Computational Analysis of Dengue Virus Envelope Protein (E) Reveals an Epitope with Flavivirus Immunodiagnostic Potential in Peptide Microarrays

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
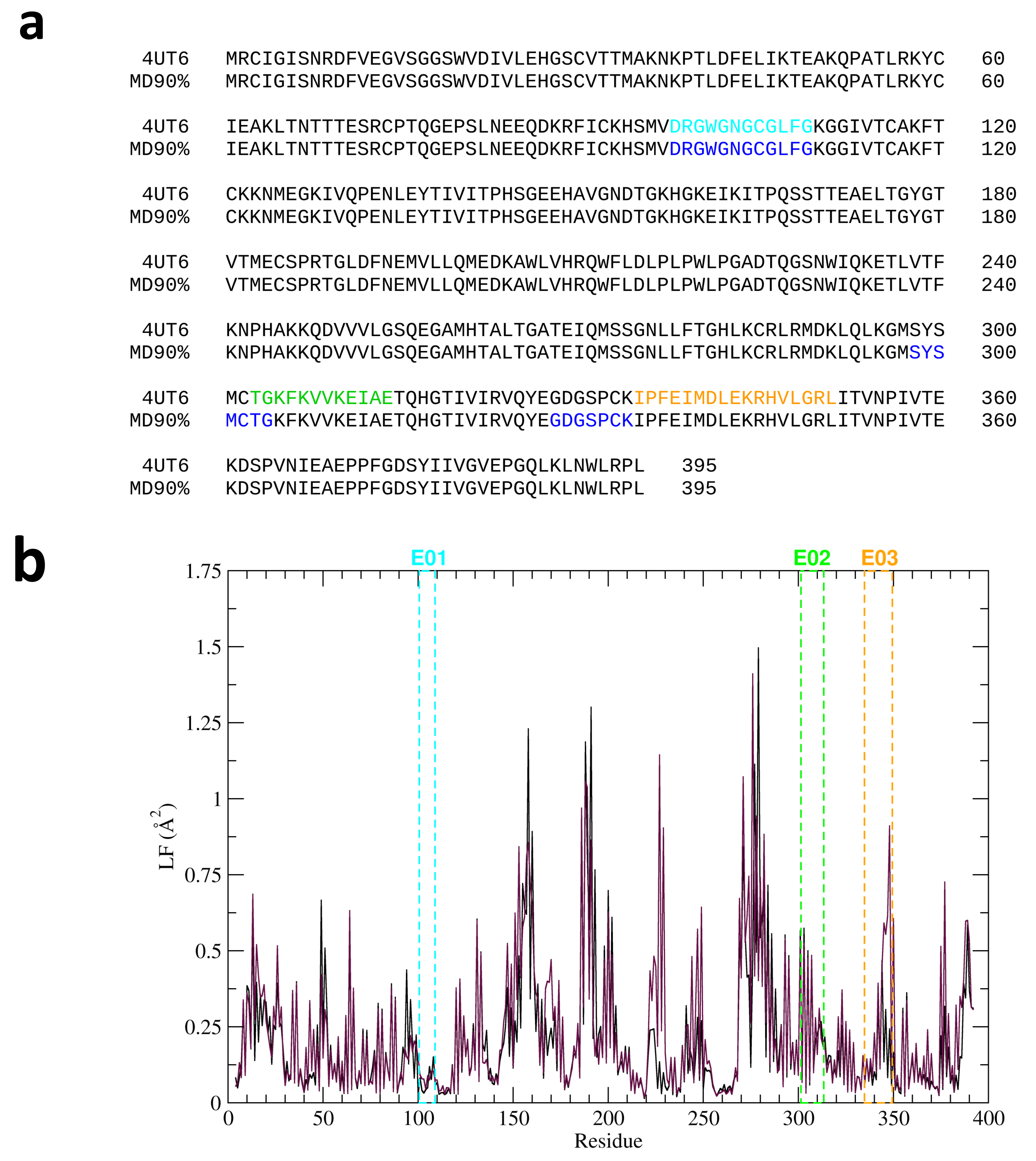
2.1. Computational Analyses
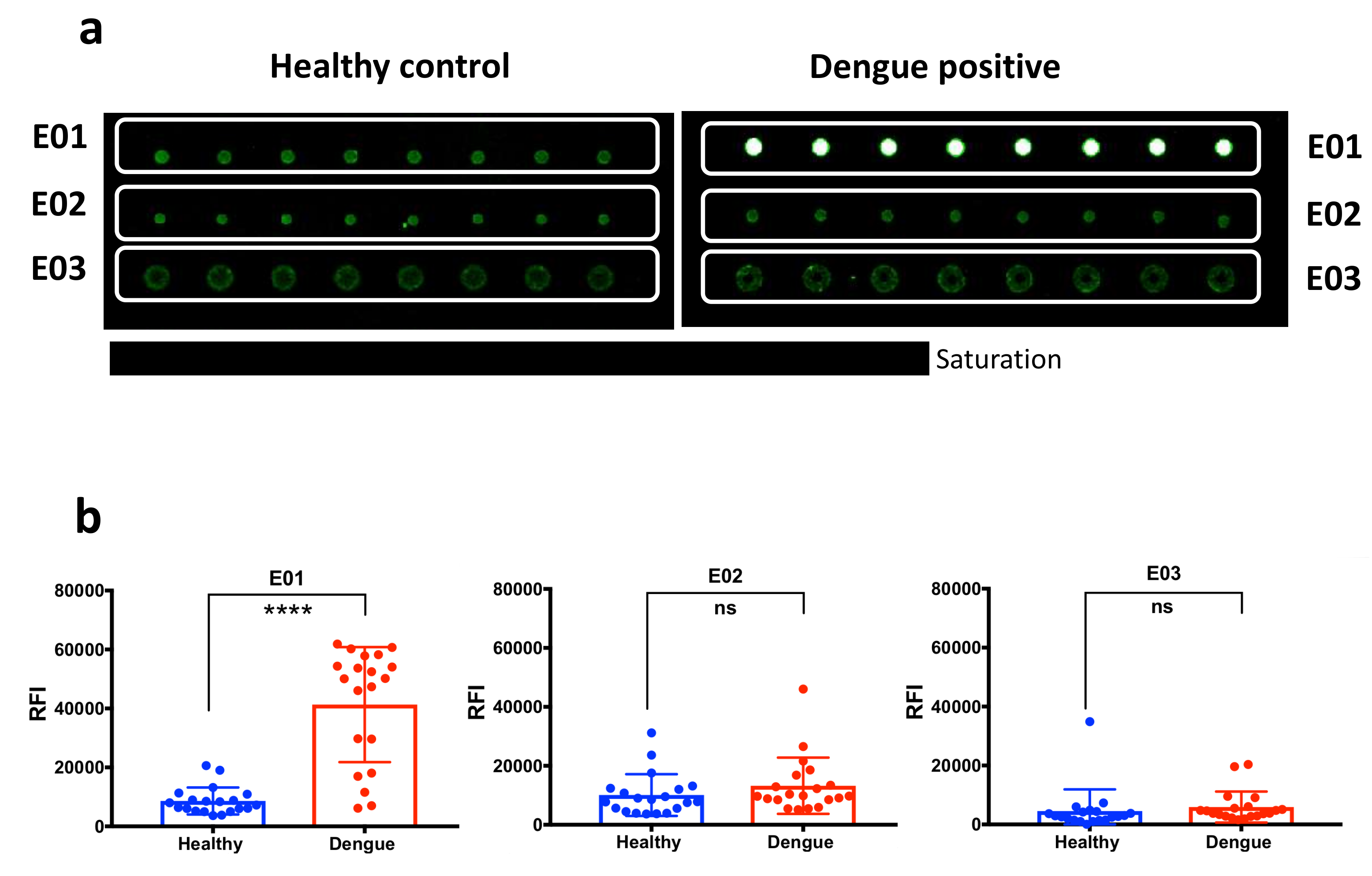
2.2. Immunoreactivity Tests
2.3. Conclusions
3. Materials and Methods
3.1. Samples
3.2. Reagents and Procedures
3.3. Molecular Dynamics Simulations
3.4. Epitope Prediction
3.5. Peptide Synthesis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Microbiol. 2010, 8, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.M. Dengue and dengue hemorrhagic fever. In Handbook Zoonoses, 2nd ed.; CRC Press: New York, NY, USA, 2017; pp. 89–99. [Google Scholar]
- WHO. Dengue and Severe Dengue. WHO Fact Sheet. 2015, pp. 1–4. Available online: www.who.int/mediacentre/factsheets/fs117/en/index.html (accessed on 20 February 2019).
