LncRNA HOTAIR in Tumor Microenvironment: What Role?
by
Gerardo Botti
1, 
Giosuè Scognamiglio
2,
Gabriella Aquino
2,
Giuseppina Liguori
2 and
Monica Cantile
2,* 1
Scientific Direction, Istituto Nazionale Tumori, Fondazione G. Pascale, IRCCS, 80131 Naples, Italy
2
Pathology Unit, Istituto Nazionale Tumori, Fondazione G. Pascale, IRCCS, 80131 Naples, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(9), 2279; https://doi.org/10.3390/ijms20092279
Submission received: 9 April 2019
/
Revised: 3 May 2019
/
Accepted: 8 May 2019
/
Published: 8 May 2019
(This article belongs to the Special Issue Tumor Microenvironment 2019)
{kind=link}
{kind=link}
Abstract
:lncRNAs participate in many cellular processes, including regulation of gene expression at the transcriptional and post-transcriptional levels. In addition, many lncRNAs can contribute to the development of different human diseases including cancer. The tumor microenvironment (TME) plays an important role during tumor growth and metastatic progression, and most of these lncRNAs have a key function in TME intracellular signaling. Among the numerous identified lncRNAs, several experimental evidences have shown the fundamental role of the lncRNA HOTAIR in carcinogenesis, also highlighting its use as a circulating biomarker. In this review we described the contribution of HOTAIR in the TME modulation, highlighting its relation with cellular and non-cellular components during tumor evolution and progression.
1. Introduction
Long non-coding RNAs (lncRNAs) represent a large class of transcribed RNA molecules with a length of more than 200 nucleotides. Like other protein coding genes, they are probably transcribed by RNA polymerase II (RNA pol II), and are polyadenylated [1]. Generally, expression levels of lncRNAs appear to be lower than protein-coding genes [2] and specific to the tissues [3]. Although the function of most of the lncRNAs is unknown, an increasing number of them have been functionally characterized. LncRNAs have been involved in many biological activities such as; (i) the regulation of mRNA processing; (ii) the regulation of transcription, functioning as cis- and trans-acting modulators of protein-coding gene expression; (iii) the modulation of post-transcriptional control; (iv) the regulation of protein activity; (v) the production of scaffolds for higher-order complexes, such as Polycomb repressive complex 2 (PRC2) [4]. LncRNAs also represent an emerging class of tumor biomarkers being widely documented as to their deregulation in cancer development and progression [5].
Although most of the studies in the literature have shown the role of lncRNAs in tumor cells, a series of recent experimental evidences have suggested investigating their role in tumor microenvironment (TME) [6]. LncRNAs, as well as small noncoding RNAs, are transported in the circulation by extracellular vesicles, mainly exosomes [7]. These play a fundamental role in the interaction between cancer cells and the surrounding environment during tumor progression [8]. In fact, exosomal shuttle RNAs (exRNAs) might be used as a signal to change gene expression patterns in the recipient cell of TME [9,10]. Emerging data suggested that, during tumor disease progression, lncRNAs are involved in tumor-stroma signaling, regulating the phenotype of cancer-associated stromal cells. Stromal lncRNAs are able to induce the release of several chemokines enhancing the metastatic potential of tumor cells [11]. LncRNAs can also influence cancer stem cells (CSC) phenotype and differentiation status within the TME promoting tumor growth, invasion and metastatic progression [12]. In addition, some studies have shown than lncRNAs may be involved both in mechanisms of communication between metastatic cancer cells and extracellular matrix (ECM) and in ECM turnover [13].
Many lncRNAs are also expressed in immune cells, and appear to play critical roles in the regulation of immune response [14]. In animal models, lncRNAs are implicated in T cell activation/differentiation [15], and are able to modulate cytotoxicity CD8+ T cells recruitment within the TME, promoting metastatic disease. [16].
LncRNA HOTAIR (HOX Transcript Antisense Intergenic RNA) located on chr.12q13.13 (between HOXC11 and HOXC12) [17] acts as a regulator of chromatin states by binding, with its 5′end, the PRC2. With this mechanism HOTAIR can guide PRC2 to target genes related with tumor metastasis in breast cancer [18]. HOTAIR further interacts with the methyltransferase specific to histone 3 lysine 27, Enhancer of Zeste homolog 2 (EZH2), and lysine-specific demethylase 1 (LSD1), a central player in epigenetic regulation, through its 5′ end and 3′ end, respectively. Through this mechanism, HOTAIR can function as a scaffold directing these proteins to co-occupy the same genomic regions [19]. The same scaffold is able to link E3 ubiquitin ligases with their substrates to accelerate proteolysis [20]. A recent study showed that HOTAIR can inhibit Androgen Receptor (AR) ubiquitination, preventing AR protein degradation, and increasing AR protein stability. In fact, HOTAIR is able to bind the N-Terminal Domain (NTD) of the AR protein blocking recruitment of the E3 ubiquitin ligase MDM2 that interacts with AR through the same domain [21].
Summing up the experimental evidences, HOTAIR is able to: (i) promote the epigenetic activation/repression of gene expression; (ii) affect the target suppression of gene expression through competitive binding to miRNAs; (iii) modify gene expression, at post-transcriptional level, interacting with transcription factors and ribosomes or binding to splicing factors [22].
Aberrant HOTAIR expression has been detected in several human cancers showing a fundamental role in tumor proliferation, angiogenesis, progression, drug resistance and worse prognosis [23]. In addition, numerous experimental evidences have focused the attention on the potential role of HOTAIR as a circulating marker and therapeutic target for cancer patients [24].
This review aimed to highlight the potential role of HOTAIR in the modulation of TME, focusing attention in particular on its relation with cellular components of TME and ECM proteins. HOTAIR, through interaction with microenvironment elements, could play a main role in the activation of specific growth and invasion molecular pathways, and in the regulation of immune response during tumor progression.
2. Tumor Microenvironment Cells
The tumor microenvironment consists of different cellular and non-cellular secreted components. Cellular components include cancer-associated fibroblasts (CAF), endothelial cells, immune cells (T-cells, tumor associated macrophages (TAM), dendritic cells, mast cells etc.) and cancer stem cells (CSC) [25]. Secreted non cellular components comprise cytokines, growth factors, metabolites and extracellular matrix (ECM) proteins [26]. All TME elements play fundamental roles in tumor growth and progression [25].
2.1. HOTAIR and Cancer-Associated Fibroblasts
Fibroblasts, the predominant stromal cell type in the TME, are activated by tumor cells into cancer-associated fibroblasts (CAFs) through the secretion of paracrine growth factors, cytokines, and ECM proteins [26]. CAFs are important mediators of tumor-stroma communication, and are able, directly or indirectly, to promote different biological processes, such as proliferation, invasion, angiogenesis and drug resistance [27].
