.; Mollenhauer, J.; Schmidt-Rohlfing, B. Expression of ion channels of the TRP family in articular chondrocytes from osteoarthritic patients: Changes between native and in vitro propagated chondrocytes. Mol. Cell. Biochem. 2009, 321, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Krupkova, O.; Zvick, J.; Wuertz-Kozak, K. The role of transient receptor potential channels in joint diseases. Eur. Cell. Mater. 2017, 34, 180–201. [Google Scholar] [CrossRef] [PubMed]
- Patapoutian, A.; Tate, S.; Woolf, C.J. Transient receptor potential channels: Targeting pain at the source. Nat. Rev. Drug Discov. 2009, 8, 55–68. [Google Scholar] [CrossRef]
- Wu, L.J.; Sweet, T.B.; Clapham, D.E. International Union of Basic and Clinical Pharmacology. LXXVI. Current progress in the mammalian TRP ion channel family. Pharmacol. Rev. 2010, 62, 381–404. [Google Scholar] [CrossRef]
- Gees, M.; Colsoul, B.; Nilius, B. The Role of Transient Receptor Potential Cation Channels in Ca2+ Signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a003962. [Google Scholar] [CrossRef] [PubMed]
- Montell, C.; Birnbaumer, L.; Flockerzi, V.; Bindels, R.J.; Brudorf, E.A.; Caterina, M.J.; Clapham, D.E.; Harteneck, C.; Heller, S.; Julius, D.; et al. A unified nomenclature for the superfamily of TRP cation channels. Mol. Cell 2002, 9, 229–231. [Google Scholar] [CrossRef]
- Walter, B.A.; Purmessur, D.; Moon, A.; Occhiogrosso, J.; Laudier, D.M.; Hecht, A.C.; Iatridis, J.C. Reduced tissue osmolarity increases TRPV4 expression and pro-inflammatory cytokines in intervertebral disc cells. Eur. Cell. Mater. 2016, 32, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Franco-Obregon, A.; Cambria, E.; Greutert, H.; Wernas, T.; Hitzl, W.; Egli, M.; Sekiguchi, M.; Boos, N.; Hausmann, O.; Ferguson, S.J.; et al. TRPC6 in simulated microgravity of intervertebral disc cells. Eur. Spine J. 2018, 27, 2621–2630. [Google Scholar] [CrossRef]
- Sadowska, A.; Touli, E.; Hitzl, W.; Greutert, H.; Ferguson, S.J.; Wuertz-Kozak, K.; Hausmann, O.N. Inflammaging in cervical and lumbar degenerated intervertebral discs: Analysis of proinflammatory cytokine and TRP channel expression. Eur. Spine J. 2018, 27, 564–577. [Google Scholar] [CrossRef]
- Marrone, M.C.; Morabito, A.; Giustizieri, M.; Chiurchiu, V.; Leuti, A.; Mattioli, M.; Marinelli, S.; Riganti, L.; Lombardi, M.; Murana, E.; et al. TRPV1 channels are critical brain inflammation detectors and neuropathic pain biomarkers in mice. Nat. Commun. 2017, 8, 15292. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, E.S.; Fernandes, M.A.; Keeble, J.E. The functions of TRPA1 and TRPV1: Moving away from sensory nerves. Br. J. Pharmacol. 2012, 166, 510–521. [Google Scholar] [CrossRef]
- Kelly, S.; Chapman, R.J.; Woodhams, S.; Sagar, D.R.; Turner, J.; Burston, J.J.; Bullock, C.; Paton, K.; Huang, J.; Wong, A.; et al. Increased function of pronociceptive TRPV1 at the level of the joint in a rat model of osteoarthritis pain. Ann. Rheum. Dis. 2015, 74, 252–259. [Google Scholar] [CrossRef]
