Biochemical and Anatomical Investigation of Sesbania herbacea (Mill.) McVaugh Nodules Grown under Flooded and Non-Flooded Conditions

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Sesbania Plants Grown in Flooded Fields Produce More Biomass
2.2. Flooding Increases the Number of Nodules on Sesbania Roots
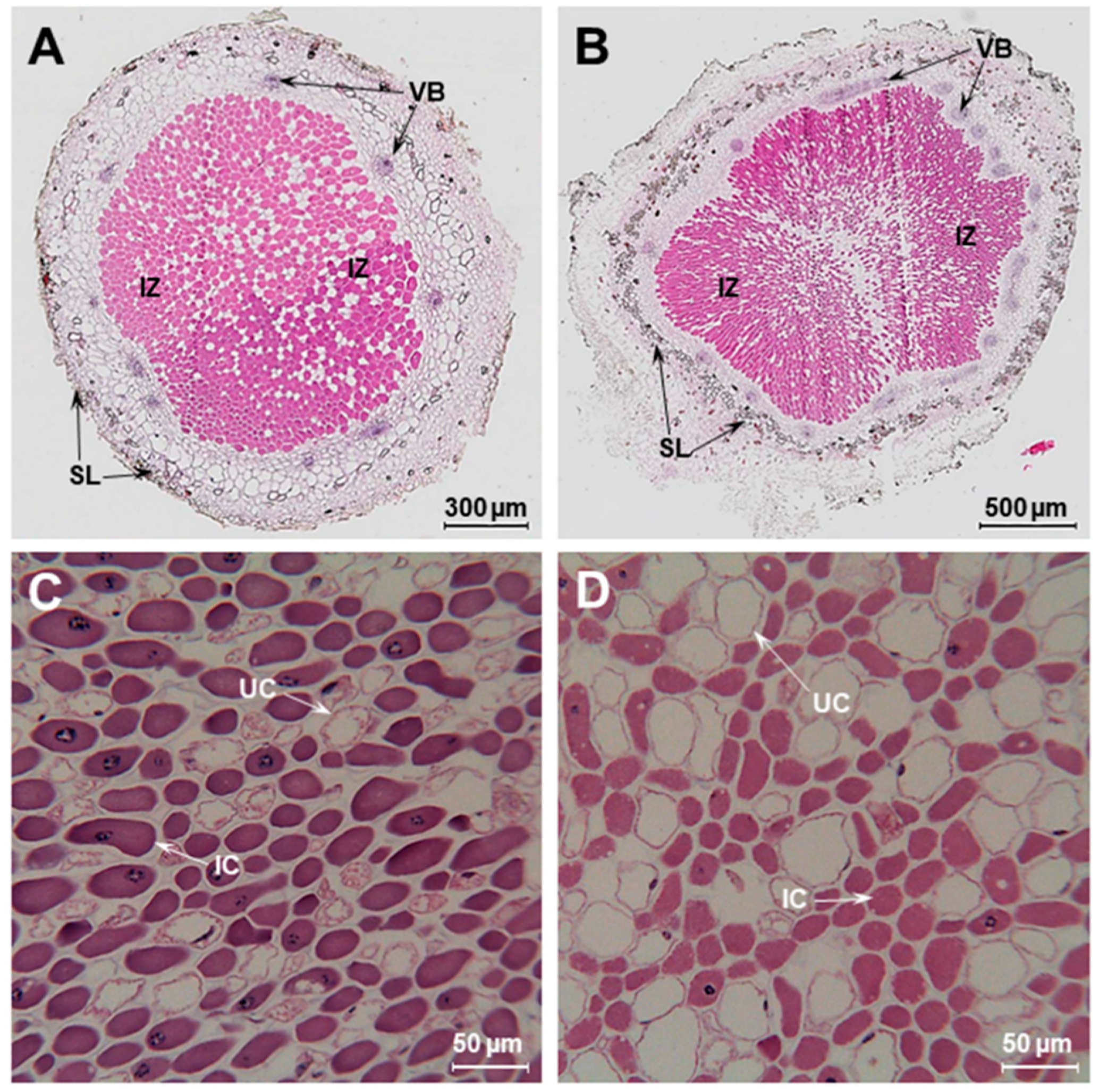
2.3. Comparison of Nodule Anatomy of Sesbania Nodules Grown under Flooded and Non-Flooded Conditions
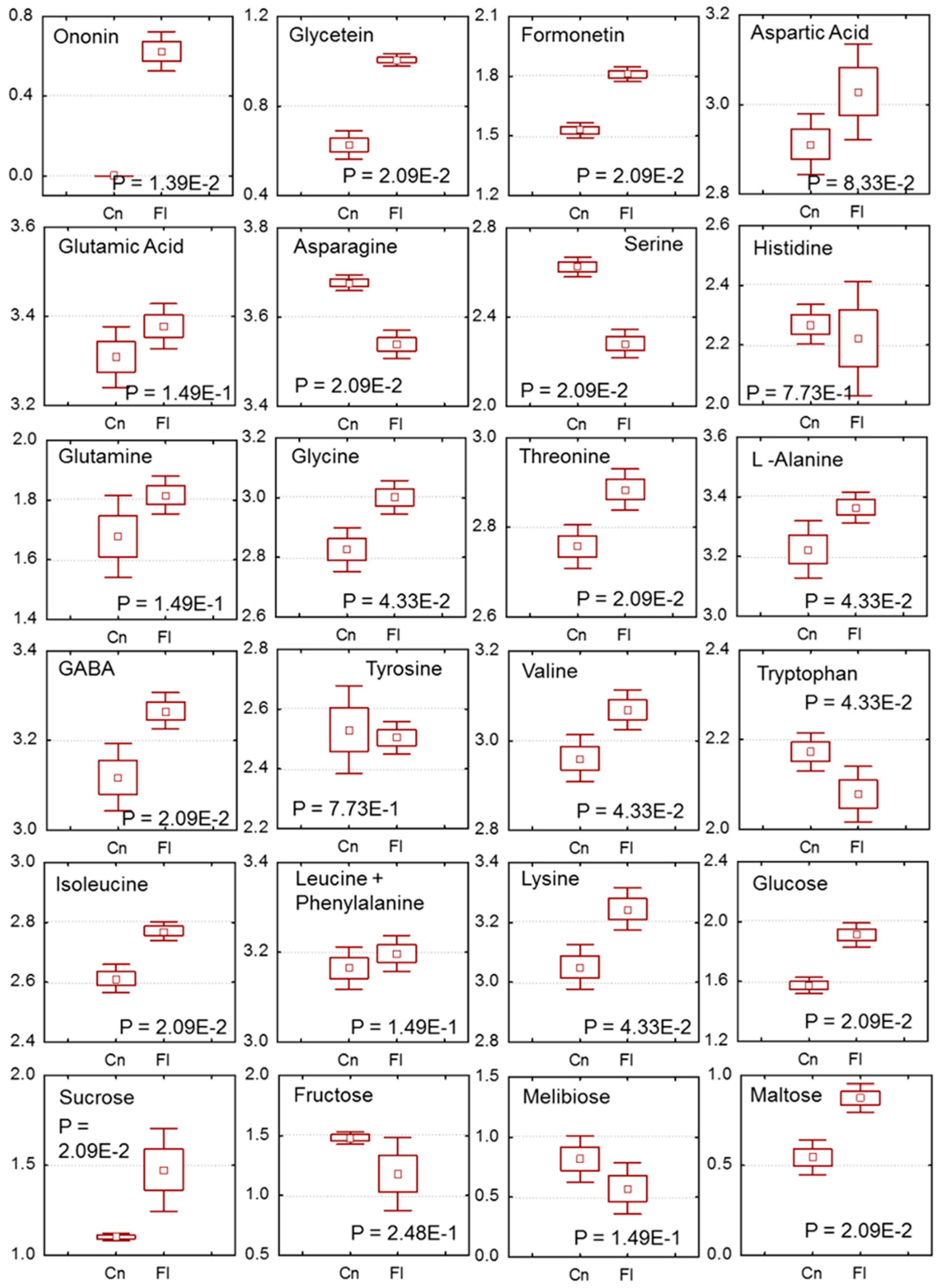
2.4. Changes in Sesbania Nodule Metabolites Due to Flooding
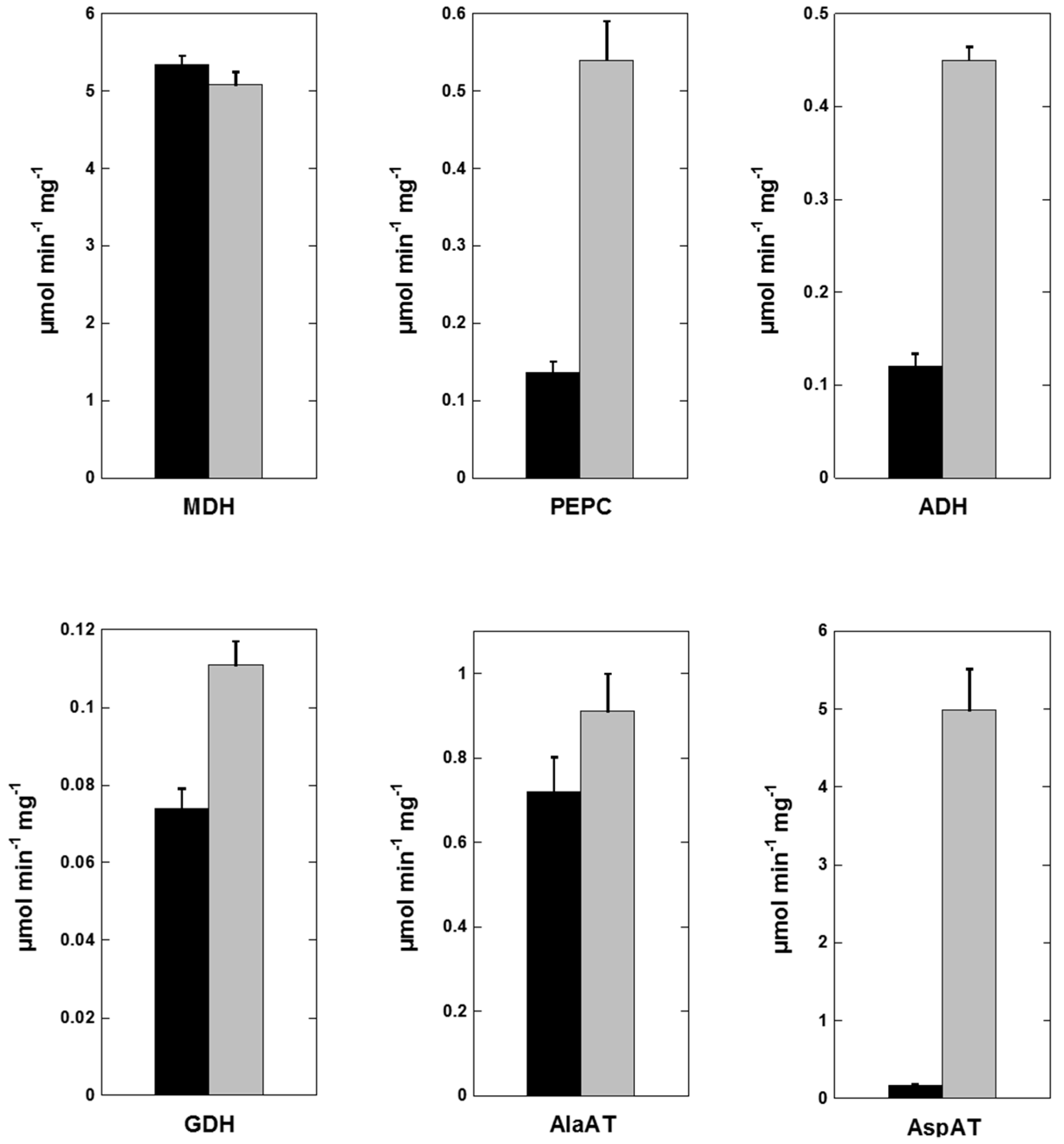
2.5. Flooding-Induced Changes in Enzyme Activities
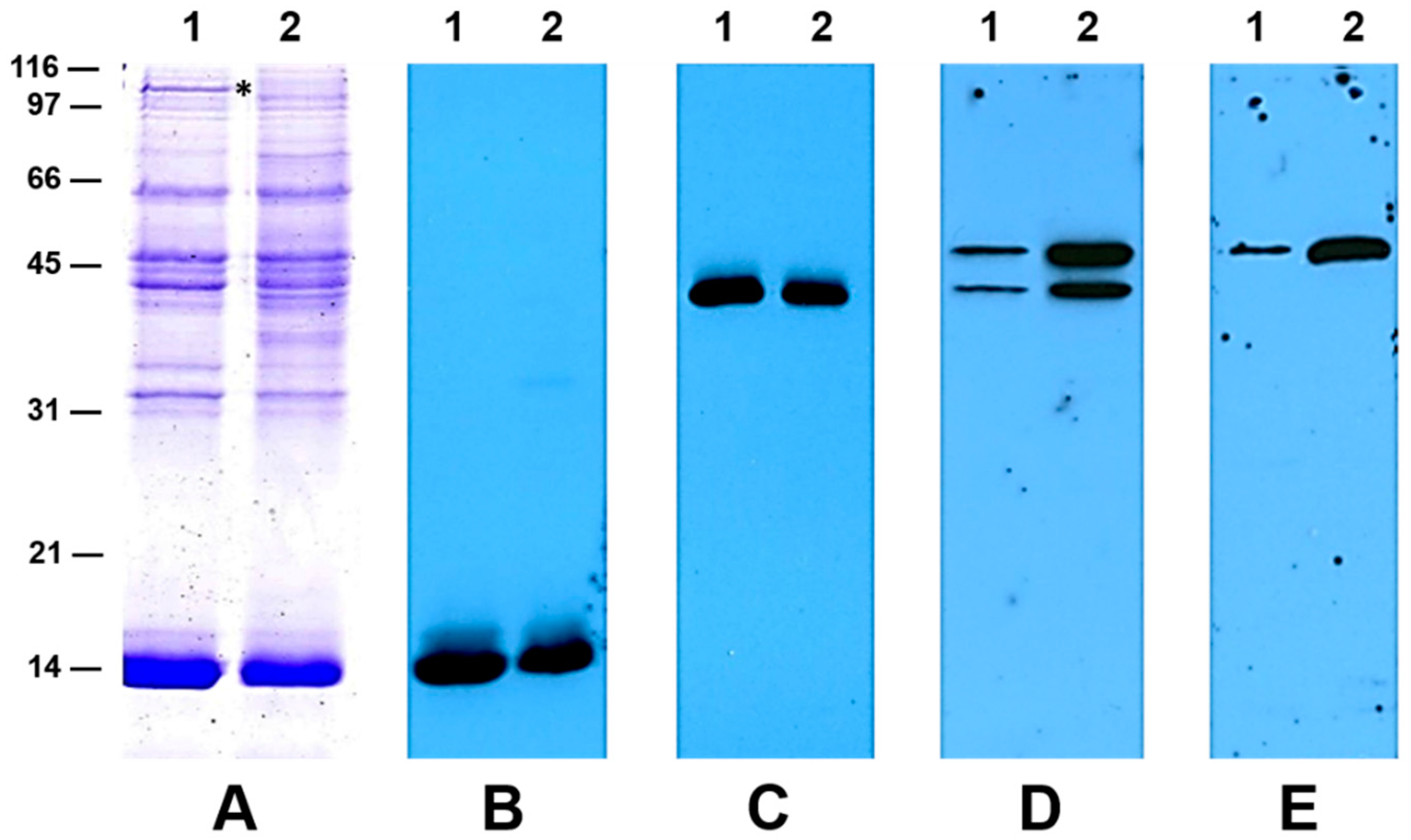
2.6. Western Blot Analysis Reveals a Higher Accumulation of Aspartate Aminotransferase in Flooded Nodules
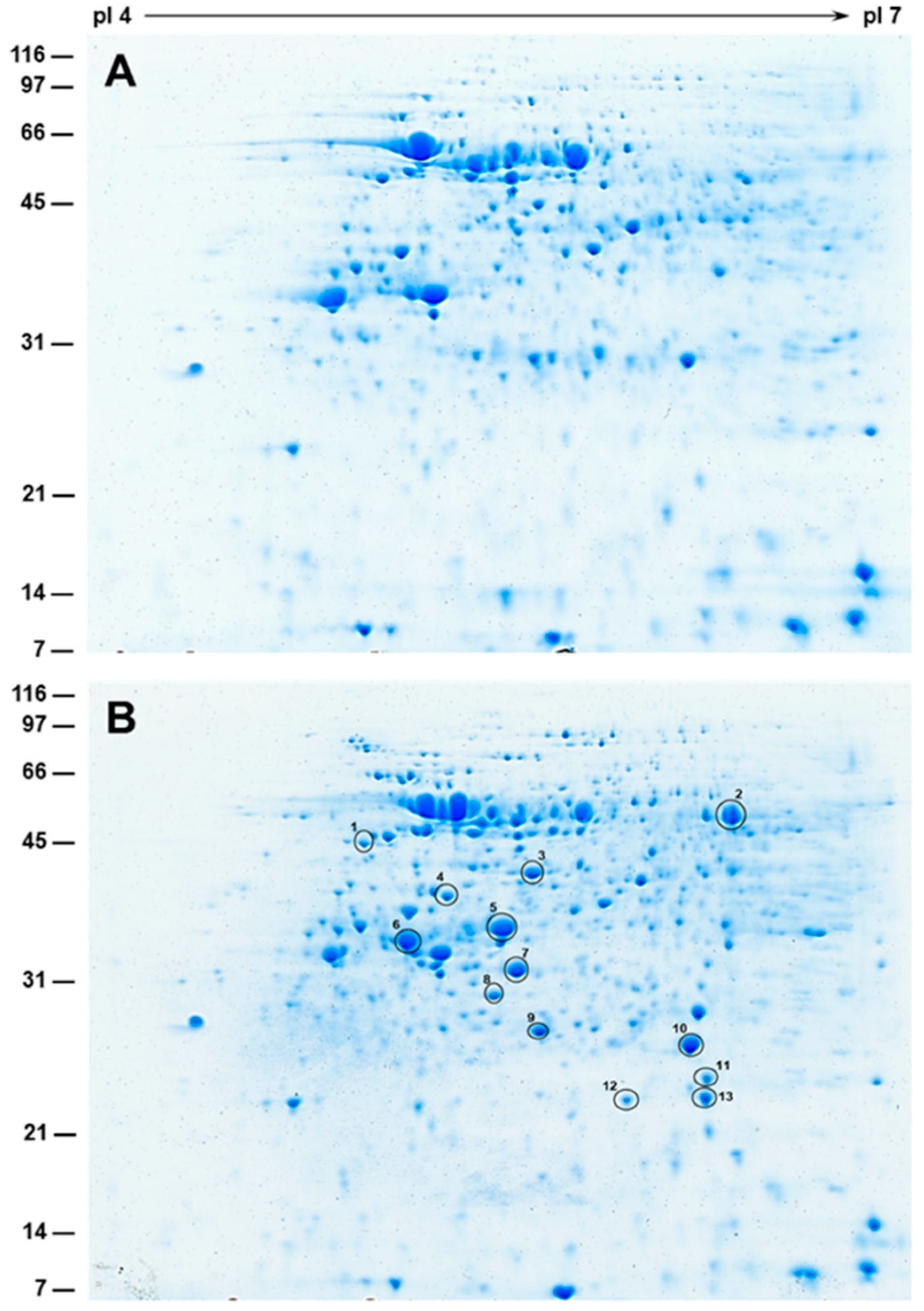
2.7. 2-D SDS-PAGE and Protein Identification by Mass Spectrometry
3. Discussion
4. Materials and Methods
