The Application of Selenium and Copper Nanoparticles Modifies the Biochemical Responses of Tomato Plants under Stress by Alternaria solani

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
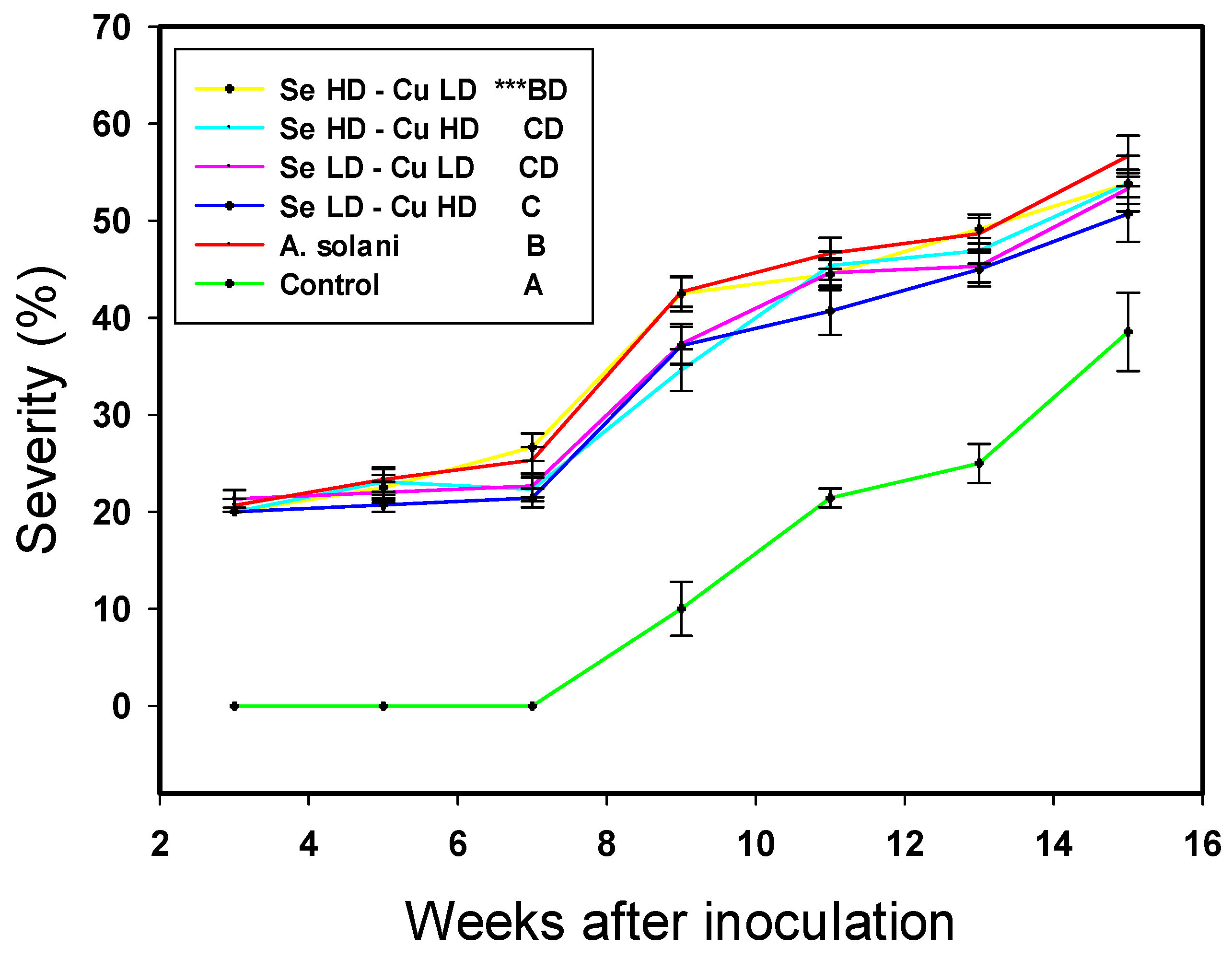
2.1. Crop Growth and Severity of Alternaria solani
2.2. Photosynthetic Pigments
2.3. Non-Enzymatic Compounds
2.4. Enzymatic Compounds
2.5. Total Antioxidant Capacity
3. Discussion
3.1. Crop Growth and Severity of Alternaria solani
3.2. Photosynthetic Pigments
3.3. Non-Enzymatic Compounds
3.4. Enzymatic Compounds
3.5. Total Antioxidant Capacity
4. Materials and Methods
4.1. Crop Development
4.2. Application of Treatments
4.3. Inoculation Preparation and Evaluation of the Severity of Alternaria solani
4.4. Sample Processing
4.5. Photosynthetic Pigments
4.6. Non-Enzymatic Antioxidant Compounds
4.7. Enzymatic Antioxidant Compounds and Phenylalanine Ammonia Lyase
4.8. Antioxidant Capacity
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Akhtar, K.P.; Saleem, M.Y.; Iqbal, Q.; Asghar, M.; Hameed, A.; Sarwar, N. Evaluation of tomato genotypes for late blight resistance using low tunnel assay. J. Plant Pathol. 2016, 98, 421–428. [Google Scholar] [CrossRef]
- Castagna, A.; Chiavaro, E.; Dall’Asta, C.; Rinaldi, M.; Galaverna, G.; Ranieri, A. Effect of postharvest UV-B irradiation on nutraceutical quality and physical properties of tomato fruits. Food Chem. 2013, 137, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, B.S. NSA-39406-nanotechnology-in-agri-food-production—An-overview. Nanotechnol. Sci. Appl. 2014, 31–53. [Google Scholar] [CrossRef] [PubMed]
- Bebber, D.P.; Gurr, S.J. Crop-destroying fungal and oomycete pathogens challenge food security. Fungal. Genet. Biol. 2015, 74, 62–64. [Google Scholar] [CrossRef]
- Bramhanwade, K.; Shende, S.; Bonde, S.; Gade, A.; Rai, M. Fungicidal activity of Cu nanoparticles against Fusarium causing crop diseases. Environ. Chem. Lett. 2016, 14, 229–235. [Google Scholar] [CrossRef]
- Monteiro, S.; Pizza, T.; Carneeiro, G.; Davidson, E.; Pignoni, E.; Teixeira, M.Z.; Da, E.M.; Romano, B.; Vasconcelos, C.; Gomes, C.J. Effect of biotherapic of Alternaria solani on the early blight of tomato-plant and the in vitro development of the fungus. Int. J. High Dilution Res. 2010, 9, 147–155. [Google Scholar]
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Simon-Delso, N.; San Martin, G.; Bruneau, E.; Hautier, L. Time-to-death approach to reveal chronic and cumulative toxicity of a fungicide for honeybees not revealed with the standard ten-day test. Sci. Rep. 2018, 8, 7241. [Google Scholar] [CrossRef] [PubMed]