- Yu, L.; Yang, F.; Luo, H.; Liu, F.Y.; Han, J.S.; Xing, G.G.; Wan, Y. The role of TRPV1 in different subtypes of dorsal root ganglion neurons in rat chronic inflammatory nociception induced by complete Freund’s adjuvant. Mol. Pain 2008, 4, 61. [Google Scholar] [CrossRef]
- Nummenmaa, E.; Hamalainen, M.; Moilanen, L.J.; Paukkeri, E.L.; Nieminen, R.M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Transient receptor potential ankyrin 1 (TRPA1) is functionally expressed in primary human osteoarthritic chondrocytes. Arthr. Res. Ther. 2016, 18, 185. [Google Scholar] [CrossRef] [Green Version]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhe, V.; Plee-Gautier, E.; Carre, J.L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: Pro-inflammatory response induced by their activation and their sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef]
- Sadowska, A.; Hitzl, W.; Jaszczuk, P.; Cherif, H.; Haglund, L.; Hausmann, O.; Wuertz-Kozak, K. Differential regulation of TRP channel gene expression by intervertebral disc degeneration and back pain. Sci. Rep. 2018. in revision. [Google Scholar]
- Flegel, C.; Schobel, N.; Altmuller, J.; Becker, C.; Tannapfel, A.; Hatt, H.; Gisselmann, G. RNA-Seq Analysis of Human Trigeminal and Dorsal Root Ganglia with a Focus on Chemoreceptors. PLoS ONE 2015, 10, e0128951. [Google Scholar] [CrossRef]
- Asmar, A.; Barrett-Jolley, R.; Werner, A.; Kelly, R., Jr.; Stacey, M. Membrane channel gene expression in human costal and articular chondrocytes. Organogenesis 2016, 12, 94–107. [Google Scholar] [CrossRef] [Green Version]
- Gees, M.; Alpizar, Y.A.; Boonen, B.; Sanchez, A.; Everaerts, W.; Segal, A.; Xue, F.; Janssens, A.; Owsianik, G.; Nilius, B.; et al. Mechanisms of transient receptor potential vanilloid 1 activation and sensitization by allyl isothiocyanate. Mol. Pharmacol. 2013, 84, 325–334. [Google Scholar] [CrossRef]
- Everaerts, W.; Gees, M.; Alpizar, Y.A.; Farre, R.; Leten, C.; Apetrei, A.; Dewachter, I.; van Leuven, F.; Vennekens, R.; de Ridder, D.; et al. The Capsaicin Receptor TRPV1 Is a Crucial Mediator of the Noxious Effects of Mustard Oil. Curr. Biol. 2011, 21, 316–321. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, M.Z.P.; Jordt, S.E. TRPA1: A Sensory Channel of Many Talents. In TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades; Liedtke, W.B., Heller, S., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 151–160. [Google Scholar]
- The Electrophysiology Team at Nanion Technologies GmbH, Munich. TRPA1 Activation by Allyl Isothiocyanate Recorded on the Port-a-Patch®. Available online: https://www.nanion.de/en/products/port-a-patch/137-articles/413-trpa1-trpa1-activation-by-allyl-isothiocyanate-recorded-on-the-port-a-patch.html (accessed on 8 April 2019).
