Immune Checkpoint Expression on Immune Cells of HNSCC Patients and Modulation by Chemo- and Immunotherapy


 , , ,
, , ,
Abstract
:1. Introduction
1.1. Tumor Microenvironment and Squamous Cell Carcinoma of the Head and Neck
1.2. Immune Checkpoint Inhibitors in Clinical Use
2. Results
2.1. Cohort Characteristics and PD-L1 Status of HNSCC Patients
2.2. Altered Checkpoint Expression on Peripheral Blood Lymphocytes (PBL) of Healthy Donors and HNSCC Patients
2.3. OX40 Upregulation on Treg of HPV-Positive HNSCC Patients
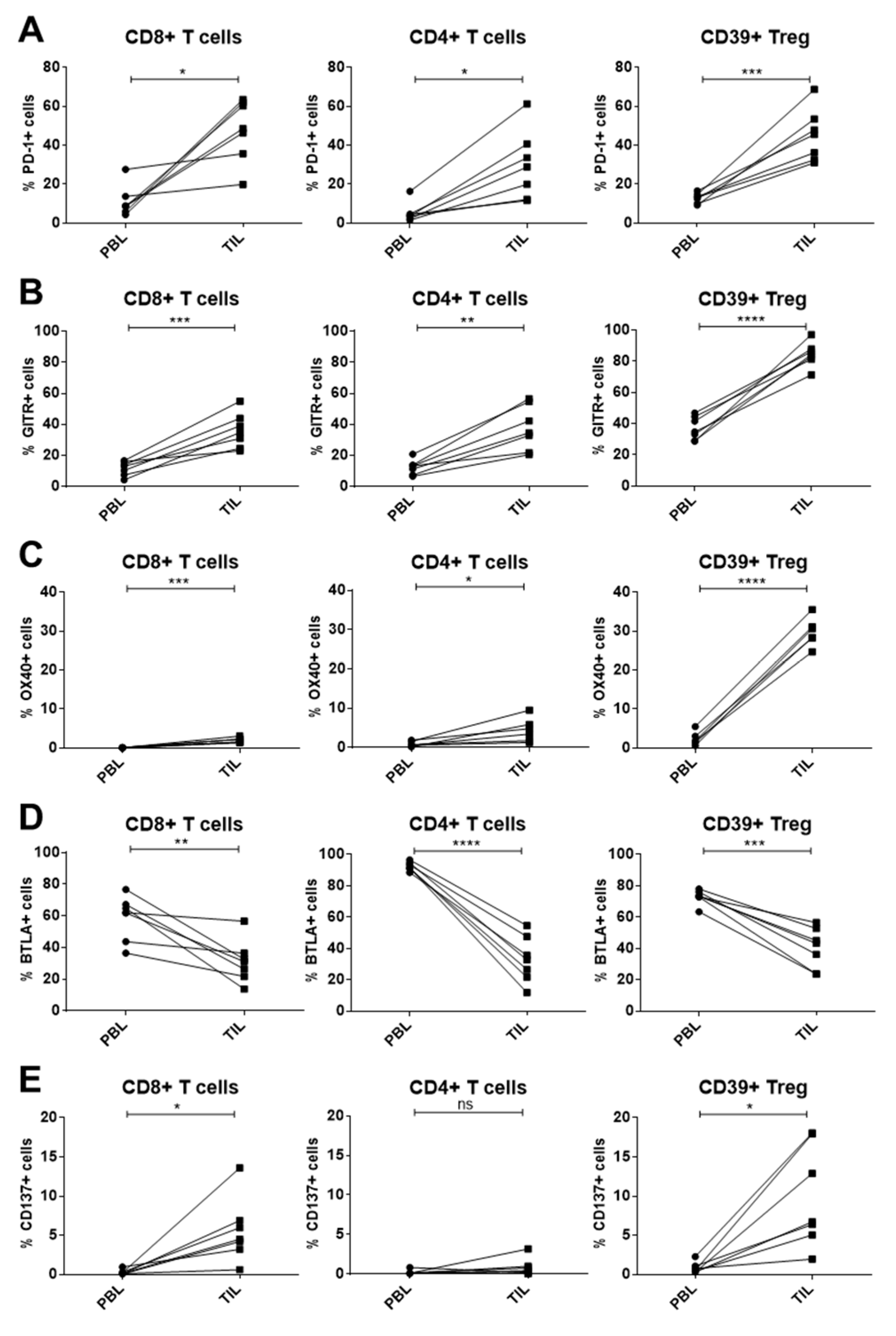
2.4. Immune Checkpoint Expression on T-Cell Subsets of PBL and Tumor-Infiltrating Lymphocytes (TIL) in HNSCC Patients
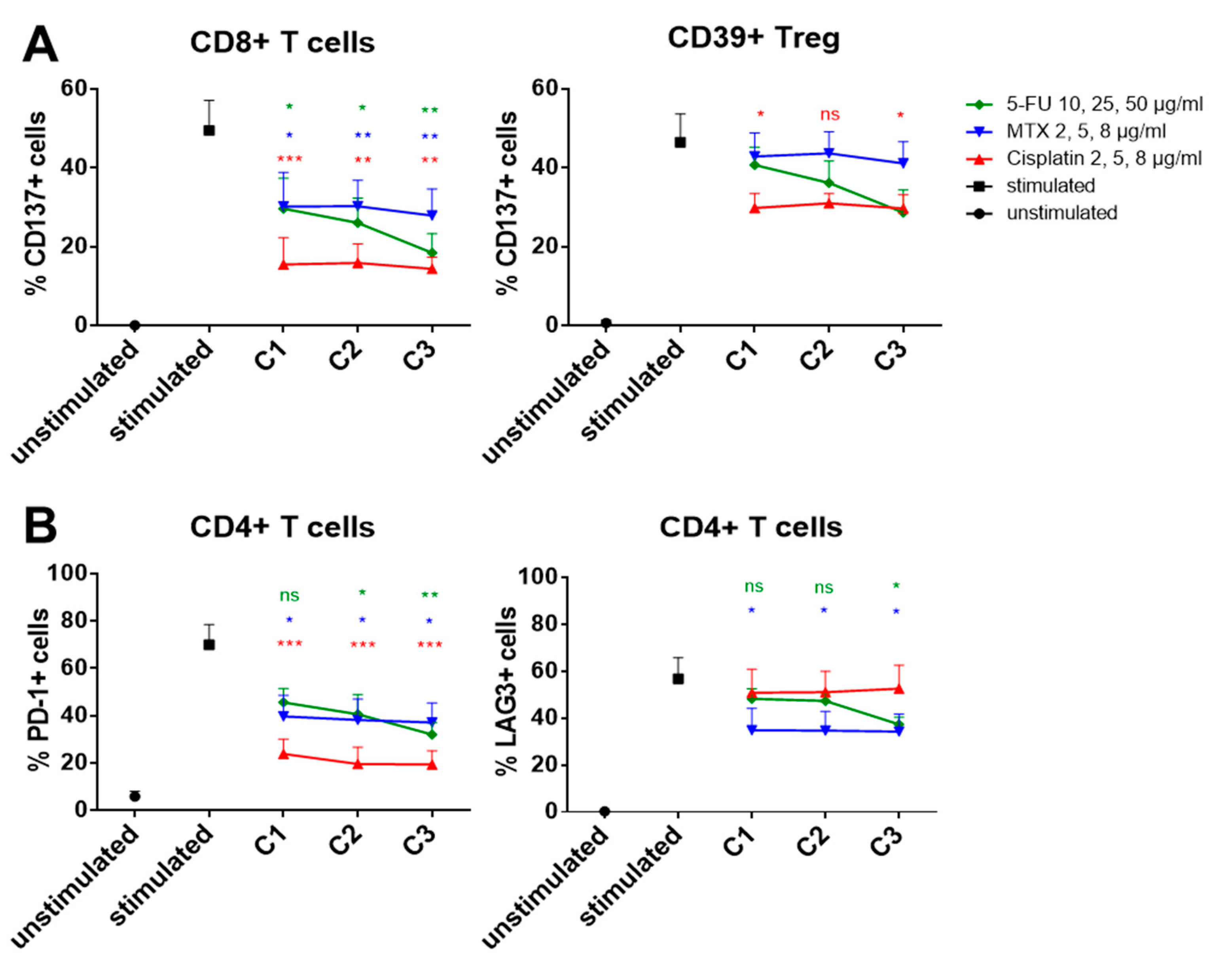
2.5. Influence of In Vitro Chemotherapy Treatment on Immune Checkpoint Expression
2.6. PD-1 Reduction on Immune Cells after In Vitro Nivolumab Treatment
3. Discussion
3.1. Altered Expression of Co-Inihbitory Molecules on PBL of HNSCC Patients
3.2. Dysregulation of Co-Stimulatory Molecules on PBL of HNSCC Patients
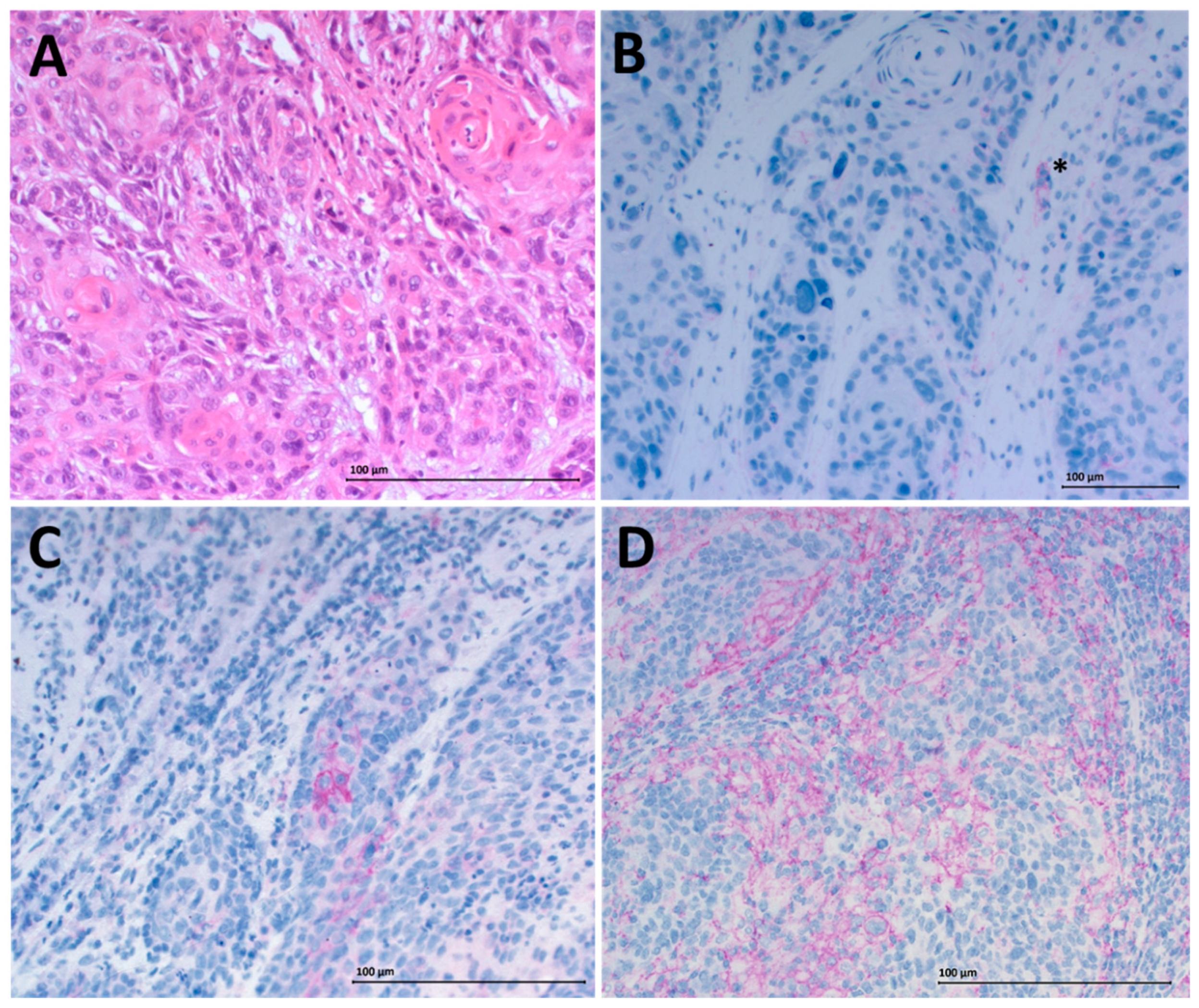
3.3. Modulated Immune Checkpoint Expression on Tumor-Infiltrating Immune Cells
3.4. Chemo-Immunomodulation of Circulating Immune Cell Subsets
3.5. PD-1 Reduction after In Vitro Checkpoint Blockade
4. Materials and Methods
4.1. Patient Samples and Specimens
4.2. Isolation of Peripheral Lymphocytes
4.3. Isolation of TIL
4.4. Stimulation of PBL In Vitro
4.5. Chemotherapeutic Treatment of PBL
4.6. Nivolumab Treatment of PBL
4.7. Antibodies and Reagents
4.8. Flow Cytometry
4.9. Gating Strategy
4.10. RT-PCR
4.11. IHC
4.12. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BTLA | B- and T-lymphocyte attenuator |
| CD137 | cluster of differentiation 137/tumor necrosis factor receptor superfamily member 9 (TNFRSF9) |
| CRT | chemoradiotherapy |
| DOAJ | Directory of open access journal |
| GITR | glucocorticoid-induced TNFR-related |
| HNSCC | head and neck squamous cell carcinoma |
| IL-2 | interleukin 2 |
| LAG3 | lymphocyte-activation gene 3 |
| mAb | monoclonal antibody |