4.1. Greenhouse Experiments
4.2. Embedment of Sesbania Nodules in Paraffin
4.3. Light and Transmission Electron Microscopy
4.4. Protein Isolation, 1-D SDS-PAGE, and Immunoblot Analysis
4.5. Protein Isolation, 2-D SDS-PAGE and Image Analysis
4.6. MALDI-TOF/MS
4.7. Extraction of Secondary Metabolites
4.8. Amino Acids Extraction
4.9. Soluble Sugars Extraction
4.10. LC-MS Analysis for Secondary Metabolites
4.11. LC-MS Analysis for Amino Acids
4.12. IC-MS Analysis for Sugarslog10
4.13. Data Processing and Multivariate Analysis
4.14. Enzyme Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 1-D | One-dimensional |
| 2-D | Two-dimensional |
| NH3 | Ammonia |
| ATP | Adenosine triphosphate |
| MDH | Malate dehydrogenase |
| ADH | Alcohol dehydrogenase |
| GDH | Glutamate dehydrogenase |
| PEPC | Phosphenolpyruvate carboxylase |
| AspAT | Aspartate aminotransferase |
| AlaAT | Alanine aminotransferase |
| GABA | Gamma-Amino Butyric acid |
| LC-MS | Liquid chromatography–mass spectrometry |
| IC-MS | Ion chromatography–mass spectrometry |
| MALDI-TOF/MS | Matrix-Assisted Laser Desorption Ionization-Time of Flight/MS |
| TEM | Transmission electron microscopy |
| PBM | Peribacteroid membrane |
| DAI | Days after inoculation |
| PHB | Poly β-hydroxybutyrate |
| EPS | Exopolysaccharides |
| LPS | Lipopolysaccharides |
References
- Sheahan, C.M. Plant Guide for Bigpod Sesbania (Sesbania exaltata); Cape May Plant Materials Center: Cape May, NJ, USA, 2013. [Google Scholar]
- Bryson, C.T. Interference and critical time of removal of hemp sesbania (Sesbania exaltata) in cotton (Gossypium hirsutum). Weed Technol. 1990, 4, 833–837. [Google Scholar] [CrossRef]
- McWhorter, C.; Anderson, J. Hemp sesbania (Sesbania exaltata) competition in soybeans (Glycine max). Weed Sci. 1979, 27, 58–64. [Google Scholar]
- King, C.A.; Purcell, L.C. Interference between hemp sesbania (Sesbania exaltata) and soybean (Glycine max) in response to irrigation and nitrogen. Weed Sci. 1997, 45, 91–97. [Google Scholar]
- Norsworthy, J.K.; Oliver, L.R. Hemp sesbania interference in drill-seeded glyphosate-resistant soybean. Weed Sci. 2002, 50, 34–41. [Google Scholar]
- Evans, D.O.; Rotar, P.P. Sesbania in Agriculture; Westview Press: Boulder, CO, USA, 1987. [Google Scholar]
- Summer Cover Crop Use in Arizona Vegetable Production Systems; College of Agriculture and Life Sciences, University of Arizona: Tucson, AZ, USA, 2010.
- Lovelace, M.L.; Oliver, L.R. Effects of interference and tillage on hemp sesbania and pitted morningglory emergence, growth, and seed production. In Proceedings of the Annual Meeting of Southern Weed Science Society 2000, Tulsa, OK, USA, 24–26 January 2000; p. 202. [Google Scholar]
- Handbook of Energy Crops; Center for New Crops and Plants Products, Purdue University: Lafeyette, IN, USA, 1983.
