Chemical Mechanisms of Nanoparticle Radiosensitization and Radioprotection: A Review of Structure-Function Relationships Influencing Reactive Oxygen Species


Abstract
:1. Introduction
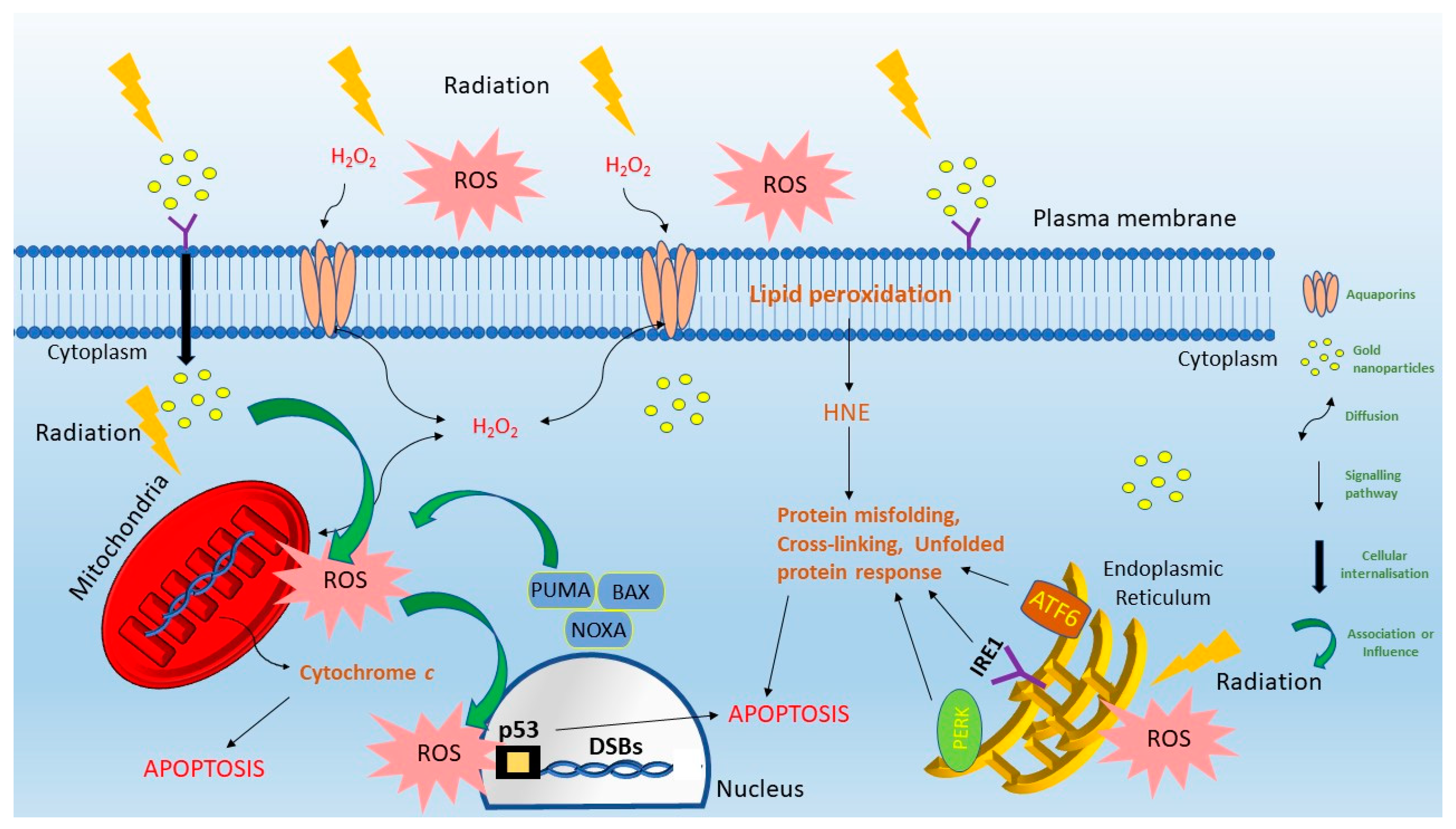
2. Nanoparticle Localization, ROS Transport and Cellular Damage
3. Mechanisms of Nanoparticle ROS Enhancement
4. Types of ROS and Analysis Methods
4.1. Raman Spectroscopy
4.2. Fluorescent Dyes
4.3. Electron Spin Resonance
5. Dependence on Metal Content
6. Dependence on Size
7. Dependence on Shape, Structure and Stability
8. Dependence on Surface Functionalization
9. Summary
Funding
Conflicts of Interest
References
- Kunz-Schughart, L.A.; Dubrovska, A.; Peitzsch, C.; Ewe, A.; Aigner, A.; Schellenburg, S.; Muders, M.H.; Hampel, S.; Cirillo, G.; Iemma, F.; et al. Nanoparticles for radiooncology: Mission, vision, challenges. Biomaterials 2017, 120, 155–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Tse, K.; Zahedi, P.; Harding, S.M.; Zafarana, G.; Jaffray, D.A.; Bristow, R.G.; Allen, C. Hypoxia and Cellular Localization Influence the Radiosensitizing Effect of Gold Nanoparticles (AuNPs) in Breast Cancer Cells. Radiat. Res. 2014, 182, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Jeynes, J.C.; Merchant, M.J.; Spindler, A.; Wera, A.C.; Kirkby, K.J. Investigation of gold nanoparticle radiosensitization mechanisms using a free radical scavenger and protons of different energies. Phys. Med. Biol. 2014, 59, 6431–6443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Lou, J.; Hristov, D. X-ray responsive nanoparticles with triggered release of nitrite, a precursor of reactive nitrogen species, for enhanced cancer radiosensitization. Nanoscale 2017, 9, 14627–14634. [Google Scholar] [CrossRef] [PubMed]