- Thiruvengadam, M.; Rajakumar, G.; Chung, I.-M. Nanotechnology: Current uses and future applications in the food industry. 3 Biotech 2018, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Ingle, A. Role of nanotechnology in agriculture with special reference to management of insect pests. Appl. Microbiol. Biotechnol. 2012, 94, 287–293. [Google Scholar] [CrossRef]
- Kanhed, P.; Birla, S.; Gaikwad, S.; Gade, A.; Seabra, A.B.; Rubilar, O.; Duran, N.; Rai, M. In vitro antifungal efficacy of copper nanoparticles against selected crop pathogenic fungi. Mater. Lett. 2014, 115, 13–17. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalaby, T.A.; Bayoumi, Y.; Abdalla, N.; Taha, H. Nanoparticles, soils, plants and sustainable agriculture. In Nanoscience in Food and Agriculture 1; Ranjan, S., Dasgupta, N., Lichtfouse, E., Eds.; Springer International Publishing: Basel, Switzerland, 2016; Volume 20, pp. 283–312. ISBN 978-3-319-39302-5. [Google Scholar]
- Raliya, R.; Saharan, V.; Dimkpa, C.; Biswas, P. Nanofertilizer for precision and sustainable agriculture: Current state and future perspectives. J. Agric. Food Chem. 2018, 66, 6487–6503. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Maldonado, A.; Ortega-Ortiz, H.; González-Morales, S.; Morelos-Moreno, Á.; Cabrera-de la Fuente, M.; Sandoval-Rangel, A.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Nanoparticles and nanomaterials as plant biostimulants. Int. J. Mol. Sci. 2019, 20, 162. [Google Scholar] [CrossRef]
- Feregrino-perez, A.A.; Magaña-lópez, E.; Guzmán, C.; Esquivel, K. A general overview of the bene fi ts and possible negative e ff ects of the nanotechnology in horticulture. Sci. Hortic. 2018, 238, 126–137. [Google Scholar] [CrossRef]
- Lewis, R.W.; Bertsch, P.M.; Mcnear, D.H. Nanotoxicity of engineered nanomaterials (ENMs) to environmentally relevant beneficial soil bacteria—A critical review. Nanotoxicology 2019, 14, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, H.; Al-Dhabi, A.; Karuppiah, P.; Kumar Rajaram, S. Microbial synthesis of selenium nanocomposite using Saccharomyces cerevisiae and its antimicrobial activity againts pathogens causing nosocomial infection. Chalcogenide Lett. 2012, 9, 509–515. [Google Scholar]
- Abdel-Kareem, M.M.; Ahmed-Zohri, A.-N. Inhibition of three toxigenic fungal strains and their toxins production using selenium nanoparticles. Czech Mycol. 2017, 69, 193–204. [Google Scholar] [CrossRef]