- Allen, O.N.; Allen, E.K. The Leguminosae, a Source Book of Characteristics, Uses, and Nodulation; University of Wisconsin Press: Madison, WI, USA 1981. [Google Scholar]
- Loureiro, M.F.; James, E.K.; Franco, A.A. Nitrogen fixation by legumes in flooded regions. Oecol. Bras. 1998, 4, 191–219. [Google Scholar] [CrossRef]
- Wang, E.T.; Martinez-Romero, E. Sesbania herbacea–Rhizobium huautlense nodulation in flooded soils and comparative characterization of S. herbacea-nodulating Rhizobia in different environments. Microb. Ecol. 2000, 40, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Loveday, J. Influence of oxygen diffusion rate on nodulation of subterranean clover. Aust. J. Sci. 1963, 26, 90–91. [Google Scholar]
- Minchin, F.R.; Pate, J.S. Effects of water, aeration, and salt regime on nitrogen fixation in a nodulated legume—definition of an optimum root environment. J. Exp. Bot. 1975, 26, 60–69. [Google Scholar] [CrossRef]
- Eaglesham, A.R.J.; Ayanaba, A. Tropical stress ecology of rhizobia, root nodulation and legume fixation. In Current Developments in Biological Nitrogen Fixation; Subba Rao, N.S., Ed.; Edward Arnold Ltd.: London, UK, 1984; pp. 1–35. [Google Scholar]
- De Faria, S.M.; Lewis, G.P.; Sprent, J.I.; Sutherland, J.M. Occurrence of nodulation in the Leguminosae. New Phytol. 1989, 111, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Dreyfus, B.; Dommergues, Y.R. Nitrogen-fixing nodules induced by Rhizobium on the stem of the tropical legume Sesbania rostrata. FEMS Microbiol. Lett. 1981, 10, 313–317. [Google Scholar] [CrossRef]
- Dreyfus, B.; Garcia, J.L.; Gillis, M. Characterization of Azorhizobium caulinodans gen. nov., sp. nov., a stem-nodulating nitrogen-fixing bacterium isolated from Sesbania rostrata. Int. J. Syst. Evol. Microbiol. 1988, 38, 89–98. [Google Scholar] [CrossRef]
- Capoen, W.; Oldroyd, G.; Goormachtig, S.; Holsters, M. Sesbania rostrata: A case study of natural variation in legume nodulation. New Phytol. 2010, 186, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Van Berkum, P.; Beyene, D.; Sui, X.H.; Dorado, O.; Chen, W.X.; Martínez-Romero, E. Rhizobium huautlense sp. nov., a symbiont of Sesbania herbacea that has a close phylogenetic relationship with Rhizobium galegae. Int. J. Syst. Evol. Microbiol. 1998, 48, 687–699. [Google Scholar] [CrossRef]
- Rana, D.; Krishnan, H.B. A new root-nodulating symbiont of the tropical legume Sesbania, Rhizobium sp SIN-1, is closely related to R. galegae, a species that nodulates temperate legumes. FEMS Microbiol. Lett. 1995, 134, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.B.; Pueppke, S.G. Ineffective nodulation of Sesbania macrocarpa by Sinorhizobium meliloti strain RCR2011. FEMS Microbiol. Lett. 1998, 165, 207–214. [Google Scholar] [CrossRef]
- Cummings, S.P.; Gyaneshwar, P.; Vinuesa, P.; Farruggia, F.T.; Andrews, M.; Humphry, D.; Elliott, G.N.; Nelson, A.; Orr, C.; Pettitt, D.; et al. Nodulation of Sesbania species by Rhizobium (Agrobacterium) strain IRBG74 and other rhizobia. Environ. Microbiol. 2009, 11, 2510–2525. [Google Scholar] [CrossRef]
- Mousavi, S.A.; Österman, J.; Wahlberg, N.; Nesme, X.; Lavire, C.; Vial, L.; Paulin, L.; De Lajudie, P.; Lindström, K. Phylogeny of the Rhizobium–Allorhizobium–Agrobacterium clade supports the delineation of Neorhizobium gen. nov. Syst. Appl. Microbiol. 2014, 37, 208–215. [Google Scholar] [CrossRef]
- Armstrong, W. Aeration in higher plants. In Advances in Botanical Research; Elsevier: Cambridge, MA, USA, 1980; Volume 7, pp. 225–332. [Google Scholar]
- Colmer, T. Aerenchyma and an inducible barrier to radial oxygen loss facilitate root aeration in upland, paddy and deep-water rice (Oryza sativa L.). Ann. Bot. 2002, 91, 301–309. [Google Scholar] [CrossRef]
- James, E.K.; Sprent, J.I.; Sutherland, J.M.; McInroy, S.G.; Minchin, F.R. The structure of nitrogen fixing root nodules on the aquatic mimosoid legume Neptunia plena. Ann. Bot. 1992, 69, 173–180. [Google Scholar] [CrossRef]
- Subba-Rao, N.S.; Mateos, P.F.; Baker, D.; Pankratz, H.S.; Palma, J.; Dazzo, F.B.; Sprent, J.I. The unique root-nodule symbiosis between Rhizobium and the aquatic legume, Neptunia natans (L. f.) Druce. Planta 1995, 196, 311–320. [Google Scholar] [CrossRef]
- Loureiro, M.F.; de Faria, S.M.; James, E.K.; Pott, A.; Franco, A.A. Nitrogen-fixing stem nodules of the legume, Discolobium pulchellum Benth. New Phytol. 1994, 128, 283–295. [Google Scholar] [CrossRef]
- James, E.K.; de Fatima, L.M.; Pott, A.; Pott, V.J.; Martins, C.M.; Franco, A.A.; Sprent, J.I. Flooding-tolerant legume symbioses from the Brazilian Pantanal. New Phytol. 2001, 150, 723–738. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, M.F.; James, E.K.; Sprent, J.I.; Franco, A.A. Stem and root nodules on the tropical wetland legume Aeschynomene fluminensis. New Phytol. 1995, 130, 531–544. [Google Scholar] [CrossRef]