Although the role of lncRNAs in CAF modulation is poorly investigated, some studies suggested that they can contribute to: i) CAFs phenotype and function; ii) induction of epithelial-mesenchymal transition (EMT) in tumor cells, mainly promoting TGF-β signaling [28].
A recent study highlighted that breast cancer (BC) cell lines (Estrogen positive (ER+) and Triple Negative (TNBC)), grown in CAFs conditioned media (CAF-CM) lead the production of abundant levels of TGF-B1 cytokine inducing EMT of BC cells. To analyze the epigenetic mechanism by which CAFs induce EMT, several lncRNAs have been measured in BC cell lines revealing the up-regulation of HOTAIR in CAF-CM.
In addition, knock-down of HOTAIR in cancer cells is able to induce E-cadherin expression and vimentin and beta-catenin repression, suggesting that CAFs promote EMT and metastasis by activating HOTAIR expression. Knockdown of HOTAIR in orthotropic mice is able to attenuate the metastatic potential of tumor cells, blocking the crosstalk between CAFs and cancer cells [29]. However, the mechanisms by which CAFs activate HOTAIR remain to be addressed. TGF-β is able to assemble a receptor complex that activates Smad proteins (SMAD2, SMAD3 and SMAD4) regulating transcription [30]. In CAF-CM of BC cells, high levels of phosphorylated Smads proteins have been detected. The induced repression of these proteins leads to a 50–60% reduction in HOTAIR expression. CAFs could be able to stimulate the direct binding of SMAD2, 3, and 4 to the promoter region of HOTAIR inducing its transactivation [29]. In addition, since the activation of CDK5 is essential for CAF-induced EMT, Ren and collaborators also demonstrated that CAFs mediate HOTAIR expression and EMT by targeting CDK5 signaling [29]. These observations suggest a novel epigenetic mechanism by which CAFs would modulate EMT and tumor progression by TGF-ß1/CDK5/HOTAIR axis.
2.2. HOTAIR and Endothelial Cells
TME endothelial cells are responsible for supporting tumor neovasculature, also participating in several molecular signaling pathways. In fact, tumor-associated endothelial cells display high TGF-beta1 and CD105 expression involved in promoting angiogenesis [31].
Several lncRNAs are described as able to directly or indirectly modulate neo-angiogenesis in TME [32], but between these, HOTAIR appears to have a fundamental role in the promotion of angiogenesis during tumor progression. In nasopharyngeal carcinoma (NPC) HOTAIR is strongly up-regulated in cell lines and tissues, and its knockdown leads to a significant decrease of several angiogenic factors, such as vascular endothelial growth factor A (VEGFA) and angiopoietin 2 (Ang2) [33]. These data have been also been validated in animal xenograft, suggesting the powerful evidence that silencing of HOTAIR can act as an anti-angiogenesis agent to NPC carcinogenesis. HOTAIR is directly involved in the promotion of VEGFA transcription. In fact, the results of luciferase assays revealed that HOTAR activated the transcription of 2.3 kb VEGFA promoter. Moreover, a functional proteomic profiling identified 14 proteins up-regulated and 29 proteins down-regulated by HOTAIR silencing in NPC cells. Between these, Glucose regulated protein 78 (GRP78), belonging to the heat shock protein 70 (HSP70) family [34], has been validated as an anti-angiogenetic target of HOTAIR in NPC cells [33].
HOTAIR is also involved in the angiogenesis modulation of glioma cells. Its downregulation leads to the inhibition of pro-angiogenic activity in in vitro models. Also in this case, the silencing of HOTAIR in glioma cells dramatically decreases VEGFA expression affecting proliferation, migration and tube formation [35]. Experimental evidences showed that cancer cells are able to secrete extracellular vesicles (EV) to support tumor progression [36]. Numerous lncRNAs, including HOTAIR, are found in EVs derived from different cancer cells [37]. Ma and collaborators through RT-PCR analysis confirmed the presence of HOTAIR in conditioned medium of glioma cells and experiments using detergents, and RNAse indicated that HOTAIR remains stable because it is protected by membranes [35]. These observations suggested that HOTAIR, produced by tumor cells, would be able to be released in TME through EVs and transmitted to endothelial cells promoting angiogenesis.
In a non-neoplastic model, a recent study showed that HOTAIR is down-regulated in endothelial cells isolated from atherosclerotic plaque in atherosclerosis patients compared with corresponding vascular wall, suggesting that HOTAIR plays a protective role for endothelial cells injury [38]. Moreover, serum levels of thymic stromal lymphopoietin (TSLP), belonging to IL-7 like cytokine family, decrease in atherosclerosis patients, and positively correlates with HOTAIR expression in endothelial cells. TSLP is able to activate HOTAIR transcription through the PI3K/AKT pathway regulating endothelial cells proliferation and migration [38].
2.3. HOTAIR and Cancer Stem Cells
Cancer stem cells (CSCs) can be a critical component of TME being strongly involved in promoting invasion/metastasis process and drug resistance [39,40]. CSC proliferation can be regulated by many extrinsic factors derived from microenvironment cells. Several studies described that different lncRNAs are able to modulate CSCs proliferation promoting tumor progression [41]. Li et al., showed that HOTAIR is able to accelerate liver cancer stem cells proliferation in vitro and in vivo, on xenograft mice, through downregulation of the histone methyltransferase SET Domain-Containing Protein 2 (SETD2). The SETD2 gene has been shown to play a tumor suppressor role in human cancer [42]. In fact, cells lacking SETD2 display microsatellite instability (MSI) and an elevated spontaneous mutation frequency [43]. HOTAIR is able to reduce the SETD2 transcriptional expression through blocking RNApolII catalytic function by dissociating the CREB-P300- RNApolII complex. Therefore, SETD2 represents a crucial target of HOTAIR involved in epigenetic DNA damage repair, abnormal expression of cell cycle-related genes and microsatellite instability (MSI) [44].
HOTAIR appears as a key modulator of breast CSCs being strongly upregulated in BC CSCs models. HOTAIR is able to modulate proliferation, colony formation, migration and self-renewal capacity by negatively regulating miR34a [45]. The searching of the functional region of HOTAIR involved in this regulation showed that the full-length HOTAIR sequence is required for its regulatory role. Since miR-34a directly targets the stem cell marker Sox2, reducing its protein level [46], its mRNA expression has been also evaluated in BC CSCs. Sox2 appears strongly upregulated in BC CSCs, highlighting its indirect modulation by HOTAIR. In conclusion, HOTAIR might play a critical role in maintaining CSC self-renewal capacity by negatively regulating miR-34a and, consequently, positively Sox2 [45]. In non-small cell lung cancer (NSLC) patients, high HOTAIR expression is associated with drug resistance. In in vitro models, drug resistance induced by elevated HOTAIR expression may be caused by the promotion of tumor sphere cell growth and activation of tumor stem cell biomarkers such as Nanog, Oct3/4, Sox2, c-Myc, β-catenin, and Klf4 [47]. In oral squamous cell carcinoma (OSCC), overexpression of HOTAIR is tightly associated with the metastatic features of tumor cells, and is able to enhance cancer stemness. On the contrary, silencing of HOTAIR strongly attenuates oncogenic and invasion potential in xenograft nude mice. In addition, HOTAIR downregulation leads to the decreased expression of mesenchymal-like markers (vimentin, FN1, Snail, Twist and ZEB1) and increased expression of epithelial protein (E-cadherin) in OCSC cells, highlighting its main role of HOTAIR in the modulation of EMT features [48].