- Shirsat, S.; Kadam, A.; Naushad, M.; Mane, R.S. Selenium nanostructures: Microbial synthesis and applications. RSC Adv. 2015, 5, 92799–92811. [Google Scholar] [CrossRef]
- Kasana, R.C.; Panwar, N.R.; Kaul, R.K.; Kumar, P. Biosynthesis and effects of copper nanoparticles on plants. Environ. Chem. Lett. 2017, 15, 233–240. [Google Scholar] [CrossRef]
- Adhikari, T.; Kundu, S.; Biswas, A.K.; Kundu, S.; Tarafdar, J.C.; Rao, A.S. Effect of copper oxide nano particle on seed germination of selected crops. J. Agric. Sci. Technol. 2012, 2, 815–823. [Google Scholar]
- Ponmurugan, P.; Manjukarunambika, K.; Elango, V.; Gnanamangai, B.M. Antifungal activity of biosynthesised copper nanoparticles evaluated against red root-rot disease in tea plants. J. Exp. Nanosci. 2016, 11, 1019–1031. [Google Scholar] [CrossRef]
- Ouda, S.M. Antifungal activity of silver and copper nanoparticles pathogens, Alternaria alternata and Botrytis cinerea. Res. J. Microbiol. 2014, 9, 34–42. [Google Scholar] [CrossRef]
- Hernández-Hernández, H.; González-Morales, S.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Juárez-Maldonado, A. Effects of chitosan–PVA and Cu nanoparticles on the growth and antioxidant capacity of tomato under saline stress. Molecules 2018, 23, 178. [Google Scholar] [CrossRef]
- Nandini, B.; Hariprasad, P.; Prakash, H.S.; Shetty, H.S.; Geetha, N. Trichogenic-selenium nanoparticles enhance disease suppressive ability of Trichoderma against downy mildew disease caused by Sclerospora graminicola in pearl millet. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sathiyabama, M.; Manikandan, A. Application of copper-chitosan nanoparticles stimulate growth and induce resistance in finger millet (Eleusine coracana Gaertn.) plants against blast disease. J. Agric. Food Chem. 2018, 66, 1784–1790. [Google Scholar] [CrossRef]
- Latha, P.; Anand, T.; Ragupathi, N.; Prakasam, V.; Samiyappan, R. Antimicrobial activity of plant extracts and induction of systemic resistance in tomato plants by mixtures of PGPR strains and zimmu leaf extract against Alternaria solani. Biol. Control 2009, 50, 85–93. [Google Scholar] [CrossRef]
- Swarupa, V.; Ravishankar, K.V.; Rekha, A. Characterization of tolerance to Fusarium oxysporum f.sp., cubense infection in banana using suppression subtractive hybridization and gene expression analysis. Physiol. Mol. Plant Pathol. 2013, 83, 1–7. [Google Scholar] [CrossRef]
- Cumplido-nájera, C.F.; González-morales, S.; Ortega-ortíz, H. Scientia Horticulturae The application of copper nanoparticles and potassium silicate stimulate the tolerance to Clavibacter michiganensis in tomato plants. Sci. Hortic. 2019, 245, 82–89. [Google Scholar] [CrossRef]