- James, E.K.; Crawford, R.M.M. Effect of oxygen availability on nitrogen fixation by two Lotus species under flooded conditions. J. Exp. Bot. 1998, 49, 599–609. [Google Scholar] [CrossRef]
- James, E.K.; Sprent, J.I. Development of N2-fixing nodules on the wetland legume Lotus uliginosus exposed to conditions of flooding. New Phytologist 1999, 142, 219–231. [Google Scholar] [CrossRef]
- Pugh, R.; Witty, J.F.; Mytton, L.R.; Minchin, F.R. The effect of waterlogging on nitrogen fixation and nodule morphology in soil-grown white clover (Trifolium repens L.). J. Exp. Bot. 1995, 46, 285–290. [Google Scholar] [CrossRef]
- Striker, G.G.; Colmer, T.D. Flooding tolerance of forage legumes. J. Exp. Bot. 2016, 68, 1851–1872. [Google Scholar] [CrossRef]
- Konnerup, D.; Toro, G.; Pedersen, O.; Colmer, T.D. Waterlogging tolerance, tissue nitrogen and oxygen transport in the forage legume Melilotus siculus: A comparison of nodulated and nitrate-fed plants. Ann. Bot. 2018, 121, 699–709. [Google Scholar] [CrossRef]
- Trainer, M.A.; Charles, T.C. The role of PHB metabolism in the symbiosis of rhizobia with legumes. Appl. Microbiol. Biotechnol. 2006, 71, 377–386. [Google Scholar] [CrossRef]
- Ratcliff, W.C.; Kadam, S.V.; Denison, R.F. Poly-3-hydroxybutyrate (PHB) supports survival and reproduction in starving rhizobia. FEMS Microbiol. Ecol. 2008, 65, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Quelas, J.I.; Mesa, S.; Mongiardini, E.J.; Jendrossek, D.; Lodeiro, A.R. Regulation of polyhydroxybutyrate synthesis in the soil bacterium Bradyrhizobium diazoefficiens. Appl. Environ. Microbiol. 2016, 82, 4299–4308. [Google Scholar] [CrossRef]
- Puiatti, M.; Sodek, L. Waterlogging affects nitrogen transport in the xylem of soybean. Plant Physiol. Biochem. 1999, 37, 767–773. [Google Scholar] [CrossRef]
- Oliveira, H.C.; Freschi, L.; Sodek, L. Nitrogen metabolism and translocation in soybean plants subjected to root oxygen deficiency. Plant Physiol. Biochem. 2013, 66, 141–149. [Google Scholar] [CrossRef]
- Borella, J.; Amarante, L. d.; Oliveira, D.S.; Oliveira, A.C.; Braga, E.J. Waterlogging-induced changes in fermentative metabolism in roots and nodules of soybean genotypes. Sci. Agric. 2014, 71, 499–508. [Google Scholar] [CrossRef] [Green Version]
- De Sousa, C.; Sodek, L. Alanine metabolism and alanine aminotransferase activity in soybean (Glycine max) during hypoxia of the root system and subsequent return to normoxia. Environ. Exp. Bot. 2003, 50, 1–8. [Google Scholar] [CrossRef]
- Rocha, M.; Licausi, F.; Araujo, W.L.; Nunes-Nesi, A.; Sodek, L.; Fernie, A.R.; van Dongen, J.T. Glycolysis and the tricarboxylic acid cycle are linked by alanine aminotransferase during hypoxia induced by waterlogging of Lotus japonicus. Plant Physiol. 2010, 152, 1501–1513. [Google Scholar] [CrossRef]
- Driscoll, B.T.; Finan, T.M. NAD+-dependent malic enzyme of Rhizobium meliloti is required for symbiotic nitrogen fixation. Mol. Microbiol. 1993, 7, 865–873. [Google Scholar] [CrossRef]
- Schulze, J.; Tesfaye, M.; Litjens, R.; Bucciarelli, B.; Trepp, G.; Miller, S.; Samac, D.; Allan, D.; Vance, C. Malate plays a central role in plant nutrition. Plant Soil 2002, 247, 133–139. [Google Scholar] [CrossRef]
- Rawsthorne, S.; Minchin, F.R.; Summerfield, R.J.; Cookson, C.; Coombs, J. Carbon and nitrogen metabolism in legume root nodules. Phytochemistry 1980, 19, 341–355. [Google Scholar] [CrossRef]
- Day, D. Carbon metabolism and compartmentation in nitrogen fixing legume nodules. Plant Physiol. Biochem. 1991, 29, 185–201. [Google Scholar]
- Miyashita, Y.; Dolferus, R.; Ismond, K.P.; Good, A.G. Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana. Plant. J. 2007, 49, 1108–1121. [Google Scholar] [CrossRef]
- De la Torre, F.; Cañas, R.A.; Pascual, M.B.; Avila, C.; Cánovas, F.M. Plastidic aspartate aminotransferases and the biosynthesis of essential amino acids in plants. J. Exp. Bot. 2014, 65, 5527–5534. [Google Scholar] [CrossRef] [Green Version]
- Farnham, M.W.; Miller, S.S.; Griffith, S.M.; Vance, C.P. Aspartate aminotransferase in alfalfa root nodules: II. Immunological distinction between two forms of the enzyme. Plant Physiol. 1990, 93, 603–610. [Google Scholar] [CrossRef]
- Lodwig, E.M.; Hosie, A.H.; Bourdes, A.; Findlay, K.; Allaway, D.; Karunakaran, R.; Downie, J.; Poole, P.S. Amino-acid cycling drives nitrogen fixation in the legume–Rhizobium symbiosis. Nature 2003, 422, 722–726. [Google Scholar] [CrossRef]
- Sprent, J.I.; Sprent, P. Nitrogen Fixing Organisms: Pure and Applied Aspects; Chapman & Hall: London, UK, 1990. [Google Scholar]
- Ladha, J.; Pareek, R.; Becker, M. Stem-nodulating legume-Rhizobium symbiosis and its agronomic use in lowland rice. In Advances in Soil Science; Springer: New York, NY, USA, 1992; pp. 147–192. [Google Scholar]