| MDPI | Multidisciplinary Digital Publishing Institute |
| MTX | methotrexate |
| OX40 | tumor necrosis factor receptor superfamily member 4 (TNFRSF4) |
| PBL | peripheral blood lymphocytes |
| PD-1 | programmed cell death protein 1 |
| PD-L1 | programmed cell death-ligand 1 |
| SEB | Staphylococcus enterotoxin b |
| TIL | Tumor-infiltrating lymphocytes |
| TIM3 | t-cell immunoglobulin and mucin-domain containing-3 |
| Treg | regulatory T cells |
| TLA | Three letter acronym |
| 5-FU | 5-fluorouracil |
References
- Weinberg, A.D.; Morris, N.P.; Kovacsovics-Bankowski, M.; Urba, W.J.; Curti, B.D. Science gone translational: The OX40 agonist story. Immunol. Rev. 2011, 244, 218–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumeister, S.H.; Freeman, G.J.; Dranoff, G.; Sharpe, A.H. Coinhibitory Pathways in Immunotherapy for Cancer. Annu. Rev. Immunol. 2016, 34, 539–573. [Google Scholar] [CrossRef] [PubMed]
- Jie, H.B.; Gildener-Leapman, N.; Li, J.; Srivastava, R.M.; Gibson, S.P.; Whiteside, T.L.; Ferris, R.L. Intratumoral regulatory T cells upregulate immunosuppressive molecules in head and neck cancer patients. Br. J. Cancer 2013, 109, 2629–2635. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, A.G. Immune checkpoint blockade immunotherapy to activate anti-tumour T-cell immunity. Br. J. Haematol. 2013, 162, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Kuss, I.; Hathaway, B.; Ferris, R.L.; Gooding, W.; Whiteside, T.L. Decreased absolute counts of T lymphocyte subsets and their relation to disease in squamous cell carcinoma of the head and neck. Clin. Cancer Res. 2004, 10, 3755–3762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauernhofer, T.; Kuss, I.; Henderson, B.; Baum, A.S.; Whiteside, T.L. Preferential apoptosis of CD56dim natural killer cell subset in patients with cancer. Eur. J. Immunol. 2003, 33, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, C.; Kim, G.G.; Albers, A.; Hoermann, K.; Myers, E.N.; Whiteside, T.L. Characteristics of CD4+CD25+ regulatory T cells in the peripheral circulation of patients with head and neck cancer. Br. J. Cancer 2005, 92, 913–920. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Pignon, J.P.; le Maître, A.; Maillard, E.; Bourhis, J. Meta-analysis of chemotherapy in head and neck cancer (MACH-NC): An update on 93 randomised trials and 17,346 patients. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2009, 92, 4–14. [Google Scholar] [CrossRef]
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tân, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Hisamatsu, K.; Suzui, N.; Hara, A.; Tomita, H.; Miyazaki, T. A Review of HPV-Related Head and Neck Cancer. J. Clin. Med. 2018, 7, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, J.; Domenge, C.; Ozsahin, M.; Matuszewska, K.; Lefèbvre, J.L.; Greiner, R.H.; Giralt, J.; Maingon, P.; Rolland, F.; Bolla, M.; et al. Postoperative irradiation with or without concomitant chemotherapy for locally advanced head and neck cancer. N. Engl. J. Med. 2004, 350, 1945–1952. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.S.; Pajak, T.F.; Forastiere, A.A.; Jacobs, J.; Campbell, B.H.; Saxman, S.B.; Kish, J.A.; Kim, H.E.; Cmelak, A.J.; Rotman, M.; et al. Postoperative concurrent radiotherapy and chemotherapy for high-risk squamous-cell carcinoma of the head and neck. N. Engl. J. Med. 2004, 350, 1937–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siano, M.; Infante, G.; Resteghini, C.; Cau, M.C.; Alfieri, S.; Bergamini, C.; Granata, R.; Miceli, R.; Locati, L.; Licitra, L.; et al. Outcome of recurrent and metastatic head and neck squamous cell cancer patients after first line platinum and cetuximab therapy. Oral Oncol. 2017, 69, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.; Braun, M.; Dietrich, D.; Aktekin, S.; Hoft, S.; Kristiansen, G.; Goke, F.; Schrock, A.; Bragelmann, J.; Held, S.A.E.; et al. PD-L1: A novel prognostic biomarker in head and neck squamous cell carcinoma. Oncotarget 2017, 8, 52889–52900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burtness, B.; Harrington, K.J.; Greil, R.; Soulieres, D.; Tahara, M.; de Castro, G., Jr.; Psyrri, A.; Baste, N.; Neupane, P.; Bratland, A.; et al. Pembrolizumab alone or with chemotherapy versus cetuximab with chemotherapy for recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-048): A randomised, open-label, phase 3 study. Lancet 2019, 394, 1915–1928. [Google Scholar] [CrossRef]