- Saharan, V.; Sharma, G.; Yadav, M.; Kumari, M.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P. Synthesis and in vitro antifungal efficacy of Cu—Chitosan nanoparticles against pathogenic fungi of tomato. Int. J. Biol. Macromol. 2015, 75, 346–353. [Google Scholar] [CrossRef]
- Khiralla, G.M.; El-Deeb, B.A. Antimicrobial and antibiofilm effects of selenium nanoparticles on some foodborne pathogens. LWT Food Sci. Technol. 2015, 63, 1001–1007. [Google Scholar] [CrossRef]
- Ismail, A.-W.A.; Sidkey, N.M.; Arafa, R.A.; Fathy, R.M.; El-Batal, A.I.; Yean Yean, C. Evaluation of in vitro antifungal activity of silver and selenium nanoparticles against Alternaria solani caused early blight disease on potato. Br. Biotechnol. J. 2016, 12, 1–11. [Google Scholar] [CrossRef]
- Dehgahi, R.; Subramaniam, S.; Zakaria, L.; Joniyas, A.; Firouzjahi, F.B.; Haghnama, K.; Razinataj, M. Review of research on fungal pathogen attack and plant defense mechanism against pathogen. Int. J. Sci. Res. Agric. Sci. 2016, 2, 197–208. [Google Scholar] [CrossRef]
- Rajiv, P.; Vanathi, P.; Thangamani, A. An investigation of phytotoxicity using Eichhornia mediated zinc oxide nanoparticles on Helianthus annuus. Biocatal. Agric. Biotechnol. 2018, 16, 419–424. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Priyanka, N.; Manikandan, K.; Ganeshbabu, I.; Indiraarulselvi, P.; Geetha, N.; Muralikrishna, K.; Bhattacharya, R.C.; Tiwari, M.; Sharma, N.; et al. Enhanced plant growth promoting role of phycomolecules coated zinc oxide nanoparticles with P supplementation in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 110, 118–127. [Google Scholar] [CrossRef]
- Chandra, S.; Chakraborty, N.; Dasgupta, A.; Sarkar, J.; Panda, K.; Acharya, K. Chitosan nanoparticles: A positive modulator of innate immune responses in plants. Sci. Rep. 2015, 5, 15195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marslin, G.; Sheeba, C.J.; Franklin, G. Nanoparticles alter secondary metabolism in plants via ROS burst. Front. Plant Sci. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil-Maurizi, C.; Poinssot, B. Role of glutathione in plant signaling under biotic stress. Plant Signal. Behav. 2012, 7, 210–212. [Google Scholar] [CrossRef] [Green Version]
- Raigond, P.; Raigond, B.; Kaundal, B.; Singh, B.; Joshi, A.; Dutt, S. Effect of zinc nanoparticles on antioxidative system of potato plants. J. Environ. Biol. 2017, 38, 435–439. [Google Scholar] [CrossRef]