The role of HOTAIR in the modulation of stem-cell phenotypes has been also investigated in normal conditions. Exosomal HOTAIR released from gluteal-femoral fat might be able to promote intestinal cells proliferation, inducing both the stem cell and proliferation markers, such as several Wnt pathway-related genes (Lgr5, Cyclin d1, cMyc) [49]. HOTAIR, together with miR-10b, are involved in the malignant transformation of normal liver stem cells. Induced expression of both ncRNAs leads to the loss of E-cadherin and promotes EMT [46]. In bone marrow derived mesenchymal stem cells (MSCs) the overexpression and knockdown of HOTAIR interferes with cell differentiation and modulate senescence-associated changes in gene expression and DNA methylation. Targeting of HOTAIR to specific sites in the genome seems to be mediated by triple helix formation [50].
2.4. HOTAIR and Hypoxic Microenvironment
One of the most investigated features of TME is hypoxia, defined as a reduction in the normal level of tissue oxygen tension [51]. The hypoxic TME is involved in many oncogenic mechanisms, such as decreased DNA repair, increased mutation rate and chromosomal instability [52], cell proliferation signaling [51], invasion and metastasis [53], angiogenesis [54], inflammation and immunity [55] and reduced sensitivity to radiotherapy, chemotherapy and immunotherapy [56].
Several experimental evidences highlighted an increased production of exosomes in response to hypoxia [57,58] and oxidative stress [59]. In addition, many studies have demonstrated the aberrant expression of several non-coding RNAs under hypoxic condition.
Both microRNA and lncRNA in hypoxic microenvironment are able to drive the selection of a stem-like and more aggressive cancer cell population [60,61]. In particular, hypoxia-responsive lncRNAs (HRLs) play crucial roles in regulating hypoxic gene expression by acting as effectors of the indirect response to the transcription factor HIF (hypoxia-inducible factor) or direct modulators of the HIF-transcriptional cascade [62].
Different studies describe the role of HOTAIR in the hypoxic microenvironment of human cancers. HOTAIR is upregulated by hypoxia in NSCLC cells, and it is able to directly interact with HIF-1α through putative hypoxia-responsive elements (HREs) in the promoter region. The binding of HIF-1α to the HOTAIR promoter in vivo lead the increase of cancer cell proliferation, migration, and invasion [63]. In renal cell carcinoma (Rcc) HOTAIR is able to promote tumor progression through inhibition of miR-217 expression, and modulating the expression of HIF-1α and AXL receptor tyrosine kinase. In addition, HOTAIR knockdown significantly suppresses tumor growth and ki-67 expression, increases miR-217 levels and decreases the expression of HIF-1α and AXL, inhibiting tumor growth and EMT [64]. More recent studies showed that HOTAIR can be transcriptionally induced upon hypoxic conditions also in other tumor types, including colon cancer, breast cancer, cervical cancer, and neuroblastoma cells [65]. While HOTAIR expression is induced upon hypoxia, several HOTAIR target genes and tumor suppressors such as HOXD10, HOXD8, PCDHGA8, and PCDHB5 are down-regulated under hypoxic conditions. This mechanism of silencing could be related to the HOTAIR recruitment of PRC2 and LSD1 complexes which introduce H3K27-trimethylation and H3K4-demethylation respectively at the above target gene promoters [65]. Through its interaction with HIF-1a, HOTAIR is also able to induce radio-resistance in cervical cancer cells, suggesting that HOTAIR-HIF-1α axis might be a crucial mechanism for radio-resistance modulation hypoxia-induced [66].
2.5. HOTAIR and Immune Cells
Immune cells represent the most abundant cellular component within TME. In particular, T-cells account for about 50% of intratumoral cells playing a main role in influencing antitumor immunity in both positive and negative ways, depending upon the cell type [67].
Tumor-associated macrophages (TAM) cells play a fundamental role in the inhibition of antitumor immunity by secreting suppressive cytokines, leading to an increase of angiogenic process and promoting tumor growth [68]. At same manner, an increased presence of myeloid-derived suppressor cells (MDSC) are strongly associated with tumor progression and poorer outcome [69].
LncRNAs play fundamental roles in immune cell development, lineage differentiation and effector function [70]. However, information on the role of HOTAIR in immune cells modulation within TME is limited. Hepatoma cell lines with HOTAIR overexpression secreted higher CCL2 and this promotes TAM and MDSCs proliferation. CCL2 is required for TAM and MDSCs recruitment in the tumor microenvironment, suggesting that HOTAIR has a main role in the modulation of this process promoting tumor growth and metastasis [71]. The analysis of a correlation between CCL2 and HOTAIR expression in Gene Expression Omnibus (GEO) data set of HCC, revealed that patients with high expression levels of both markers are associated with advanced clinical stages, suggesting that HOTAIR could be a specific target for immunotherapy against CCL2 [67]. Prostate cancer cells treated with androgen deprivation therapy strongly increase the recruitment of mast cells. In turn, infiltrating mast cells are able to decrease AR transcription through HOTAIR activation increasing cell invasion via MMP expression induction [72].