- Jackson, M.B. Ethylene-promoted elongation: An adaptation to submergence stress. Ann. Bot. 2007, 101, 229–248. [Google Scholar] [CrossRef]
- Lorbiecke, R.; Sauter, M. Adventitious root growth and cell-cycle induction in deepwater rice. Plant Physiol. 1999, 119, 21–30. [Google Scholar] [CrossRef]
- Lee, K.H.; LaRue, T.A. Exogenous ethylene inhibits nodulation of Pisum sativum L. cv Sparkle. Plant Physiol. 1992, 100, 1759–1763. [Google Scholar] [CrossRef]
- Fernández-López, M.; Goormachtig, S.; Gao, M.; D’Haeze, W.; Van Montagu, M.; Holsters, M. Ethylene-mediated phenotypic plasticity in root nodule development on Sesbania rostrata. Proc. Natl. Acad. Sci. USA 1998, 95, 12724–12728. [Google Scholar]
- Xie, Z.P.; Staehelin, C.; Wiemken, A.; Boller, T. Ethylene responsiveness of soybean cultivars characterized by leaf senescence, chitinase induction and nodulation. J. Plant Physiol. 1996, 149, 690–694. [Google Scholar] [CrossRef]
- D’Haeze, W.; De Rycke, R.; Mathis, R.; Goormachtig, S.; Pagnotta, S.; Verplancke, C.; Capoen, W.; Holsters, M. Reactive oxygen species and ethylene play a positive role in lateral root base nodulation of a semiaquatic legume. Proc. Natl. Acad. Sci. USA 2003, 100, 11789–11794. [Google Scholar] [Green Version]
- Bomfeti, C.; Ferreira, P.; Carvalho, T.; De Rycke, R.; Moreira, F.; Goormachtig, S.; Holsters, M. Nodule development on the tropical legume Sesbania virgata under flooded and non-flooded conditions. Plant Biol. 2013, 15, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Mukherjee, A.; Wiley-Kalil, A.; Das, S.; Owen, H.; Reddy, P.M.; Ané, J.M.; James, E.K.; Gyaneshwar, P. A rhamnose-deficient lipopolysaccharide mutant of Rhizobium sp. IRBG74 is defective in root colonization and beneficial interactions with its flooding-tolerant hosts Sesbania cannabina and wetland rice. J. Exp. Bot. 2016, 67, 5869–5884. [Google Scholar] [CrossRef]
- Colmer, T.; Voesenek, L. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef]
- Souza, S.C.; Mazzafera, P.; Sodek, L. Flooding of the root system in soybean: Biochemical and molecular aspects of N metabolism in the nodule during stress and recovery. Amino Acids 2016, 48, 1285–1295. [Google Scholar] [CrossRef]
- Narsai, R.; Rocha, M.; Geigenberger, P.; Whelan, J.; van Dongen, J.T. Comparative analysis between plant species of transcriptional and metabolic responses to hypoxia. New Phytol. 2011, 190, 472–487. [Google Scholar] [CrossRef] [Green Version]
- Buchner, P.; Takahashi, H.; Hawkesford, M.J. Plant sulphate transporters: Co-ordination of uptake, intracellular and long-distance transport. J. Exp. Bot. 2004, 55, 1765–1773. [Google Scholar] [CrossRef]
- Good, A.G.; Crosby, W.L. Anaerobic induction of alanine aminotransferase in barley root tissue. Plant Physiol. 1989, 90, 1305–1309. [Google Scholar] [CrossRef]
- Good, A.G.; Muench, D.G. Purification and characterization of an anaerobically induced alanine aminotransferase from barley roots. Plant Physiol. 1992, 99, 1520–1525. [Google Scholar] [CrossRef]
- Muench, D.G.; Good, A.G. Hypoxically inducible barley alanine aminotransferase: cDNA cloning and expression analysis. Plant Mol. Biol. 1994, 24, 417–427. [Google Scholar] [CrossRef]
- Reggiani, R.; Cantu, C.A.; Brambilla, I.; Bertani, A. Accumulation and interconversion of amino acids in rice roots under anoxia. Plant. Cell Physiol. 1988, 29, 981–987. [Google Scholar]
- Appleby, C.A. Leghemoglobin and Rhizobium respiration. Annu Rev. Plant Physiol. 1984, 35, 443–478. [Google Scholar] [CrossRef]
- James, E.K.; Minchin, F.R.; Sprent, J.I. The physiology and nitrogen-fixing capability of aquatically and terrestrially grown Neptunia plena: The importance of nodule oxygen supply. Ann. Bot. 1992, 69, 181–187. [Google Scholar] [CrossRef]
- Schwinghamer, E.; Evans, H.; Dawson, M. Evaluation of effectiveness in mutant strains of Rhizobium by acetylene reduction relative to other criteria of N 2 fixation. Plant Soil 1970, 33, 192–212. [Google Scholar] [CrossRef]
- Krishnan, H.B. NolX of Sinorhizobium fredii USDA257, a type III-secreted protein involved in host range determination, is localized in the infection threads of cowpea (Vigna unguiculata [L.] Walp) and soybean (Glycine max [L.] Merr.) nodules. J. Bacteriol. 2002, 184, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.B.; Song, B.; Oehrle, N.W.; Cameron, J.C.; Jez, J.M. Impact of overexpression of cytosolic isoform of O-acetylserine sulfhydrylase on soybean nodulation and nodule metabolome. Sci. Rep. 2018, 8, 2367. [Google Scholar] [CrossRef]
- Krishnan, H.B.; Alaswad, A.A.; Oehrle, N.W.; Gillman, J.D. Deletion of the SACPD-C locus alters the symbiotic relationship between Bradyrhizobium japonicum USDA110 and soybean, resulting in elicitation of plant defense response and nodulation defects. Mol. Plant-Microbe Interact. 2016, 29, 862–877. [Google Scholar] [CrossRef]