- McNamara, C.R.; Mandel-Brehm, J.; Bautista, D.M.; Siemens, J.; Deranian, K.L.; Zhao, M.; Hayward, N.J.; Chong, J.A.; Julius, D.; Moran, M.M.; et al. TRPA1 mediates formalin-induced pain. Proc. Natl. Acad. Sci. USA 2007, 104, 13525–13530. [Google Scholar] [CrossRef] [Green Version]
- Horvath, A.; Tekus, V.; Boros, M.; Pozsgai, G.; Botz, B.; Borbely, E.; Szolcsanyi, J.; Pinter, E.; Helyes, Z. Transient receptor potential ankyrin 1 (TRPA1) receptor is involved in chronic arthritis: In vivo study using TRPA1-deficient mice. Arthr. Res. Ther. 2016, 18, 6. [Google Scholar] [CrossRef]
- Barton, N.J.; McQueen, D.S.; Thomson, D.; Gauldie, S.D.; Wilson, A.W.; Salter, D.M.; Chessell, I.P. Attenuation of experimental arthritis in TRPV1R knockout mice. Exp. Mol. Pathol. 2006, 81, 166–170. [Google Scholar] [CrossRef]
- Szabo, A.; Helyes, Z.; Sandor, K.; Bite, A.; Pinter, E.; Nemeth, J.; Banvolgyi, A.; Bolcskei, K.; Elekes, K.; Szolcsanyi, J. Role of transient receptor potential vanilloid 1 receptors in adjuvant-induced chronic arthritis: In vivo study using gene-deficient mice. J. Pharmacol. Exp. Ther. 2005, 314, 111–119. [Google Scholar] [CrossRef]
- Schumacher, M.A. Transient Receptor Potential Channels in Pain and Inflammation: Therapeutic Opportunities. Pain Pract. 2010, 10, 185–200. [Google Scholar] [CrossRef]
- Aoki, Y.; Nakajima, A.; Ohtori, S.; Takahashi, H.; Watanabe, F.; Sonobe, M.; Terajima, F.; Saito, M.; Takahashi, K.; Toyone, T.; et al. Increase of nerve growth factor levels in the human herniated intervertebral disc: Can annular rupture trigger discogenic back pain? Arthr. Res. Ther. 2014, 16, R159. [Google Scholar] [CrossRef]
- Chen, J.; Hackos, D.H. TRPA1 as a drug target—Promise and challenges. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 451–463. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Moriello, A.S. Modulation of the TRPV1 channel: Current clinical trials and recent patents with focus on neurological conditions. Recent Pat. CNS Drug Discov. 2013, 8, 180–204. [Google Scholar] [CrossRef]
- Botz, B.; Bolcskei, K.; Helyes, Z. Challenges to develop novel anti-inflammatory and analgesic drugs. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9, e1427. [Google Scholar] [CrossRef]
- Lehto, S.G.; Weyer, A.D.; Youngblood, B.D.; Zhang, M.; Yin, R.; Wang, W.; Teffera, Y.; Cooke, M.; Stucky, C.L.; Schenkel, L.; et al. Selective antagonism of TRPA1 produces limited efficacy in models of inflammatory- and neuropathic-induced mechanical hypersensitivity in rats. Mol. Pain 2016, 12. [Google Scholar] [CrossRef] [Green Version]
- Kun, J.; Szitter, I.; Kemeny, A.; Perkecz, A.; Kereskai, L.; Pohoczky, K.; Vincze, A.; Godi, S.; Szabo, I.; Szolcsanyi, J.; et al. Upregulation of the transient receptor potential ankyrin 1 ion channel in the inflamed human and mouse colon and its protective roles. PLoS ONE 2014, 9, e108164. [Google Scholar] [CrossRef] [PubMed]