- Davis, R.J.; Ferris, R.L.; Schmitt, N.C. Costimulatory and coinhibitory immune checkpoint receptors in head and neck cancer: Unleashing immune responses through therapeutic combinations. Cancers Head Neck 2016, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Detre, S.; Saclani Jotti, G.; Dowsett, M. A “quickscore” method for immunohistochemical semiquantitation: Validation for oestrogen receptor in breast carcinomas. J. Clin. Pathol. 1995, 48, 876–878. [Google Scholar] [CrossRef] [Green Version]
- Gros, A.; Robbins, P.F.; Yao, X.; Li, Y.F.; Turcotte, S.; Tran, E.; Wunderlich, J.R.; Mixon, A.; Farid, S.; Dudley, M.E.; et al. PD-1 identifies the patient-specific CD8+ tumor-reactive repertoire infiltrating human tumors. J. Clin. Investig. 2014, 124, 2246–2259. [Google Scholar] [CrossRef]
- Kim, H.R.; Park, H.J.; Son, J.; Lee, J.G.; Chung, K.Y.; Cho, N.H.; Shim, H.S.; Park, S.; Kim, G.; In Yoon, H.; et al. Tumor microenvironment dictates regulatory T cell phenotype: Upregulated immune checkpoints reinforce suppressive function. J. Immunother. Cancer 2019, 7, 339. [Google Scholar] [CrossRef]
- Ko, K.; Yamazaki, S.; Nakamura, K.; Nishioka, T.; Hirota, K.; Yamaguchi, T.; Shimizu, J.; Nomura, T.; Chiba, T.; Sakaguchi, S. Treatment of advanced tumors with agonistic anti-GITR mAb and its effects on tumor-infiltrating Foxp3+CD25+CD4+ regulatory T cells. J. Exp. Med. 2005, 202, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Shevach, E.M.; Stephens, G.L. The GITR-GITRL interaction: Co-stimulation or contrasuppression of regulatory activity? Nat. Rev. Immunol. 2006, 6, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, K.M.; Rennert, P.D.; Freeman, G.J. Combination cancer immunotherapy and new immunomodulatory targets. Nat. Rev. Drug Discov. 2015, 14, 561–584. [Google Scholar] [CrossRef]