- Morrison, R.A.; Rybak-Smith, M.J.; Thompson, J.M.; Thiebaut, B.; Hill, M.A.; Townley, H.E. Efficacy of radiosensitizing doped titania nanoparticles under hypoxia and preparation of an embolic microparticle. Int. J. Nanomed. 2017, 12, 3851–3863. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhou, M.; Li, C. Synthetic nanoparticles for delivery of radioisotopes and radiosensitizers in cancer therapy. Cancer Nano 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Vilotte, F.; Jumeau, R.; Bourhis, J. High Z nanoparticles and radiotherapy: A critical view. Lancet Oncol. 2019, 20, e557. [Google Scholar] [CrossRef] [Green Version]
- Bonvalot, S.; le Pechoux, C.; de Baere, T.; Kantor, G.; Buy, X.; Stoeckle, E.; Terrier, P.; Sargos, P.; Coindre, J.M.; Lassau, N.; et al. First-in-Human Study Testing a New Radioenhancer Using Nanoparticles (NBTXR3) Activated by Radiation Therapy in Patients with Locally Advanced Soft Tissue Sarcomas. Clin. Cancer Res. 2017, 23, 908–917. [Google Scholar] [CrossRef] [Green Version]
- Verry, C.; Sancey, L.; Dufort, S.; le Duc, G.; Mendoza, C.; Lux, F.; Grand, S.; Arnaud, J.; Quesada, J.L.; Villa, J.; et al. Treatment of multiple brain metastases using gadolinium nanoparticles and radiotherapy: NANO-RAD, a phase I study protocol. BMJ Open 2019, 9, e023591. [Google Scholar] [CrossRef]
- Bonvalot, S.; Rutkowski, P.L.; Thariat, J.; Carrère, S.; Ducassou, A.; Sunyach, M.-P.; Agoston, P.; Hong, A.; Mervoyer, A.; Rastrelli, M.; et al. NBTXR3, a first-in-class radioenhancer hafnium oxide nanoparticle, plus radiotherapy versus radiotherapy alone in patients with locally advanced soft-tissue sarcoma (Act.In.Sarc): A multicentre, phase 2–3, randomised, controlled trial. Lancet Oncol. 2019, 20, 1148–1159. [Google Scholar] [CrossRef]
- Nanobiotix. Nanoxray Technology. Available online: http://www.nanobiotix.com/_en/nanoxray/ (accessed on 14 January 2020).