- Kistner, K.; Siklosi, N.; Babes, A.; Khalil, M.; Selescu, T.; Zimmermann, K.; Wirtz, S.; Becker, C.; Neurath, M.F.; Reeh, P.W.; et al. Systemic desensitization through TRPA1 channels by capsazepine and mustard oil—A novel strategy against inflammation and pain. Sci. Rep. 2016, 6, 28621. [Google Scholar] [CrossRef] [PubMed]
- Cayouette, S.; Lussier, M.P.; Mathieu, E.L.; Bousquet, S.M.; Boulay, G. Exocytotic insertion of TRPC6 channel into the plasma membrane upon G(q) protein-coupled receptor activation. J. Biol. Chem. 2004, 279, 7241–7246. [Google Scholar] [CrossRef]
- Qu, Z.W.; Wang, Y.Q.; Li, X.; Wu, L.; Wang, Y.Z. TRPC6 expression in neurons is differentially regulated by NR2A-and NR2B-containing NMDA receptors. J. Neurochem. 2017, 143, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Chen, Y.Q.; Yang, K.; Wang, Y.F.; Tian, L.C.; Zhang, J.; Wang, E.W.; Sun, D.J.; Lu, W.J.; Wang, J. Chronic Hypoxia Increases TRPC6 Expression and Basal Intracellular Ca2+ Concentration in Rat Distal Pulmonary Venous Smooth Muscle. PLoS ONE 2014, 9, e112007. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, S.; Erickson, G.R.; Guilak, F. Hyperosmotically induced volume change and calcium signaling in intervertebral disk cells: The role of the actin cytoskeleton. Biophys. J. 2002, 83, 2502–2510. [Google Scholar] [CrossRef]
- Shimosato, G.; Amaya, F.; Ueda, M.; Tanaka, Y.; Decosterd, I.; Tanaka, M. Peripheral inflammation induces up-regulation of TRPV2 expression in rat DRG. Pain 2005, 119, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Neeper, M.P.; Liu, Y.; Hutchinson, T.L.; Lubin, M.L.; Flores, C.M. TRPV2 is activated by cannabidiol and mediates CGRP release in cultured rat dorsal root ganglion neurons. J. Neurosci. 2008, 28, 6231–6238. [Google Scholar] [CrossRef] [PubMed]
- Krock, E.; Rosenzweig, D.H.; Currie, J.B.; Bisson, D.G.; Ouellet, J.A.; Haglund, L. Toll-like Receptor Activation Induces Degeneration of Human Intervertebral Discs. Sci. Rep. 2017, 7, 17184. [Google Scholar] [CrossRef]
- Krupkova, O.; Sekiguchi, M.; Klasen, J.; Hausmann, O.; Konno, S.; Ferguson, S.J.; Wuertz-Kozak, K. Epigallocatechin 3-gallate suppresses interleukin-1beta-induced inflammatory responses in intervertebral disc cells in vitro and reduces radiculopathic pain in rats. Eur. Cell. Mater. 2014, 28, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Leung, V.Y.; Chan, W.C.; Hung, S.C.; Cheung, K.M.; Chan, D. Matrix remodeling during intervertebral disc growth and degeneration detected by multichromatic FAST staining. J. Histochem. Cytochem. 2009, 57, 249–256. [Google Scholar] [CrossRef]
- Boudes, M.; Uvin, P.; Pinto, S.; Freichel, M.; Birnbaumer, L.; Voets, T.; de Ridder, D.; Vennekens, R. Crucial role of TRPC1 and TRPC4 in cystitis-induced neuronal sprouting and bladder overactivity. PLoS ONE 2013, 8, e69550. [Google Scholar] [CrossRef]
- Alkhani, H.; Ase, A.R.; Grant, R.; O’Donnell, D.; Groschner, K.; Seguela, P. Contribution of TRPC3 to store-operated calcium entry and inflammatory transductions in primary nociceptors. Mol. Pain 2014, 10, 43. [Google Scholar] [CrossRef]