- Slon Campos, J.L.; Mongkolsapaya, J.; Screaton, G.R. The immune response against flaviviruses. Nat. Immunol. 2018, 19, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Mizumoto, K.; Ejima, K.; Yamamoto, T.; Nishiura, H. On the risk of severe dengue during secondary infection: A systematic review coupled with mathematical modeling. J. Vect. Borne Dis. 2014, 51, 153–164. [Google Scholar]
- Dussart, P.; Labeau, B.; Lagathu, G.; Louis, P.; Nunes, M.R.T.; Rodrigues, S.G.; Storck-Herrmann, C.; Cesaire, R.; Morvan, J.; Flamand, M.; et al. Evaluation of an Enzyme Immunoassay for Detection of Dengue Virus NS1 Antigen in Human Serum. Clin. Vacc. Immunol. 2006, 13, 1185–1189. [Google Scholar] [CrossRef]
- Hunsperger, E.A.; Yoksan, S.; Buchy, P.; Nguyen, V.C.; Sekaran, S.D.; Enria, D.A.; Pelegrino, J.L.; Vázquez, S.; Artsob, H.; Drebot, M.; et al. Evaluation of commercially available anti-dengue virus immunoglobulin M tests. Emerg. Infect. Dis. 2009, 15, 436–440. [Google Scholar] [CrossRef]
- Hunsperger, E.A.; Sharp, T.M.; Lalita, P.; Tikomaidraubuta, K.; Cardoso, Y.R.; Naivalu, T.; Khan, A.S.; Marfel, M.; Hancock, W.T.; Tomashek, K.M.; et al. Use of a rapid test for diagnosis of dengue during suspected dengue outbreaks in resource-limited regions. J. Clin. Microbiol. 2016, 54, 2090–2095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, S.; Pereira Carvalho, W.S.; Jiang, Y.; Serpe, M.J. Portable point-of-care diagnostic devices. Anal. Methods 2016, 8, 7847–7867. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Zapatero-Rodríguez, J.; Estrela, P.; O’Kennedy, R. Point-of-Care Diagnostics in Low Resource Settings: Present Status and Future Role of Microfluidics. Biosensors 2015, 5, 577–601. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Wei, J.; Wang, Y.; Zhang, H.; Wang, Z. The role of peptide microarrays in biomedical research. Anal. Methods 2018, 10, 4614–4624. [Google Scholar] [CrossRef]
- Carmona, SJ.; Nielsen, M.; Schafer-Nielsen, C.; Mucci, J.; Altcheh, J.; Balouz, V.; Tekiel, V.; Frasch, A.C.; Campetella, O.; Buscaglia, C.A.; et al. Towards High-throughput Immunomics for Infectious Diseases: Use of Next-generation Peptide Microarrays for Rapid Discovery and Mapping of Antigenic Determinants. Mol. Cell Proteom. 2015, 14, 1871–1884. [Google Scholar] [CrossRef]
- Shen, G.; Behera, D.; Bhalla, M.; Nadas, A.; Laal, S. Peptide-based antibody detection for tuberculosis diagnosis. Clin. Vacc. Immunol. 2009, 16, 49–54. [Google Scholar] [CrossRef]
- Winkler, G.; Heinz, FX.; Kunz, C. Studies on the glycosylation of flavivirus E proteins and the role of carbohydrate in antigenic structure. Virology 1987, 159, 237–243. [Google Scholar] [CrossRef]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. A ligand-binding pocket in the dengue virus envelope glycoprotein. Proc. Natl. Acad. Sci. USA 2003, 100, 6986–6991. [Google Scholar] [CrossRef] [Green Version]