- Buchan, S.L.; Dou, L.; Remer, M.; Booth, S.G.; Dunn, S.N.; Lai, C.; Semmrich, M.; Teige, I.; Martensson, L.; Penfold, C.A.; et al. Antibodies to Costimulatory Receptor 4-1BB Enhance Anti-tumor Immunity via T Regulatory Cell Depletion and Promotion of CD8 T Cell Effector Function. Immunity 2018, 49, 958–970.e957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zappasodi, R.; Sirard, C.; Li, Y.; Budhu, S.; Abu-Akeel, M.; Liu, C.; Yang, X.; Zhong, H.; Newman, W.; Qi, J.; et al. Rational design of anti-GITR-based combination immunotherapy. Nat. Med. 2019, 25, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.B.; Duhen, R.; Leidner, R.S.; Curti, B.D.; Ballesteros-Merino, C.; Piening, B.; Bernard, B.; Pucilowska, J.; Bifulco, C.B.; Fox, B.A.; et al. Neoadjuvant anti-OX40 (MEDI6469) prior to surgery in head and neck squamous cell carcinoma. J. Clin. Oncol. 2018, 36, 6011. [Google Scholar] [CrossRef]
- Murphy, K.M.; Nelson, C.A.; Sedy, J.R. Balancing co-stimulation and inhibition with BTLA and HVEM. Nat. Rev. Immunol. 2006, 6, 671–681. [Google Scholar] [CrossRef]
- Haymaker, C.; Wu, R.; Bernatchez, C.; Radvanyi, L. PD-1 and BTLA and CD8(+) T-cell “exhaustion” in cancer: “Exercising” an alternative viewpoint. Oncoimmunology 2012, 1, 735–738. [Google Scholar] [CrossRef] [Green Version]
- Schuler, P.J.; Harasymczuk, M.; Schilling, B.; Saze, Z.; Strauss, L.; Lang, S.; Johnson, J.T.; Whiteside, T.L. Effects of adjuvant chemoradiotherapy on the frequency and function of regulatory T cells in patients with head and neck cancer. Clin. Cancer Res. 2013, 19, 6585–6596. [Google Scholar] [CrossRef] [Green Version]
- Schuler, P.J.; Trellakis, S.; Greve, J.; Bas, M.; Bergmann, C.; Bolke, E.; Lehnerdt, G.; Mattheis, S.; Albers, A.E.; Brandau, S.; et al. In vitro chemosensitivity of head and neck cancer cell lines. Eur. J. Med Res. 2010, 15, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Caraglia, M.; Correale, P.; Giannicola, R.; Staropoli, N.; Botta, C.; Pastina, P.; Nesci, A.; Caporlingua, N.; Francini, E.; Ridolfi, L.; et al. GOLFIG Chemo-Immunotherapy in Metastatic Colorectal Cancer Patients. A Critical Review on a Long-Lasting Follow-Up. Front. Oncol. 