- Pairet, N.; Mang, S.; Fois, G.; Keck, M.; Kuhnbach, M.; Gindele, J.; Frick, M.; Dietl, P.; Lamb, D.J. TRPV4 inhibition attenuates stretch-induced inflammatory cellular responses and lung barrier dysfunction during mechanical ventilation. PLoS ONE 2018, 13, e0196055. [Google Scholar] [CrossRef]
- Hummel, D.M.; Fetahu, I.S.; Groschel, C.; Manhardt, T.; Kallay, E. Role of proinflammatory cytokines on expression of vitamin D metabolism and target genes in colon cancer cells. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt A, 91–95. [Google Scholar] [CrossRef]
- Haraguchi, K.; Kawamoto, A.; Isami, K.; Maeda, S.; Kusano, A.; Asakura, K.; Shirakawa, H.; Mori, Y.; Nakagawa, T.; Kaneko, S. TRPM2 Contributes to Inflammatory and Neuropathic Pain through the Aggravation of Pronociceptive Inflammatory Responses in Mice. J. Neurosci. 2012, 32, 3931–3941. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, R.; Hyun, E.; Zhao, L.N.; Lapointe, T.K.; Chapman, K.; Hirota, C.L.; Ghosh, S.; McKemy, D.D.; Vergnolle, N.; Beck, P.L.; et al. TRPM8 activation attenuates inflammatory responses in mouse models of colitis. Proc. Natl. Acad. Sci. USA 2013, 110, 7476–7481. [Google Scholar] [CrossRef] [Green Version]
- Wuertz, K.; Haglund, L. Inflammatory Mediators in Intervertebral Disk Degeneration and Discogenic Pain. Glob. Spine J. 2013, 3, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Brew, K.; Nagase, H. The tissue inhibitors of metalloproteinases (TIMPs): An ancient family with structural and functional diversity. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 55–71. [Google Scholar] [CrossRef] [Green Version]
- Hilario, M.O.; Terreri, M.T.; Len, C.A. Nonsteroidal anti-inflammatory drugs: Cyclooxygenase 2 inhibitors. J. Pediatr. 2006, 82 (Suppl. 5), S206–S212. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Degenerated Lumbar IVDs | n | 2−dCt (Mean ± SD) | Region | Pain score | Grade | Age (Mean ± SD) |
|---|---|---|---|---|---|---|
| TRPA1 | in 4 out of 20 | 0.0006 ± 0.001 | 2 in AF 2 in NP | 2 pain score 2; 2 pain score 3 | 2 grade 2; 2 grade 3 | 60 ± 15.6 max: 76; min: 39 |
| TRPV1 [26] | 19 out of 20 | 0.0047 ± 0.0024 | 10 in NP 9 in AF 2 in mix | 6 in pain score 1; 11 in pain score 2; 4 in pain score 3 | 4 grade 2; 8 grade 3; 6 grade 4; 3 grade 5 | 54 ± 15 max: 80; min: 31 |
| Treatment | Untreated | IL-1β 5 ng/mL | IL-1β 10 ng/mL | TNF-α 5 ng/mL | TNF-α 10 ng/mL |
|---|---|---|---|---|---|
| TRPA1 | No expression | 385.18 ± 233.06 | 333.42 ± 199.80 | 842.06 ± 659.98 | 780.23 ± 600.17 |
| TRPV1 | Expression | 0.64 ± 0.18 | 0.69 ± 0.23 | 0.53 ± 0.11 | 0.45 ± 0.22 |
| Tissue | |||||||
| Donor | Age | Gender | Pathology | Tissue | Level | Grade | Experiments |
| T1 | 30 | m | DDD | AF, NP | L4/5 | II | qPCR |
| T2 | 46 | f | DH | AF, NP | L5/S1 | III | qPCR |
| T3 | 34 | m | DH | AF, NP | L5/S1 | III | qPCR |
| T4 | 46 | f | DH | AF, NP | L4-S1 | V | qPCR |
| T5 | 59 | f | DDD | AF, NP | L5/S1 | V | qPCR |
| T6 | 62 | f | DDD | AF | L5/S1 | V | qPCR |
| T7 | 66 | f | DH | AF | L4/5 | II | qPCR |