- Fritz, R.; Blazevic, J.; Taucher, C.; Pangerl, K.; Heinz, F.X.; Stiasny, K. The unique transmembrane hairpin of flavivirus fusion protein E is essential for membrane fusion. J. Virol. 2011, 85, 4377–4385. [Google Scholar] [CrossRef] [PubMed]
- Sola, L.; Damin, F.; Gagni, P.; Consonni, R.; Chiari, M. Synthesis of Clickable Polymers by Postpolymerization Modification: Application in Microarray Technology. Langmuir 2016, 32, 10284–10295. [Google Scholar] [CrossRef]
- Gori, A.; Sola, L.; Gagni, P.; Bruni, G.; Liprino, M.; Peri, C.; Colombo, G.; Cretich, M.; Chiari, M. Screening Complex Biological Samples with Peptide Microarrays: The Favorable Impact of Probe Orientation via Chemoselective Immobilization Strategies on Clickable Polymeric Coatings. Bioconjug. Chem. 2016, 27, 2669–2677. [Google Scholar] [CrossRef]
- Sola, L.; Gagni, P.; D’Annessa, I.; Capelli, R.; Bertino, C.; Romanato, A.; Damin, F.; Bergamaschi, G.; Marchisio, E.; Cuzzocrea, A.; et al. Enhancing Antibody Serodiagnosis Using a Controlled Peptide Coimmobilization Strategy. ACS Infect. Dis. 2018, 4, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, P.; Puijk, W.C.; Boshuizen, R.S.; van Dijken, P.; Slootstra, J.W.; Beurskens, F.J.; Parren, P.W.H.I.; Huber, A.; Bachmann, M.F.; Meloen, R.H. Functional Reconstruction of Structurally Complex Epitopes using CLIPSTM Technology. Open Vacc. J. 2009, 2, 56–67. [Google Scholar] [CrossRef]
- Smeenk, L.E.J.; Dailly, N.; Hiemstra, H.; Van Maarseveen, J.H.; Timmerman, P. Synthesis of water-soluble scaffolds for peptide cyclization, labeling, and ligation. Org. Lett. 2012, 14, 1194–1197. [Google Scholar] [CrossRef] [PubMed]
- Van de Langemheen, H.; van Hoeke, M.; Quarles van Ufford, H.C.; Kruijtzer, J.A.W.; Liskamp, R.M.J. Scaffolded multiple cyclic peptide libraries for protein mimics by native chemical ligation. Org. Biomol. Chem. 2014, 12, 4471–4478. [Google Scholar] [CrossRef]
- Meuleman, T.J.; Dunlop, J.I.; Owsianka, A.M.; van de Langemheen, H.; Patel, A.H.; Liskamp, R.M.J. Immobilization by Surface Conjugation of Cyclic Peptides for Effective Mimicry of the HCV-Envelope E2 Protein as a Strategy toward Synthetic Vaccines. Bioconjug. Chem. 2018, 29, 1091–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdine, G.L.; Hilinski, G.J. Stapled peptides for intracellular drug targets. Methods Enzymol. 2012, 503, 3–33. [Google Scholar]
- Bird, G.H.; Irimia, A.; Ofek, G.; Kwong, P.D.; Wilson, I.A.; Walensky, L.D. Stapled HIV-1 peptides recapitulate antigenic structures and engage broadly neutralizing antibodies. Nat. Struct. Mol. Biol. 2014, 21, 1058–1067. [Google Scholar] [CrossRef] [Green Version]
- Morelli, L.; Fallarini, S.; Lombardi, G.; Colombo, C.; Lay, L.; Compostella, F. Synthesis and biological evaluation of a trisaccharide repeating unit derivative of Streptococcus pneumoniae 19A capsular polysaccharide. Bioorg. Med. Chem. 2018, 26, 5682–5690. [Google Scholar] [CrossRef] [PubMed]