- Lopez-Vargas, E.R.; Ortega-ortiz, H.; Cadenas-pliego, G.; De-Alba-Romenus, K.; Cabrera-De-La-Fuente, M.; Benavides-Mendoza, A.; Juarez-Maldonado, A. Foliar application of copper nanoparticles increases the fruit quality and the content of bioactive compounds in tomatoes. Appl. Sci. 2018, 8, 1020. [Google Scholar] [CrossRef]
- Juarez-Maldonado, A.; Ortega-Ortíz, H.; Pérez-Labrada, F.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Cu Nanoparticles absorbed on chitosan hydrogels positively alter morphological, production, and quality characteristics of tomato. J. Appl. Bot. Food Qual. 2016, 89, 183–189. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Belliraj, N.; Bossmann, S.H.; Prasad, P.V.V. High-temperature stress alleviation by selenium nanoparticle treatment in grain sorghum. ACS Omega 2018, 3, 2479–2491. [Google Scholar] [CrossRef]
- Gowayed, S.M.; Kadasa, N.M. Effect of zinc oxide nanoparticles on antioxidative system of faba bean (Vicia faba L) seedling expose to cadmium. Life Sci. J. 2016, 13, 18–27. [Google Scholar] [CrossRef]
- Astaneh, R.K.; Bolandnazar, S.; Nahandi, F.Z.; Oustan, S. Effect of selenium application on phenylalanine ammonia-lyase (PAL) activity, phenol leakage and total phenolic content in garlic (Allium sativum L.) under NaCl stress. Inf. Process. Agric. 2018, 5, 339–344. [Google Scholar] [CrossRef]
- Heldt, H.-W.; Piechulla, B. Phenylpropanoids comprise a multitude of plant secondary metabolites and cell wall components. Plant. Biochem. 2011, 431–449. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant. Soil 1961, 15, 134–154. [Google Scholar] [CrossRef] [Green Version]
- Safari, M.; Oraghi Ardebili, Z.; Iranbakhsh, A. Selenium nano-particle induced alterations in expression patterns of heat shock factor A4A (HSFA4A), and high molecular weight glutenin subunit 1Bx (Glu-1Bx) and enhanced nitrate reductase activity in wheat (Triticum aestivum L.). Acta Physiol. Plant. 2018, 40. [Google Scholar] [CrossRef]
- Kong, H.; Yang, J.; Zhang, Y.; Fang, Y.; Nishinari, K.; Phillips, G.O. Synthesis and antioxidant properties of gum arabic-stabilized selenium nanoparticles. Int. J. Biol. Macromol. 2014, 65, 155–162. [Google Scholar] [CrossRef]
- Cadenas-Pliego, G.; Pérez-Alvarez, M.; Ávila, O.; Sierra-Ávila, R.; Ortega-Ortiz, H.; Betancourt-Galindo, R.; Jiménez-Regalado, E.; Barriga-Castro, E.; Palacios-Mireles, I.M. Proceso De Síntesis De Nanopartículas Metálicas Mediante el Uso De Moléculas Bi-Funcionales. Expediente: MX/a/2013/015221, Folio MX/E/2013/09476. 2013. Available online: https://siga.impi.gob.mx/newSIGA/content/common/principal.jsf (accessed on 19 April 2019).