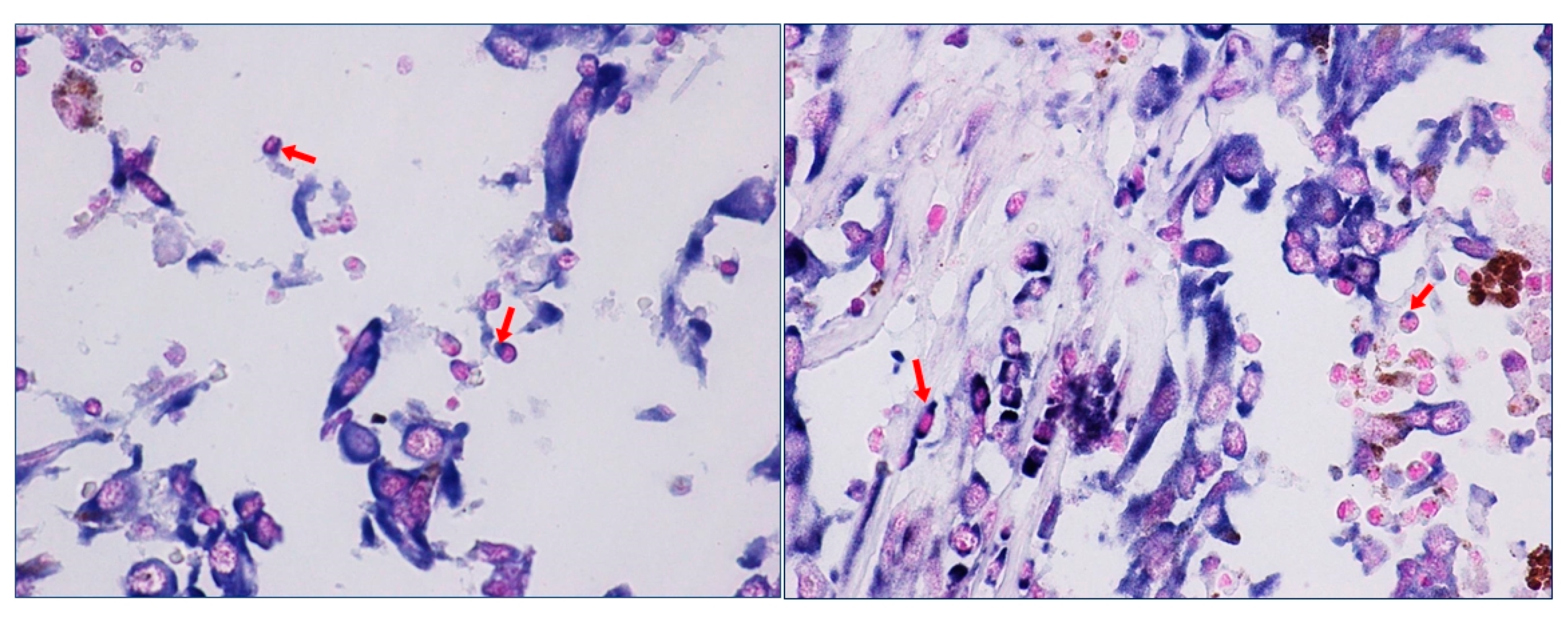
We have recently highlighted the aberrant expression of HOTAIR in metastatic melanoma patients and shown its high expression in lymphocytes surrounding metastatic tumor cells [73]. In our study we clearly showed the in situ expression of HOTAIR on immune cells surface within TME. The images of the staining at higher magnification highlight the details of HOTAIR staining on the plasma membrane of lymphocytes (Figure 1 (source: ref 74)). HOTAIR expression would appear to be associated with specific “vesicle-like” membrane projections.
However, it is not clear whether the presence of HOTAIR on intra-tumor lymphocytes of patients with metastatic melanoma is associated with endogenous production of the lncRNA, or it is the result of a signal transmission between tumor cells and TME cells.
HOTAIR expression can be induced in normal immune cells upon treatment with lipopolysaccharide (LPS), an endotoxin able to promote immune and pro-inflammatory response in macrophages. In turn, HOTAIR is able to promote several cytokines and inflammatory genes, such as IL-6, iNOS, TNFα, and MIP-1B, highlighting its critical role in immune response upon LPS-stimulation. HOTAIR is also required for LPS-induced degradation of IκBα (nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha) and activation of NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) with its nuclear translocation [74]. It is known that NF-κB activation in immune cells enhances inflammation in the TME by increasing the secretion of pro-inflammatory cytokines such as TNF-α and IL-6, and promotes the rapid tumor cells proliferation [75].
2.6. HOTAIR and Extracellular Matrix
Although extracellular matrix constitutes the non-cellular component of TME, it represents a dynamic and versatile structure able to influence fundamental aspects of tumor cell biology [76]. In fact, during cancer progression, overcoming the physical barriers through the interaction with ECM components is a fundamental step of the invasion/metastatic process. Several studies highlighted an important role of lncRNAs in ECM organization and remodeling [77].
HOTAIR has an important function in ECM signaling in breast cancer, above all in invasive “claudin low” molecular subtype. BC cells grown in a 3D culture on a substrate enriched in laminin show a significant up-regulation of HOTAIR expression. Not all the different isoforms of HOTAIR have been characterized, even if the aberrant activation of the promoter region upstream of HOTAIR-N has been mainly implied in invasive breast cancer [78]. Li et al. showed that a novel isoform HOTAIR-N is activated in 3D culture of Claudin-low BC cells enriched in laminin. HOTAIR expression is induced by increased H3K4me3 and BRD4 binding to the novel HOTAIR-N promoter in BC cells attached to ECM, suggesting its main role in ECM organization and tumor invasion pathway signaling [79].
The interplay between ECM components and HOTAIR has been also demonstrated in lung cancer cells. The enrichment of collagen I in a 3D culture model determines the loss of morphological features of well-differentiated lung adenocarcinoma and the up-regulation of HOTAIR expression. On the contrary, the treatment of lung cancer cells with an antibody against collagen I receptor lead to a decrease of HOTAIR expression, suggesting that HOTAIR expression in tumor tissues is the result of tumor cell response to collagen I, abundantly present in TME [80].
Osteopontin is another component of ECM associated with several biological processes, such as angiogenesis, inflammation, tissues remodeling, but also metastatic progression. In fact, OPN increases pro-MMP2 expression in an NFKb-dependent manner during ECM invasion [81]. OPN is able to induce HOTAIR expression in a dose and time-dependent manner in different tumor cell lines, while OPN knockdown decreases HOTAIR expression. Activation of the PI3K/AKT pathway seems to be associated with the effect of OPN on the regulation of HOTAIR. Moreover, the interferon regulatory factor IRF1, a transcriptional regulator of IFN-stimulated genes, might participate in the transcriptional regulation of HOTAIR. In fact, promoter activity and ChIP assays confirmed that IRF1 could bind to the HOTAIR promoter and inhibit HOTAIR expression [82].