- Legnani, L.; Compostella, F.; Sansone, F.; Toma, L. Cone Calix[4]arenes with Orientable Glycosylthioureido Groups at the Upper Rim: An In-Depth Analysis of Their Symmetry Properties. J. Org. Chem. 2015, 80, 7412–7418. [Google Scholar] [CrossRef]
- Sharma, K.K.; Lim, X.X.; Tantirimudalige, S.N.; Gupta, A.; Marzinek, J.K.; Holdbrook, D.; Lim, X.Y.E.; Bond, P.J.; Anand, G.S.; Wohland, T. Infectivity of Dengue Virus Serotypes 1 and 2 is Correlated to E Protein Intrinsic Dynamics But Not to Envelope Conformations. Structure 2019, 4, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Guillot, B.; Ruiz-Lopez, M.F.; Genoni, A. Libraries of Extremely Localized Molecular Orbitals. 1. Model Molecules Approximation and Molecular Orbitals Transferability. J. Chem. Theory Comput. 2016, 12, 1052–1067. [Google Scholar] [CrossRef]
- Meyer, B.; Guillot, B.; Ruiz-Lopez, M.F.; Jelsch, C.; Genoni, A. Libraries of Extremely Localized Molecular Orbitals. 2. Comparison with the Pseudoatoms Transferability. J. Chem. Theory Comput. 2016, 12, 1068–1081. [Google Scholar] [CrossRef]
- Pirri, G.; Damin, F.; Chiari, M.; Bontempi, E.; Depero, L.E. Characterization of A Polymeric Adsorbed Coating for DNA Microarray Glass Slides. Anal. Chem. 2004, 76, 1352–1358. [Google Scholar] [CrossRef]
- Cretich, M.; Pirri, G.; Damin, F.; Solinas, I.; Chiari, M. A new polymeric coating for protein microarrays. Anal. Biochem. 2004, 332, 67–74. [Google Scholar] [CrossRef]
- Rouvinski, A.; Guardado-Calvo, P.; Barba-Spaeth, G.; Duquerroy, S.; Vaney, M.-C.; Kikuti, C.M.; Sanchez, M.E.N.; Dejnirattisai, W.; Wongwiwat, W.; Haouz, A.; et al. Recognition determinants of broadly neutralizing human antibodies against dengue viruses. Nature 2015, 520, 109–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Joung, I.S.; Cheatham, T.E. Determination of Alkali and Halide Monovalent Ion Parameters for Use in Explicitly Solvated Biomolecular Simulations. J. Phys. Chem. B. 2008, 112, 9020–9041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loncharich, R.J.; Brooks, B.R.; Pastor, R.W. Langevin dynamics of peptides: The frictional dependence of isomerization rates of N-acetylalanyl-N′-methylamide. Biopolymers 1992, 32, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Åqvist, J.; Wennerström, P.; Nervall, M.; Bjelic, S.; Brandsdal, B.O. Molecular dynamics simulations of water and biomolecules with a Monte Carlo constant pressure algorithm. Chem. Phys. Lett. 2004, 384, 288–294. [Google Scholar] [CrossRef]
- Ryckaert, J.-P.; Ciccotti, G.; Berendsen, H.J. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N log( N ) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Le Grand, S.; Götz, A.W.; Walker, R.C. SPFP: Speed without compromise—A mixed precision model for GPU accelerated molecular dynamics simulations. Comput. Phys. Commun. 2013, 184, 374–380. [Google Scholar] [CrossRef]