| T8 | 53 | m | DH | NP | L5/S1 | II | qPCR |
| T9 | 59 | m | DH | NP | L4/L5 | II | qPCR |
| T10 | 52 | m | DDD | NP | L4/L5 | III | qPCR |
| T11 | 64 | f | DDD | NP | L4/L5 | IV | qPCR |
| T12 | 76 | f | DH | NP | L4/L5 | III | qPCR |
| T13 | 16 | f | DH | NP | L4/L5 | III | qPCR |
| T14 | 31 | m | DDD | AF | L4/5 L5/S1 | IV | qPCR |
| T15 | 54 | f | DH | NP | L5/S1 | II | qPCR |
| T16 | 33 | m | DH | AF | L5/S1 | II | qPCR |
| T17 | 70 | f | DDD | NP | L4/5 | IV | qPCR |
| T18 | 39 | m | DH | AF | L5/S1 | III | qPCR |
| T19 | 28 | m | DH | NP | L5/S1 | II | qPCR |
| T20 | 21 | f | DDD | AF | L4/5 | III | qPCR |
| Cells | |||||||
| Donor | Age | Gender | Pathology | Tissue | Level | Grade | Experiments |
| C1 | 44 | F | uk | uk | L3/L4 | uk | qPCR |
| C2 | 82 | M | uk | uk | L5/S1 | uk | qPCR |
| C3 | 28 | M | uk | uk | L5/S1 | uk | qPCR |
| C4 | uk | uk | uk | uk | uk | uk | qPCR, ELISA |
| C5 | uk | uk | uk | uk | uk | uk | qPCR, ELISA |
| C6 | 39 | M | DDD, DH | Mix | L4/L5 | IV | ELISA |
| C7 | 58 | M | DDD, DH | Mix | L5/S1 | IV | qPCR, ELISA |
| C8 | 46 | F | DH | - | L5/S1 | IV | qPCR, ELISA |
| C9 | 52 | M | DDD, DH | Mix | L5/S1 | V | qPCR, ELISA |
| C10 | 46 | F | DDD, DH | Mix | L4/L5 | IV | qPCR, Ca imaging |
| C11 | 58 | M | uk | Mix | L4/L5 | IV | Ca imaging |
| C12 | 31 | M | DDD, DH | Mix | L5/S1 | IV | qPCR |
| C13 | 46 | M | DH | Mix | C5/C6 | III | qPCR |
| C14 | 40 | F | DH | Mix | L4/L5 | III | qPCR |
| C15 | 40 | M | DH | NP | L4/L5 | III | qPCR |
| C16 | 66 | F | DH | NP | L4/L5 | III | Ca imaging |
| C17 | uk | uk | uk | uk | uk | uk | qPCR, ELISA |
| C18 | 50 | F | DH | Mix | L5/S1 | IV | qPCR |
| C19 | 42 | M | DH | Mix | L5/S1 | V | qPCR |
| C20 | 68 | F | listhesis | Mix | L4/L5 | III | qPCR |
| C21 | 38 | M | DH | Mix | L5/S1 | III | qPCR, ELISA |
| C22 | 41 | F | DH | Mix | L4/5 | III | qPCR, ELISA |
| C23 | 42 | M | DH | NP | L5/S1 | IV | qPCR, ELISA |
| C24 | 41 | F | DH | Mix | L5/S1 | III | qPCR, ELISA |
| C25 | 45 | M | DH | NP | L4/L5 | IV | qPCR, ELISA |
| C26 | 71 | uk | DDD | Mix | L4/5 | III | qPCR, ELISA |
| C27 | 55 | F | DH | NP | L5/6 | I | qPCR, ELISA |
| C28 | 55 | F | DH | NP | L5/6 | II | Immuno |
| C29 | 55 | M | DH | Mix | L5/S1 | II | Immuno |
| C30 | 58 | M | DH | Mix | L4/5 | IV | Immuno |
| C31 | 34 | M | - | NP, AF | L1/2-L2/3-L3/4 | I | qPCR |
| C32 | 55 | F | - | NP, AF | L1/2 | III | qPCR |
| C33 | 52 | M | - | NP | L1-L5 | I | qPCR |
| C34 | 17 | M | - | NP | T12-S1 | I | qPCR |
| LC1 | 16 | M | - | mix | uk | II | qPCR |
| LFC2 | Fetal IVD cells, Male, 16 weeks: p5 | qPCR | |||||
| LFC3 | Fetal IVD cells, Male, 14 weeks: p4 | qPCR | |||||
| LFC4 | Fetal IVD cells, Male, 14 weeks: p4 | qPCR | |||||
| Mice | 2 Months Old | 4 Months Old | 7 Months Old |
|---|---|---|---|
| TRPA1 WT | n = 5 | - | n = 5 |
| TRPA1 KO | n = 5 | - | n = 5 |
| TRPV1 KO | - | n = 5 | n = 5 |
| C57BL/6 | - | n = 5 | - |
| Compound | Catalog Number | Function | Concentration | Exposure Time | Experiment |
|---|---|---|---|---|---|