- Krishnan, H.B.; Natarajan, S.S.; Oehrle, N.W.; Garrett, W.M.; Darwish, O. Proteomic analysis of pigeonpea (Cajanus cajan) seeds reveals the accumulation of numerous stress-related proteins. J. Agric. Food Chem. 2017, 65, 4572–4581. [Google Scholar] [CrossRef] [PubMed]
- John, K.M.; Natarajan, S.; Luthria, D.L. Metabolite changes in nine different soybean varieties grown under field and greenhouse conditions. Food Chem. 2016, 211, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Redruello, B.; Ladero, V.; Cuesta, I.; Álvarez-Buylla, J.R.; Martín, M.C.; Fernández, M.; Alvarez, M.A. A fast, reliable, ultra high performance liquid chromatography method for the simultaneous determination of amino acids, biogenic amines and ammonium ions in cheese, using diethyl ethoxymethylenemalonate as a derivatising agent. Food Chem. 2013, 139, 1029–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, K.M.; Luthria, D. Amino acid, organic acid, and sugar profiles of 3 dry bean (Phaseolus vulgaris L.) varieties. J. Food Sci. 2015, 80, C2662–C2669. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Treatment (Location) | |
|---|---|---|
| Non-Flooded (Field) | Flooded (Field) | |
| Biomass (dry weight, kg·ha−1) | 2730 ± 541 | 7021 ± 633 |
| Plant height (cm; n = 24) | 199.39 ± 35.41 | 361.63 ± 29.99 |
| Basal stem diameter (mm; n = 24) | 12.29 ± 2.14 | 18.92 ± 3.49 |
| Pods/plant (total number; n = 24) | 104 ± 43 | 109 ± 30 |
| Seed protein content (%; n = 8) | 35.05 ± 1.17 | 37.94 ± 4.40 |
| Non-Flooded (Greenhouse) | Flooded (Greenhouse) | |
| Number of nodules/plant (n = 15) | 10 ± 3 | 70 ± 19 |
| Plant height (cm; n = 15) | 17.91 ± 2.61 | 27.02 ± 5.53 |
| Nodule fresh weight (grams/nodule; n = 15) | 0.017 ± 0.006 | 0.020 ± 0.007 |
| Nitrogenase activity (C2H4 mmol·h−1·mg−1; acetylene reduction, n = 6) | 5.25 ± 0.45 | 4.90 ± 0.90 |
| # | Protein ID * | MOWSE | Peptides Matched | % C | pI E/T | Mr E/T | MS-Fit NCBIprot: Acc. # |
|---|---|---|---|---|---|---|---|
| 1 | TldD/PmbA family (Neorhizobium huautlense) | 275 | 3 | 8 | 5.00/4.84 | 48,000/47,343 | 1::WP_105374151.1 |
| 2 | No match | ||||||
| 3 | Elongation factor Tu (Neorhizobium huautlense) | 453 | 7 | 17 | 5.60/5.29 | 43,000/42,686 | 1::WP_105374387.1 |
| 4 | Branched chain amino acid ABC transporter (Neorhizobium huautlense) | 781 | 10 | 26 | 5.30/5.34 | 41,000/39,196 | 1::WP_105372985.1 |
| 5 | Phosphonate ABC transporter (Neorhizobium huautlense) | 883 | 8 | 37 | 5.50/5.34 | 36,000/36,238 | 1::WP_105370531.1 |
| 6 | Nitrogenase iron protein (Neorhizobium huautlense) | 938 | 12 | 39 | 5.10/4.83 | 33,000/31,945 | 1::WP_105375136.1 |
| 7 | Sugar ABC transporter (Neorhizobium huautlense) | 509 | 6 | 29 | 5.60/5.59 | 33,000/35,397 | 1::WP_105371152.1 |
| 8 | No match | ||||||
| 9 | No match | ||||||
| 10 | HSP20/alpha crystalline family (Neorhizobium huautlense) | 158 | 2 | 12 | 6.10/5.87 | 28,000/19,836 | 1::WP_105372823.1 |
| 11 | Fructose-6-PO4 aldolase (Mesorhizobium sp.) | 213 | 3 | 15 | 6.15/5.46 | 25,000/23,332 | 1::WP_041004472.1 |
| 12 | Ferritin-like domain-containing (Neorhizobium huautlense) | 346 | 4 | 27 | 5.90/4.92 | 23,000/18,185 | 1::WP_105371465.1 |
| 13 | Peroxiredoxin (Neorhizobium huautlense) | 250 | 3 | 20 | 6.15/5.61 | 23,000/20,344 | 1::WP_105375072.1 |
| 14 | sn-glycerol-3-PO4 ABC trans-porter, UgpB (Neorhizobium huautlense) | 750 | 11 | 28 | 8.00/8.17 | 43,000/48,045 | 1::WP_105370415.1 |
| 15 | sn-glycerol-3-PO4 ABC trans-porter, UgpB (Neorhizobium huautlense) | 646 | 9 | 18 | 8.80/8.17 | 43,000/48,045 | 1::WP_105370415.1 |
| 16 | Phasin (Neorhizobium huautlense) | 390 | 21 | 45 | 7.50/6.75 | 16,000/16,394 | 1::WP_105374889.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krishnan, H.B.; Oehrle, N.W.; Alaswad, A.A.; Stevens, W.; Maria John, K.M.; Luthria, D.L.; Natarajan, S.S. Biochemical and Anatomical Investigation of Sesbania herbacea (Mill.) McVaugh Nodules Grown under Flooded and Non-Flooded Conditions. Int. J. Mol. Sci. 2019, 20, 1824. https://doi.org/10.3390/ijms20081824
Krishnan HB, Oehrle NW, Alaswad AA, Stevens W, Maria John KM, Luthria DL, Natarajan SS. Biochemical and Anatomical Investigation of Sesbania herbacea (Mill.) McVaugh Nodules Grown under Flooded and Non-Flooded Conditions. International Journal of Molecular Sciences. 2019; 20(8):1824. https://doi.org/10.3390/ijms20081824
Chicago/Turabian StyleKrishnan, Hari B., Nathan W. Oehrle, Alaa A. Alaswad, William (Gene) Stevens, K. M. Maria John, Devanand L. Luthria, and Savithiry S. Natarajan. 2019. "Biochemical and Anatomical Investigation of Sesbania herbacea (Mill.) McVaugh Nodules Grown under Flooded and Non-Flooded Conditions" International Journal of Molecular Sciences 20, no. 8: 1824. https://doi.org/10.3390/ijms20081824