2019, 9, 1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correale, P.; Botta, C.; Ciliberto, D.; Pastina, P.; Ingargiola, R.; Zappavigna, S.; Tassone, P.; Pirtoli, L.; Caraglia, M.; Tagliaferri, P. Immunotherapy of colorectal cancer: New perspectives after a long path. Immunotherapy 2016, 8, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Pastina, P.; Nardone, V.; Croci, S.; Battaglia, G.; Vanni, F.; Bellan, C.; Barbarino, M.; Ricci, V.; Costantini, S.; Capone, F.; et al. Anti-cancer activity of dose-fractioned mPE +/- bevacizumab regimen is paralleled by immune-modulation in advanced squamous NSLC patients. J. Thorac. Dis. 2017, 9, 3123–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correale, P.; Saladino, R.E.; Giannarelli, D.; Giannicola, R.; Agostino, R.; Staropoli, N.; Strangio, A.; Del Giudice, T.; Nardone, V.; Altomonte, M.; et al. Distinctive germline expression of class I human leukocyte antigen (HLA) alleles and DRB1 heterozygosis predict the outcome of patients with non-small cell lung cancer receiving PD-1/PD-L1 immune checkpoint blockade. J. Immunother. Cancer 2020, 8, e000733. [Google Scholar] [CrossRef] [PubMed]
- Correale, P.; Saladino, R.E.; Nardone, V.; Giannicola, R.; Agostino, R.; Pirtoli, L.; Caraglia, M. Could PD-1/PDL1 immune checkpoints be linked to HLA signature? Immunotherapy 2019, 11, 1523–1526. [Google Scholar] [CrossRef] [PubMed]
- Schuler, P.J.; Harasymczuk, M.; Schilling, B.; Lang, S.; Whiteside, T.L. Separation of human CD4+CD39+ T cells by magnetic beads reveals two phenotypically and functionally different subsets. J. Immunol. Methods 2011, 369, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeske, S.S.; Weissinger, S.E.; Veit, J.A.; Brunner, C.; Huber, U.; Theodoraki, M.N.; Hoffmann, T.K.; Schuler, P.J.; Doescher, J. Treatment-induced changes of lymphocyte subsets in patients with adenoid cystic carcinoma of the head and neck. Eur. Arch. Oto-Rhino-Laryngol. 2019, 276, 1465–1473. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group | HNSCC Patients (Blood) | HNSCC Patients (Tumor Tissue) | |