| TNF-α | 315-01A PeproTech | Inflammatory cytokine | 5, 10 ng/mL | 18 h | qPCR Ca imaging |
| IL-1β | 200-01B PeproTech | Inflammatory cytokine | 5, 10 ng/mL | 18 h | qPCR Ca imaging |
| 5 ng/mL | 1, 8, 15 days | qPCR (from 3D) | |||
| 5 ng/mL | 18 h | Immuno | |||
| AITC | 377430 Sigma | TRPA1 agonist | 3 μM, 10 μM | 18 h | qPCR, ELISA |
| 100 μM | during the measurement | Ca imaging |
| Target Gene | Assay ID | Putative Association with Inflammation |
|---|---|---|
| TBP | Hs00427620_m1 | HKG in the cell culture study and fetal cells |
| GAPDH | Hs02758991_g1 | HKG in the tissue study |
| TRPA1 | Hs00175798_m1 | inflammatory pain [12] |
| TRPC1 | Hs00608195_m1 | bladder inflammation (neuronal) [54] |
| TRPC3 | Hs00162985_m1 | inflammatory pain [55] |
| TRPC6 | Hs00988479_m1 | IVD inflammation (putative) [18,19] |
| TRPV1 | Hs00218912_m1 | neuroinflammation [20] |
| TRPV2 | Hs00901648_m1 | inflammation in DRG [49] |
| TRPV4 | Hs01099348_m1 | lung inflammation [56] |
| TRPV6 | Hs00367960_m1 | association with TNF-α [57] |
| TRPM2 | Hs01066091_m1 | inflammatory and neuropathic pain [58] |
| TRPM7 | Hs00559080_m1 | inflammation in colitis [59] |
| IL-6 | Hs00174131_m1 | inflammation mediator [60] |
| IL-8 | Hs00174103_m1 | inflammation mediator [60] |
| MMP1 | Hs00233958_m1 | cleaves mainly collagens (I, II, III) [61] |
| MMP3 | Hs00968305_m1 | cleaves proteoglycans and collagens (II, III) [61] |
| ADAMTS4 | Hs00192708_m1 | cleaves mainly aggrecan [61] |
| ADAMTS5 | Hs01095518_m1 | cleaves mainly aggrecan [61] |
| TIMP1 | Hs00234278_m1 | inhibits MMPs (1, 3) and ADAMTS (4) [61] |
| TIMP2 | Hs01092512_g1 | inhibits all MMPs [61] |
| COX-2 | Hs00153133_m1 | pain mediator [62] |
| NGF | Hs00171458_m1 | nerve ingrowth [38] |
| COL2A1 | Hs00264051_m1 | ECM constituent |
| COL1A1 | Hs00164004_m1 | ECM constituent |
| Aggrecan | Hs00153936_m1 | ECM constituent |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kameda, T.; Zvick, J.; Vuk, M.; Sadowska, A.; Tam, W.K.; Leung, V.Y.; Bölcskei, K.; Helyes, Z.; Applegate, L.A.; Hausmann, O.N.; et al. Expression and Activity of TRPA1 and TRPV1 in the Intervertebral Disc: Association with Inflammation and Matrix Remodeling. Int. J. Mol. Sci. 2019, 20, 1767. https://doi.org/10.3390/ijms20071767
Kameda T, Zvick J, Vuk M, Sadowska A, Tam WK, Leung VY, Bölcskei K, Helyes Z, Applegate LA, Hausmann ON, et al. Expression and Activity of TRPA1 and TRPV1 in the Intervertebral Disc: Association with Inflammation and Matrix Remodeling. International Journal of Molecular Sciences. 2019; 20(7):1767. https://doi.org/10.3390/ijms20071767
Chicago/Turabian StyleKameda, Takuya, Joel Zvick, Miriam Vuk, Aleksandra Sadowska, Wai Kit Tam, Victor Y. Leung, Kata Bölcskei, Zsuzsanna Helyes, Lee Ann Applegate, Oliver N. Hausmann, and et al. 2019. "Expression and Activity of TRPA1 and TRPV1 in the Intervertebral Disc: Association with Inflammation and Matrix Remodeling" International Journal of Molecular Sciences 20, no. 7: 1767. https://doi.org/10.3390/ijms20071767