3. Conclusions
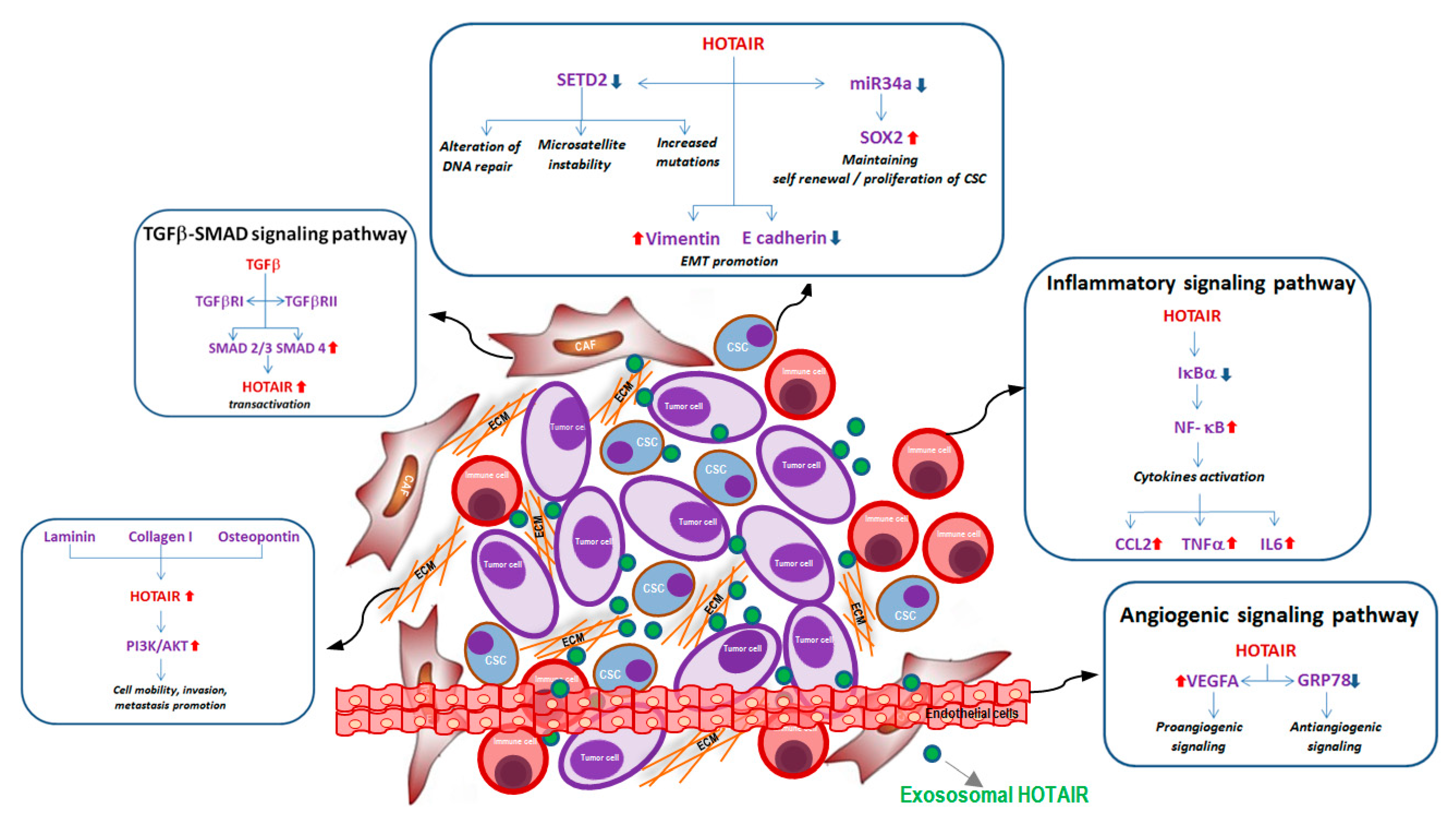
Although the role of lncRNAs within TME is still poorly investigated, a number of studies suggest the main contribution of HOTAIR in TME intracellular signaling and its important contribution in modulating different molecular pathways involved in tumor phenotype modifications during metastatic progression (Figure 2). The evaluation of HOTAIR expression in TME could provide fundamental information, not only for the prognosis, but also for the prediction of the therapeutic response of cancer patients.
Funding
This study was supported by the Italian Ministry of Health.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Noncoding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babak, T.; Blencowe, B.J.; Hughes, T.R. A systematic search for new mammalian noncoding RNAs indicates little conserved intergenic transcription. BMC Genomics. 2005, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Sunkin, S.M.; Mehler, M.F.; Mattick, J.S. Specific expression of long noncoding RNAs in the mouse brain. Proc. Natl. Acad. Sci. USA 2008, 105, 716–721. [Google Scholar] [CrossRef] [Green Version]
- St Laurent, G.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Wu, M.H.; Yeh, C.T.; Lin, K.H. Long Non-Coding RNAs as Mediators of Tumor Microenvironment and Liver Cancer Cell Communication. Int. J. Mol. Sci. 2018, 19, 3742. [Google Scholar] [CrossRef]
- Fatima, F.; Nawaz, M. Vesiculated Long Non-Coding RNAs: Offshore Packages Deciphering Trans-Regulation between Cells, Cancer Progression and Resistance to Therapies. Noncoding RNA. 2017, 3, 10. [Google Scholar] [CrossRef]
- Sun, Z.; Yang, S.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Xu, J.; Xia, K.; Chang, Y.; Liu, J.; Yuan, W. Emerging role of exosome-derived long non-coding RNAs in tumor microenvironment. Mol. Cancer. 2018, 17, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cell. Nat. Cell. Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, S.; Palanisamy, V. Horizontal transfer of RNAs: exosomes as mediators of intercellular communication. Wiley Interdiscip. Rev. RNA. 2012, 3, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Ji, G.; Le, X.; Wang, C.; Xu, L.; Feng, M.; Zhang, Y.; Yang, H.; Xuan, Y.; Yang, Y.; et al. Long noncoding RNA LINC00092 acts in cancer-associated fibroblasts to drive glycolysis and progression of ovarian cancer. Canc. Res. 2017, 77, 1369–1382. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.S.; Pelagalli, A.; Passaro, N.; Zannetti, A. Tumor-educated mesenchymal stem cells promote pro-metastatic phenotype. Oncotarget. 2017, 8, 73296–73311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.T.; Song, H.J.; Ding, X.Y. Long non-coding RNAs involved in metastasis of gastric cancer. World J. Gastroenterol. 2018, 24, 3724–3737. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Satpathy, A.T.; Chang, H.Y. Gene regulation in the immune system by long noncoding RNAs. Nat. Immunol. 2017, 18, 962–972. [Google Scholar] [CrossRef]
- Guo, Q.; Cheng, Y.; Liang, T.; He, Y.; Ren, C.; Sun, L.; Zhang, G. Comprehensive analysis of lncRNA-mRNA co-expression patterns identifies immune-associated lncRNA biomarkers in ovarian cancer malignant progression. Sci. Rep. 2015, 5, 17683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Zhao, Z.; Liu, K.; Zhang, J.; Li, G.; Wang, L. Long noncoding RNA lnc-sox5 modulates CRC tumorigenesis by unbalancing tumor microenvironment. Cell Cycle. 2017, 16, 1295–1301. [Google Scholar] [CrossRef]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; Chang, H.Y. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef]
- Yoon, J.H.; Abdelmohsen, K.; Kim, J.; Yang, X.; Martindale, J.L.; Tominaga-Yamanaka, K.; White, E.J.; Orjalo, A.V.; Rinn, J.L.; Kreft, S.G.; et al. Scaffold function of long non-coding RNA HOTAIR in protein ubiquitination. Nat. Commun. 2013, 4, 2939. [Google Scholar] [CrossRef]
- Zhang, A.; Zhao, J.C.; Kim, J.; Fong, K.W.; Yang, Y.A.; Chakravarti, D.; Mo, Y.Y.; Yu, J. LncRNA HOTAIR Enhances the Androgen-Receptor-Mediated Transcriptional Program and Drives Castration-Resistant Prostate Cancer. Cell Rep. 2015, 13, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, C.J.; Kingston, R.E. HOTAIR lifts noncoding RNAs to new levels. Cell 2007, 129, 1257–1259. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Hann, S.S. HOTAIR: An Oncogenic Long Non-Coding RNA in Human Cancer. Cell. Physiol. Biochem. 2018, 47, 893–913. [Google Scholar] [CrossRef]
- Botti, G.; Marra, L.; Malzone, M.G.; Anniciello, A.; Botti, C.; Franco, R.; Cantile, M. LncRNA HOTAIR as Prognostic Circulating Marker and Potential Therapeutic Target in Patients with Tumor Diseases. Curr. Drug Targets. 2017, 18, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [Green Version]
- Ringuette-Goulet, C.; Bernard, G.; Tremblay, S.; Chabaud, S.; Bolduc, S.; Pouliot, F. Exosomes induce fibroblast differentiation into Cancer-associated fibroblasts through TGFβ signaling. Mol. Cancer Res. Am. Assoc. Cancer Res. 2018, 16, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Erez, N.; Truitt, M.; Olson, P.; Arron, S.T.; Hanahan, D. Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-kappaB-dependent manner. Cancer Cell. 2010, 17, 135–147. [Google Scholar] [CrossRef]
- Calon, A. Tauriello DVF and Batlle E TGF-β in CAF-mediated tumor growth and metastasis. Semin Cancer Biol. 2014, 25, 15–22. [Google Scholar] [CrossRef]