- Benhamou, N.; Bélanger, R.R. Benzothiadiazole-mediated induced resistance to fusarium oxysporum f. sp. radicis-lycopersici in tomato. Plant. Physiol. 1998, 118, 1203–1212. [Google Scholar] [CrossRef]
- Smith, J.A.; Hammerschmidt, R.; Fulbright, D.W. Rapid induction of systemic resistance in cucumber by Pseudomonas syringae pv. syringae. Physiol. Mol. Plant. Pathol. 1991, 38, 223–235. [Google Scholar] [CrossRef]
- Diener, A.C.; Ausubel, F.M. Resistance to Fusarium oxysporum 1, a dominant Arabidopsis disease-resistance gene, is not race specific. Genetics 2005, 171, 305–321. [Google Scholar] [CrossRef]
- United States Department of Agriculture. United States Standards for Grades of Fresh Tomatoes; United States Department of Agriculture: Washington, DC, USA, 2016.
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. J. Jpn. Soc. Food Sci. Technol. Shokuhin Kagaku Kogaku Kaishi 1992, 39, 925–928. [Google Scholar] [CrossRef]
- Arvouet-Grand, A.; Vennat, B.; Pourrat, A.; Legret, P. Standardization of a propolis extract and identification of the main constituents. J. Pharm. Belg. 1994, 49, 462–468. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Xue, T.; Hartikainen, H.; Piironen, V. Antioxidative and growth-promoting effect of selenium on senescing lettuce. Plant. Soil 2001, 237, 55–61. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Purification of ascorbate peroxidase in spinach chloroplasts; its inactivation in ascorbate-depleted medium and reactivation by monodehydroascorbate radical. Plant. Cell Physiol. 1987, 28, 131–140. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–120. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T. Leaf senescence: Sorrelated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Sykłowska-Baranek, K.; Pietrosiuk, A.; Naliwajski, M.R.; Kawiak, A.; Jeziorek, M.; Wyderska, S.; Łojkowska, E.; Chinou, I. Effect of l-phenylalanine on PAL activity and production of naphthoquinone pigments in suspension cultures of Arnebia euchroma (Royle) Johnst. In Vitro Cell. Dev. Biol.Plant. 2012, 48, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Treatment | SD (mm) | PH (cm) | NL | NC | NF | FW (kg) | AFW (g) | FAB (g) | DAB (g) |
|---|---|---|---|---|---|---|---|---|---|
| Control | 13.01 a | 2.30 a | 29.21 a | 6.86 c | 63.21 ab | 6.80 ab | 107.92 ab | 1659.1 a | 233.17 a |
| A. solani | 12.58 a | 2.11 b | 27.53 b | 7.27 ab | 67.00 a | 7.04 a | 105.71 ab | 1608.2 ab | 224.15 ab |
| Se LD-Cu LD | 13.06 a | 2.10 b | 28.07 ab | 7.47 a | 68.73 a | 7.05 a | 102.74 ab | 1656.0 a | 232.41 a |
| Se LD-Cu HD | 12.43 a | 2.09 b | 28.07 ab | 7.21 abc | 59.86 b | 6.52 ab | 109.10 a | 1565.2 ab | 207.44 ab |