|---|---|---|---|
| n (female/male) | 23 (13/10) | 23 (9/14) | 12 (5/7) |
| Age (±SD) range (y) | 56 ± 19 (27–84) | 59 ± 11 (37–74) | 67 ± 9 (49–77) |
| Stage (n) | |||
| T1 | 4 | 2 | |
| T2 | 8 | 6 | |
| T3 | 5 | 2 | |
| T4 | 6 | 2 | |
| Nodal status (n) | |||
| N0 | 7 | 2 | |
| N1 | 5 | 3 | |
| N2 | 7 | 7 | |
| N3 | 4 | 0 | |
| Location (n) | |||
| Oral cavity | 5 | 1 | |
| Oropharynx | 10 | 8 | |
| Hypopharynx | 4 | 1 | |
| Larynx | 4 | 2 | |
| PD-L1 status on tumor tissue (n) | |||
| 0% | 3 | ||
| ≥1% | 5 | ||
| ≥5% | 4 | ||
| PD-L1 Quickscore | |||
| 0 | 3 | ||
| 1 | 3 | ||
| 2 | 2 | ||
| 4 | 2 | ||
| 6 | 2 | ||
| HPV status | |||
| Positive | 7 | ||
| Negative | 16 | ||
| Overall survival at 3 y | 22/23 | ||
| Disease-free survival at 3 y | 21/23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puntigam, L.K.; Jeske, S.S.; Götz, M.; Greiner, J.; Laban, S.; Theodoraki, M.-N.; Doescher, J.; Weissinger, S.E.; Brunner, C.; Hoffmann, T.K.; et al. Immune Checkpoint Expression on Immune Cells of HNSCC Patients and Modulation by Chemo- and Immunotherapy. Int. J. Mol. Sci. 2020, 21, 5181. https://doi.org/10.3390/ijms21155181
Puntigam LK, Jeske SS, Götz M, Greiner J, Laban S, Theodoraki M-N, Doescher J, Weissinger SE, Brunner C, Hoffmann TK, et al. Immune Checkpoint Expression on Immune Cells of HNSCC Patients and Modulation by Chemo- and Immunotherapy. International Journal of Molecular Sciences. 2020; 21(15):5181. https://doi.org/10.3390/ijms21155181
Chicago/Turabian StylePuntigam, Lisa K., Sandra S. Jeske, Marlies Götz, Jochen Greiner, Simon Laban, Marie-Nicole Theodoraki, Johannes Doescher, Stephanie E. Weissinger, Cornelia Brunner, Thomas K. Hoffmann, and et al. 2020. "Immune Checkpoint Expression on Immune Cells of HNSCC Patients and Modulation by Chemo- and Immunotherapy" International Journal of Molecular Sciences 21, no. 15: 5181. https://doi.org/10.3390/ijms21155181