- Ren, Y.; Jia, H.H.; Xu, Y.Q.; Zhou, X.; Zhao, X.H.; Wang, Y.F.; Song, X.; Zhu, Z.Y.; Sun, T.; Dou, Y.; et al. Paracrine and epigenetic control of CAF-induced metastasis: the role of HOTAIR stimulated by TGF-ß1 secretion. Mol. Cancer. 2018, 17, 5. [Google Scholar] [CrossRef] [Green Version]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef]
- Benetti, A.; Berenzi, A.; Gambarotti, M.; Garrafa, E.; Gelati, M.; Dessy, E.; Portolani, N.; Piardi, T.; Giulini, S.M.; Caruso, A.; et al. Transforming growth factor-beta1 and CD105 promote the migration of hepatocellular carcinoma-derived endothelium. Cancer Res. 2008, 68, 8626–8634. [Google Scholar] [CrossRef]
- Yu, B.; Wang, S. Angio-lncRs: lncRNAs that regulate angiogenesis and vascular disease. Theranostics 2018, 8, 3654–3675. [Google Scholar] [CrossRef]
- Fu, W.M.; Lu, Y.F.; Hu, B.G.; Liang, W.C.; Zhu, X.; Yang, H.D.; Li, G.; Zhang, J.F. Long noncoding RNA Hotair mediated angiogenesis in nasopharyngeal carcinoma by direct and indirect signaling pathways. Oncotarget 2016, 7, 4712–4723. [Google Scholar] [CrossRef]
- Li, Z.; Li, Z. Glucose regulated protein 78: a critical link between tumor microenvironment and cancer hallmarks. Biochim. Biophys Acta. 2012, 1826, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, Z.; Li, T.; Zhu, L.; Li, Z.; Tian, N. Long non-coding RNA HOTAIR enhances angiogenesis by induction of VEGFA expression in glioma cells and transmission to endothelial cells via glioma cell derived-extracellular vesicles. Am. J. Transl. Res. 2017, 9, 5012–5021. [Google Scholar] [PubMed]
- Godlewski, J.; Krichevsky, A.M.; Johnson, M.D.; Chiocca, E.A.; Bronisz, A. Belonging to a network--microRNAs, extracellular vesicles, and the glioblastoma microenvironment. Neuro Oncol. 2015, 17, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Gezer, U.; Özgür, E.; Cetinkaya, M.; Isin, M.; Dalay, N. Long non-coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol. Int. 2014, 38, 1076–1079. [Google Scholar] [CrossRef]
- Peng, Y.; Meng, K.; Jiang, L.; Zhong, Y.; Yang, Y.; Lan, Y.; Zeng, Q.; Cheng, L. Thymic stromal lymphopoietin-induced HOTAIR activation promotes endothelial cell proliferation and migration in atherosclerosis. Biosci. Rep. 2017, 37, BSR20170351. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Iwama, A.; Yokosuka, O. Cancer stem cells in hepatocellular carcinoma: Therapeutic implications based on stem cell biology. Hepatol. Res. 2016, 46, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Lin, M.; Jiang, X.; Ye, J.; Guo, T.; Shi, Y.; Bian, X. The Recent Advances on Liver Cancer Stem Cells: Biomarkers, Separation, and Therapy. Anal. Cell. Pathol. 2017, 2017, 5108653. [Google Scholar] [CrossRef]
- Chen, S.; Zhu, J.; Wang, F.; Guan, Z.; Ge, Y.; Yang, X.; Cai, J. LncRNAs and their role in cancer stem cells. Oncotarget 2017, 8, 110685–110692. [Google Scholar] [CrossRef] [Green Version]
- Al Sarakbi, W.; Sasi, W.; Jiang, W.G.; Roberts, T.; Newbold, R.F.; Mokbel, K. The mRNA expression of SETD2 in human breast cancer: correlation with clinico-pathological parameters correlation with clinico-pathological parameters. BMC Cancer. 2009, 9, 290. [Google Scholar] [CrossRef]
- Li, F.; Mao, G.; Tong, D.; Huang, J.; Gu, L.; Yang, W.; Li, G.M. The histone mark H3K36me3 regulates human DNA mismatch repair through its interaction with MutSα. Cell 2013, 153, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; An, J.; Wu, M.; Zheng, Q.; Gui, X.; Li, T.; Pu, H.; Lu, D. LncRNA HOTAIR promotes human liver cancer stem cell malignant growth through downregulation of SETD2. Oncotarget 2015, 6, 27847–27864. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Yang, M.; Jiang, R.; An, N.; Wang, X.; Liu, B. Long Non-Coding RNA HOTAIR Regulates the Proliferation, Self-Renewal Capacity, Tumor Formation and Migration of the Cancer Stem-Like Cell (CSC) Subpopulation Enriched from Breast Cancer Cells. PLoS ONE 2017, 12, e0170860. [Google Scholar] [CrossRef]
- Ye, P.; Wang, T.; Liu, W.H.; Li, X.C.; Tang, L.J.; Tian, F.Z. Enhancing HOTAIR/MiR-10b drives normal liver stem cells toward a tendency to malignant transformation through inducing epithelial- to-mesenchymal transition. Rejuvenat. Res. 2015, 18, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Li, X.Q.; Gao, T.H.; Cui, Y.; Ma, N.; Zhou, Y.; Zhang, G.J. Elevated HOTAIR expression associated with cisplatin resistance in non-small cell lung cancer patients. J. Thorac. Dis. 2016, 8, 3314–3322. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.Y.; Liao, Y.W.; Chen, P.Y.; Hsieh, P.L.; Fang, C.Y.; Wu, C.Y.; Yen, M.L.; Peng, B.Y.; Wang, D.P.; Cheng, H.C.; et al. Targeting LncRNA HOTAIR suppresses cancer stemness and metastasis in oral carcinomas stem cells through modulation of EMT. Oncotarget 2017, 8, 98542–98552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Bai, D.; Liu, X.; Zhou, C.; Yang, G. Sedentary lifestyle related exosomal release of Hotair from gluteal-femoral fat promotes intestinal cell proliferation. Sci. Rep. 2017, 7, 45648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalwa, M.; Hänzelmann, S.; Otto, S.; Kuo, C.C.; Franzen, J.; Joussen, S.; Fernandez-Rebollo, E.; Rath, B.; Koch, C.; Hofmann, A.; et al. The lncRNA HOTAIR impacts on mesenchymal stem cells via triple helix formation. Nucl. Acids Res. 2016, 44, 10631–10643. [Google Scholar] [CrossRef]
- Harris, A.L. Hypoxia--a key regulatory factor in tumour growth. Nat. Rev. Cancer. 2002, 2, 38–47. [Google Scholar] [CrossRef]