| Se HD-Cu LD | 12.42 a | 2.12 b | 27.08 b | 7.00 bc | 61.50 ab | 6.07 b | 99.39 b | 1406.9 b | 194.13 b |
| Se HD-Cu HD | 13.18 a | 2.14 b | 29.23 a | 7.31 ab | 66.15 ab | 6.75 ab | 102.87 ab | 1560.9 ab | 217.79 ab |
| CV | 11.33 | 8.2 | 6.09 | 7.35 | 14.65 | 16.17 | 11.5 | 19.19 | 21.56 |
| Treatment | Chlorophyll a (mg 100 g−1 FW) | Chlorophyll b (mg 100 g−1 FW) | Total Chlorophyll (mg 100 g−1 FW) |
|---|---|---|---|
| Control | 13.01 a | 2.30 a | 29.21 a |
| A. solani | 12.58 a | 2.11 b | 27.53 b |
| Se LD-Cu LD | 13.06 a | 2.10 b | 28.07 ab |
| Se LD-Cu HD | 12.43 a | 2.09 b | 28.07 ab |
| Se HD-Cu LD | 12.42 a | 2.12 b | 27.08 b |
| Se HD-Cu HD | 13.18 a | 2.14 b | 29.23 a |
| CV | 11.33 | 8.2 | 6.09 |
| Organ | Treatment | Vitamin C (mg 100 g−1 FW) | Glutathione (mmol 100 g−1 DW) | Flavonoids (mg 100 g−1 DW) | Phenols (mg g−1 DW) | Lycopene (mg 100 g−1 DW) |
|---|---|---|---|---|---|---|
| Leaves | Control | 167.2 a | 1.47 b | 62.4 a | 6.65 a | Nd |
| A. solani | 124.7 bc | 2.31 ab | 38.3 b | 7.09 a | Nd | |
| Se LD-Cu LD | 140.8 ab | 1.89 ab | 42.7 b | 7.91 a | Nd | |
| Se LD-Cu HD | 105.6 c | 1.49 b | 40.0 b | 6.53 a | Nd | |
| Se HD-Cu LD | 137.9 b | 2.25 ab | 45.9 b | 7.53 a | Nd | |
| Se HD-Cu HD | 137.9 b | 2.79 a | 51.3 ab | 7.25 b | Nd | |
| CV | 16.64 | 37.79 | 25.30 | 17.82 | - | |
| Fruit | Control | 16.6 b | 0.36 ab | 16.1 b | 5.29 b | 36.0 a |
| A. solani | 16.7 b | 0.31 b | 17.6 b | 5.32 b | 41.7 a | |
| Se LD-Cu LD | 19.7 a | 0.36 ab | 19.2 ab | 6.41 ab | 38.8 a | |
| Se LD-Cu HD | 20.7 a | 036 ab | 18.6 ab | 6.28 ab | 38.3 a | |
| Se HD-Cu LD | 15.8 b | 0.37 a | 18.2 b | 5.41 b | 37.1 a | |
| Se HD-Cu HD | 20.4 a | 0.39 a | 22.3 a | 6.76 a | 34.4 a | |
| CV | 10.25 | 12.32 | 17.23 | 19.24 | 23.72 |
| Organ | Treatment | APX (U g−1 TP) | GPX (U g−1 TP) | CAT (U g−1 TP) | SOD (U mL−1) | PAL (U g−1 TP) |
|---|---|---|---|---|---|---|
| Leaves | Control | 0.26 c | 10.40 b | 14.97 a | 1.52 a | 2.62 b |
| A. solani | 0.26 c | 15.12 ab | 16.80 a | 2.12 a | 4.35 b | |
| Se LD-Cu LD | 0.50 c | 30.02 ab | 36.35 a | 1.57 a | 7.48 ab | |
| Se LD-Cu HD | 0.62 bc | 19.99 ab | 20.67 a | 1.88 a | 4.81 ab | |
| Se HD-Cu LD | 1.16 ab | 29.53 ab | 37.27 a | 2.15 a | 6.88 ab | |
| Se HD-Cu HD | 1.39 a | 35.29 a | 15.72 a | 2.05 a | 9.91 a | |
| CV | 80.00 | 77.08 | 91.36 | 34.33 | 76.58 | |
| Fruit | Control | 10.32 b | 46.54 a | 2.12 a | 9.04 a | 2.51 b |
| A. solani | 37.00 ab | 113.23 a | 2.09 a | 19.45 a | 6.38 a | |
| Se LD-Cu LD | 34.08 ab | 89.82 a | 2.32 a | 16.34 a | 4.36 ab | |
| Se LD-Cu HD | 35.39 ab | 65.48 a | 2.01 a | 11.57 a | 2.40 b | |
| Se HD-Cu LD | 50.24 a | 94.73 a | 2.05 a | 16.47 a | 4.35 ab | |