- Daura, X.; Gademann, K.; Jaun, B.; Seebach, D.; van Gunsteren, W.F.; Mark, A.E. Peptide Folding: When Simulation Meets Experiment. Angew. Chem. Int. Ed. 1999, 38, 236–240. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Scarabelli, G.; Morra, G.; Colombo, G. Predicting interaction sites from the energetics of isolated proteins: A new approach to epitope mapping. Biophys. J. 2010, 98, 1966–1975. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, M.; D’Annessa, I.; Moroni, E.; Morra, G.; Paladino, A.; Rinaldi, S.; Compostella, F.; Colombo, G. Allosteric Modulators of HSP90 and HSP70: Dynamics Meets Function through Structure-Based Drug Design. J. Med. Chem. 2019, 62, 60–87. [Google Scholar] [CrossRef]
- Peri, C.; Gagni, P.; Combi, F.; Gori, A.; Chiari, M.; Longhi, R.; Cretich, M.; Colombo, G. 2 Rational epitope design for protein targeting. ACS Chem. Biol. 2013, 8, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Genoni, A.; Morra, G.; Colombo, G. Identification of domains in protein structures from the analysis of intramolecular interactions. J. Phys. Chem. B 2012, 116, 3331–3343. [Google Scholar] [CrossRef] [PubMed]
- Peri, C.; Gori, A.; Gagni, P.; Sola, L.; Girelli, D.; Sottotetti, S.; Cariani, L.; Chiari, M.; Cretich, M.; Colombo, G. Evolving serodiagnostics by rationally designed peptide arrays: The Burkholderia paradigm in Cystic Fibrosis. Sci. Rep. 2016, 6, 32873. [Google Scholar] [CrossRef] [PubMed]
- Capelli, R.; Matterazzo, E.; Amabili, M.; Peri, C.; Gori, A.; Gagni, P.; Chiari, M.; Lertmemongkolchai, G.; Cretich, M.; Bolognesi, M.; et al. Designing Probes for Immunodiagnostics: Structural Insights into an Epitope Targeting Burkholderia Infections. ACS Infect. Dis. 2017, 3, 736–743. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Code | Peptide Sequence |
|---|---|
| E01 | Prg-(O2Oc)2-DRGWGNGCGLFG |
| E02 | Prg-(O2Oc)2-TGKFKVVKEIAE |
| E03 | Prg-(O2Oc)2-IPFEIMDLEKRHVLGRL |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergamaschi, G.; Fassi, E.M.A.; Romanato, A.; D'Annessa, I.; Odinolfi, M.T.; Brambilla, D.; Damin, F.; Chiari, M.; Gori, A.; Colombo, G.; et al. Computational Analysis of Dengue Virus Envelope Protein (E) Reveals an Epitope with Flavivirus Immunodiagnostic Potential in Peptide Microarrays. Int. J. Mol. Sci. 2019, 20, 1921. https://doi.org/10.3390/ijms20081921
Bergamaschi G, Fassi EMA, Romanato A, D'Annessa I, Odinolfi MT, Brambilla D, Damin F, Chiari M, Gori A, Colombo G, et al. Computational Analysis of Dengue Virus Envelope Protein (E) Reveals an Epitope with Flavivirus Immunodiagnostic Potential in Peptide Microarrays. International Journal of Molecular Sciences. 2019; 20(8):1921. https://doi.org/10.3390/ijms20081921
Chicago/Turabian StyleBergamaschi, Greta, Enrico M. A. Fassi, Alessandro Romanato, Ilda D'Annessa, Maria Teresa Odinolfi, Dario Brambilla, Francesco Damin, Marcella Chiari, Alessandro Gori, Giorgio Colombo, and et al. 2019. "Computational Analysis of Dengue Virus Envelope Protein (E) Reveals an Epitope with Flavivirus Immunodiagnostic Potential in Peptide Microarrays" International Journal of Molecular Sciences 20, no. 8: 1921. https://doi.org/10.3390/ijms20081921