- Bristow, R.G.; Hill, R.P. Hypoxia and metabolism. Hypoxia, DNA repair and genetic instability. Nat. Rev. Cancer 2008, 8, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Rankin, E.B.; Giaccia, A.J. Hypoxic control of metastasis. Science 2016, 352, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, R.K. Antiangiogenesis strategies revisited: from starving tumors to alleviating hypoxia. Cancer Cell. 2014, 26, 605–622. [Google Scholar] [CrossRef] [PubMed]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF transcription factors, inflammation, and immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Graham, K.; Unger, E. Overcoming tumor hypoxia as a barrier to radiotherapy, chemotherapy and immunotherapy in cancer treatment. Int. J. Nanomed. 2018, 13, 6049–6058. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; Zhu, G. Exosomes derived from hypoxic Oral squamous cell carcinoma cells deliver miR-21 to normoxic cells to elicit a Prometastatic phenotype. Cancer Res. 2016, 76, 1770–1780. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Jayaprakash, P.; Dan, J.; Wise, P.; Jang, G.B.; Liang, C.; Chen, M.; Woodley, D.T.; Fabbri, M.; Li, W. PRAS40 connects microenvironmental stress signaling to exosome-mediated secretion. Mol. Cell. Biol. 2017, 37, e00171-17. [Google Scholar] [CrossRef]
- Hedlund, M.; Nagaeva, O.; Kargl, D.; Baranov, V.; Mincheva-Nilsson, L. Thermal- and oxidative stress causes enhanced release of NKG2D ligand-bearing immunosuppressive exosomes in leukemia/lymphoma T and B cells. PLoS ONE 2011, 6, e16899. [Google Scholar] [CrossRef]
- Macharia, L.W.; Wanjiru, C.M.; Mureithi, M.W.; Pereira, C.M.; Ferrer, V.P.; Moura-Neto, V. MicroRNAs, Hypoxia and the Stem-Like State as Contributors to Cancer Aggressiveness. Front. Genet. 2019, 10, 125. [Google Scholar] [CrossRef]
- Kapinova, A.; Kubatka, P.; Zubor, P.; Golubnitschaja, O.; Dankova, Z.; Uramova, S.; Pilchova, I.; Caprnda, M.; Opatrilova, R.; Richnavsky, J.; Kruzliak, P.; Danko, J. The hypoxia-responsive long non-coding RNAs may impact on the tumor biology and subsequent management of breast cancer. Biomed. Pharmacother. 2018, 99, 51–58. [Google Scholar] [CrossRef]
- Shih, J.W.; Kung, H.J. Long non-coding RNA and tumor hypoxia: new players ushered toward an old arena. J. Biomed. Sci. 2017, 24, 53. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Ye, L.; Jiang, C.; Bai, J.; Chi, Y.; Zhang, H. Long noncoding RNA HOTAIR, a hypoxia-inducible factor-1α activated driver of malignancy, enhances hypoxic cancer cell proliferation, migration, and invasion in non-small cell lung cancer. Tumour. Biol. 2015, 36, 9179–9188. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Li, O.; Zheng, W.; Xiao, W.Z.; Zhang, L.; Wu, D.; Cai, G.Y.; He, J.C.; Chen, X.M. LncRNA HOTAIR regulates HIF-1α/AXL signaling through inhibition of miR-217 in renal cell carcinoma. Cell Death Dis. 2017, 8, e2772. [Google Scholar] [CrossRef]
- Bhan, A.; Deb, P.; Shihabeddin, N.; Ansari, K.I.; Brotto, M.; Mandal, S.S. Histone methylase MLL1 coordinates with HIF and regulate lncRNA HOTAIR expression under hypoxia. Gene 2017, 629, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Meng, D.D.; Gao, L.; Xu, Y.; Liu, P.J.; Tian, Y.W.; Yi, Z.Y.; Zhang, Y.; Tie, X.J.; Xu, Z.Q. Overexpression of HOTAIR leads to radioresistance of human cervical cancer via promoting HIF-1α expression. Radiat. Oncol. 2018, 13, 210. [Google Scholar] [CrossRef] [PubMed]
- Becht, E.; Giraldo, N.A.; Germain, C.; de Reyniès, A.; Laurent-Puig, P.; Zucman-Rossi, J.; Dieu-Nosjean, M.C.; Sautès-Fridman, C.; Fridman, W.H. Immune Contexture, Immunoscore, and Malignant Cell Molecular Subgroups for Prognostic and Theranostic Classifications of Cancers. Adv. Immunol. 2016, 130, 95–190. [Google Scholar] [PubMed]
- Noy, R.; Pollard, J.W. Tumor-associated macrophages: from mechanisms to therapy. Immunity 2014, 41, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Safari, E.; Ghorghanlu, S.; Ahmadi-Khiavi, H.; Mehranfar, S.; Rezaei, R.; Motallebnezhad, M. Myeloid-derived suppressor cells and tumor: Current knowledge and future perspectives. J. Cell. Physiol. 2019, 234, 9966–9981. [Google Scholar] [CrossRef] [PubMed]
- Atianand, M.K.; Fitzgerald, K.A. Long non-coding RNAs and control of gene expression in the immune system. Trends Mol. Med. 2014, 20, 623–631. [Google Scholar] [CrossRef] [Green Version]
- Fujisaka, Y.; Iwata, T.; Tamai, K.; Nakamura, M.; Mochizuki, M.; Shibuya, R.; Yamaguchi, K.; Shimosegawa, T.; Satoh, K. Long non-coding RNA HOTAIR up-regulates chemokine (C-C motif) ligand 2 and promotes proliferation of macrophages and myeloid-derived suppressor cells in hepatocellular carcinoma cell lines. Oncol. Lett. 2018, 15, 509–514. [Google Scholar] [CrossRef]
- Li, L.; Dang, Q.; Xie, H.; Yang, Z.; He, D.; Liang, L.; Song, W.; Yeh, S.; Chang, C. Infiltrating mast cells enhance prostate cancer invasion via altering LncRNA-HOTAIR/PRC2-androgen receptor (AR)-MMP9 signals and increased stem/progenitor cell population. Oncotarget 2015, 6, 14179–14190. [Google Scholar] [CrossRef] [Green Version]
- Cantile, M.; Scognamiglio, G.; Marra, L.; Aquino, G.; Botti, C.; Falcone, M.R.; Malzone, M.G.; Liguori, G.; Di Bonito, M.; Franco, R.; et al. HOTAIR role in melanoma progression and its identification in the blood of patients with advanced disease. J. Cell. Physiol. 2017, 232, 3422–3432. [Google Scholar] [CrossRef]