| Se HD-Cu HD | 54.37 a | 66.60 a | 1.69 a | 14.56 a | 4.11 ab | |
| CV | 62.06 | 76.61 | 34.75 | 60.72 | 48.83 |
| Organ | Treatment | ABTS (µmol g−1 DW) | DPPH (µmol g−1 DW) | TAC ABTS (µmol g−1 DW) | TAC DPPH (µmol g−1 DW) | ||
|---|---|---|---|---|---|---|---|
| H | L | H | L | ||||
| Leaves | Control | 2.22 b | 1.56 bc | 1.44 d | 2.19 a | 3.78 bc | 3.64 bc |
| A. solani | 2.31 ab | 1.87 ab | 1.87 abc | 2.13 a | 4.18 abc | 4.00 abc | |
| Se LD-Cu LD | 2.45 ab | 1.87 ab | 1.58 cd | 2.12 a | 4.32 a | 3.69 bc | |
| Se LD-Cu HD | 2.19 b | 1.63 c | 1.69 bcd | 1.84 a | 3.71 c | 3.54 c | |
| Se HD-Cu LD | 2.27 ab | 2.00 a | 2.08 ab | 2.06 a | 4.27 ab | 4.14 ab | |
| Se HD-Cu HD | 2.61 a | 1.84 abc | 2.27 a | 2.20 a | 4.44 a | 4.47 a | |
| CV | 13.50 | 15.62 | 18.71 | 15.65 | 10.58 | 11.28 | |
| Fruit | Control | 3.18 a | 1.45 ab | 1.85 abc | 2.24 ab | 4.63 a | 4.08 ab |
| A. solani | 2.83 ab | 1.47 a | 1.96 ab | 2.03 b | 4.29 abc | 4.00 abc | |
| Se LD-Cu LD | 3.00 ab | 1.46 a | 1.63 bc | 2.32 a | 4.46 ab | 3.94 abc | |
| Se LD-Cu HD | 2.75 b | 1.29 c | 1.56 c | 2.07 b | 4.04 c | 3.63 c | |
| Se HD-Cu LD | 2.88 ab | 1.31 c | 2.01 a | 2.17 ab | 4.19 bc | 4.17 a | |
| Se HD-Cu HD | 2.94 ab | 1.32 bc | 1.64 bc | 2.05 b | 4.26 abc | 3.69 bc | |
| CV | 11.18 | 8.26 | 16.47 | 8.26 | 7.91 | 8.68 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiterio-Gutiérrez, T.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Hernández-Fuentes, A.D.; Sandoval-Rangel, A.; Benavides-Mendoza, A.; Cabrera-de la Fuente, M.; Juárez-Maldonado, A. The Application of Selenium and Copper Nanoparticles Modifies the Biochemical Responses of Tomato Plants under Stress by Alternaria solani. Int. J. Mol. Sci. 2019, 20, 1950. https://doi.org/10.3390/ijms20081950
Quiterio-Gutiérrez T, Ortega-Ortiz H, Cadenas-Pliego G, Hernández-Fuentes AD, Sandoval-Rangel A, Benavides-Mendoza A, Cabrera-de la Fuente M, Juárez-Maldonado A. The Application of Selenium and Copper Nanoparticles Modifies the Biochemical Responses of Tomato Plants under Stress by Alternaria solani. International Journal of Molecular Sciences. 2019; 20(8):1950. https://doi.org/10.3390/ijms20081950
Chicago/Turabian StyleQuiterio-Gutiérrez, Tomasa, Hortensia Ortega-Ortiz, Gregorio Cadenas-Pliego, Alma Delia Hernández-Fuentes, Alberto Sandoval-Rangel, Adalberto Benavides-Mendoza, Marcelino Cabrera-de la Fuente, and Antonio Juárez-Maldonado. 2019. "The Application of Selenium and Copper Nanoparticles Modifies the Biochemical Responses of Tomato Plants under Stress by Alternaria solani" International Journal of Molecular Sciences 20, no. 8: 1950. https://doi.org/10.3390/ijms20081950