- Obaid, M.; Udden, S.M.N.; Deb, P.; Shihabeddin, N.; Zaki, M.H.; Mandal, S.S. LncRNA HOTAIR regulates lipopolysaccharide-induced cytokine expression and inflammatory response in macrophages. Sci. Rep. 2018, 8, 15670. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Shen, S.; Verma, I.M. NF-κB, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, C.; Mojares, E.; Del Río Hernández, A. Role of Extracellular Matrix in Development and Cancer Progression. Int. J. Mol. Sci. 2018, 19, 3028. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Agostini, M. Long non-coding RNA and extracellular matrix: the hidden players in cancer-stroma cross-talk. Noncoding RNA Res. 2018, 3, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Milevskiy, M.J.; Al-Ejeh, F.; Saunus, J.M.; Northwood, K.S.; Bailey, P.J.; Betts, J.A.; McCart Reed, A.E.; Nephew, K.P.; Stone, A.; Gee, J.M. Long-range regulators of the lncRNA HOTAIR enhance its prognostic potential in breast cancer. Hum. Mol. Genet. 2016, 25, 3269–3283. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, X.; Zhuang, Y.; Flemington, E.K.; Lin, Z.; Shan, B. Induction of a novel isoform of the lncRNA HOTAIR in Claudin-low breast cancer cells attached to extracellular matrix. Mol Oncol. 2017, 11, 1698–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Y.; Wang, X.; Nguyen, H.T.; Zhuo, Y.; Cui, X.; Fewell, C.; Flemington, E.K.; Shan, B. Induction of long intergenic non-coding RNA HOTAIR in lung cancer cells by type I collagen. J. Hematol. Oncol. 2013, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Philip, S.; Bulbule, A.; Kundu, G.C. Osteopontin stimulates tumor growth and activation of promatrix metalloproteinase-2 through nuclear factor-kappa B-mediated induction of membrane type 1 matrix metalloproteinase in murine melanoma cells. J. Biol. Chem. 2001, 276, 44926–44935. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, S.; Gao, F.; Liu, Z.; Lu, M.; Peng, S.; Zhang, T.; Zhang, F. Osteopontin enhances the expression of HOTAIR in cancer cells via IRF1. Biochim. Biophys. Acta 2014, 1839, 837–848. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
HOTAIR expression in immune cells within tumor microenvironment of a melanoma sample. The red arrows indicate membrane projections in which HOTAIR staining is predominantly concentrated (magnification 40×).
Figure 1.
HOTAIR expression in immune cells within tumor microenvironment of a melanoma sample. The red arrows indicate membrane projections in which HOTAIR staining is predominantly concentrated (magnification 40×).

Figure 2.
Schematic representation of exosomal HOTAIR in tumor microenvironment with details of the main molecular pathways associated with cellular and non-cellular components: (i) In CAF cells: TGF-β assembling the receptor complex (TGFbRI-TGFbRII) activates Smad proteins (SMAD2, SMAD3 and SMAD4) that directly bind HOTAIR promoter, inducing its transactivation; (ii) In CSC cells: HOTAIR leads to (a) the downregulation of the tumor suppressor gene SETD2, promoting microsatellite instability, high mutation rate, and interfering with DNA damage repair, (b) the downregulation of miR34a with the consequent induction of stem cell marker Sox2; (iii) In endothelial cells: HOTAIR induces angiogenesis-promoting VEGFA transcription and downregulating the anti-angiogenetic marker GRP78; (iv) In immune cells: HOTAIR is able to induce the degradation of IκBα with the consequent activation of NF-κB pathways and secretion of pro-inflammatory cytokines; (v) ECM proteins: Laminin, Collagen I, Osteopontin are able to induce HOTAIR overexpression modulating PI3K/AKT pathways and promoting cell mobility and invasion.
Figure 2.
Schematic representation of exosomal HOTAIR in tumor microenvironment with details of the main molecular pathways associated with cellular and non-cellular components: (i) In CAF cells: TGF-β assembling the receptor complex (TGFbRI-TGFbRII) activates Smad proteins (SMAD2, SMAD3 and SMAD4) that directly bind HOTAIR promoter, inducing its transactivation; (ii) In CSC cells: HOTAIR leads to (a) the downregulation of the tumor suppressor gene SETD2, promoting microsatellite instability, high mutation rate, and interfering with DNA damage repair, (b) the downregulation of miR34a with the consequent induction of stem cell marker Sox2; (iii) In endothelial cells: HOTAIR induces angiogenesis-promoting VEGFA transcription and downregulating the anti-angiogenetic marker GRP78; (iv) In immune cells: HOTAIR is able to induce the degradation of IκBα with the consequent activation of NF-κB pathways and secretion of pro-inflammatory cytokines; (v) ECM proteins: Laminin, Collagen I, Osteopontin are able to induce HOTAIR overexpression modulating PI3K/AKT pathways and promoting cell mobility and invasion.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Botti, G.; Scognamiglio, G.; Aquino, G.; Liguori, G.; Cantile, M. LncRNA HOTAIR in Tumor Microenvironment: What Role? Int. J. Mol. Sci. 2019, 20, 2279. https://doi.org/10.3390/ijms20092279
AMA Style
Botti G, Scognamiglio G, Aquino G, Liguori G, Cantile M. LncRNA HOTAIR in Tumor Microenvironment: What Role? International Journal of Molecular Sciences. 2019; 20(9):2279. https://doi.org/10.3390/ijms20092279
Chicago/Turabian StyleBotti, Gerardo, Giosuè Scognamiglio, Gabriella Aquino, Giuseppina Liguori, and Monica Cantile. 2019. "LncRNA HOTAIR in Tumor Microenvironment: What Role?" International Journal of Molecular Sciences 20, no. 9: 2279. https://doi.org/10.3390/ijms20092279
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.