- McMahon, S.J.; Hyland, W.B.; Muir, M.F.; Coulter, J.A.; Jain, S.; Butterworth, K.T.; Schettino, G.; Dickson, G.R.; Hounsell, A.R.; O’Sullivan, J.M.; et al. Biological consequences of nanoscale energy deposition near irradiated heavy atom nanoparticles. Sci. Rep. 2011, 1, 18. [Google Scholar] [CrossRef] [Green Version]
- Rezaee, Z.; Yadollahpour, A.; Bayati, V.; Negad Dehbashi, F. Gold nanoparticles and electroporation impose both separate and synergistic radiosensitizing effects in HT-29 tumor cells: An in vitro study. Int. J. Nanomed. 2017, 12, 1431–1439. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Xue, P.; Liu, S.; Zhang, L.; Guan, Q.; Zhu, J.; Tian, X. Metal-Based Hybrid Nanoparticles as Radiosensitizers in Cancer Therapy. Colloid Interface Sci. Commun. 2018, 23, 45–51. [Google Scholar] [CrossRef]
- Stewart, C.; Konstantinov, K.; McKinnon, S.; Guatelli, S.; Lerch, M.; Rosenfeld, A.; Tehei, M.; Corde, S. First proof of bismuth oxide nanoparticles as efficient radiosensitisers on highly radioresistant cancer cells. Phys. Med. 2016, 32, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Grall, R.; Girard, H.; Saad, L.; Petit, T.; Gesset, C.; Combis-Schlumberger, M.; Paget, V.; Delic, J.; Arnault, J.-C.; Chevillard, S. Impairing the radioresistance of cancer cells by hydrogenated nanodiamonds. Biomaterials 2015, 61, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Peukert, D.; Kempson, I.; Douglass, M.; Bezak, E. Metallic nanoparticle radiosensitisation of ion radiotherapy: A review. Phys. Med. 2018, 47, 121–128. [Google Scholar] [CrossRef]
- Asharani, P.; Xinyi, N.; Hande, M.P.; Valiyaveettil, S. DNA damage and p53-mediated growth arrest in human cells treated with platinum nanoparticles. Nanomedicine 2010, 5, 51–64. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, P.; Li, F.; Jin, X.; Li, J.; Chen, W.; Li, Q. Metal-based NanoEnhancers for Future Radiotherapy: Radiosensitizing and Synergistic Effects on Tumor Cells. Thernostics 2018, 8, 1824–1849. [Google Scholar] [CrossRef]
- Kang, B.; Mackey, M.A.; El-Sayed, M.A. Nuclear targeting of gold nanoparticles in cancer cells induces DNA damage, causing cytokinesis arrest and apoptosis. J. Am. Chem. Soc. 2010, 132. [Google Scholar] [CrossRef]
- Janic, B.; Liu, F.; Bobbitt, K.; Brown, S.L.; Chetty, I.; Mao, G.; Movsas, B.; Wen, N. Cellular Uptake and Radio-sensitization Effect of Small Gold Nanoparticles in MCF-7 Breast Cancer Cells. J. Nanomed. Nanotechnol. 2018, 9. [Google Scholar] [CrossRef]
- Kong, T.; Zeng, J.; Wang, X.; Yang, X.; Yang, J.; McQuarrie, S. Enhancement of radiation cytotoxicity in breast-cancer cells by localized attachment of gold nanoparticles. Small 2008, 4. [Google Scholar] [CrossRef]
- Jain, S.; Coulter, J.A.; Hounsell, A.R.; Butterworth, K.T.; McMahon, S.J.; Hyland, W.B. Cell-specific radiosensitization by gold nanoparticles at megavoltage radiation energies. Int. J. Radiat. Oncol. Biol. Phys. 2011, 79. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, T.; Douglass, M.; Williamson, N.H.; Howard, D.; Bhardwaj, R.; Lawrence, M.; Paterson, D.J.; Bezak, E.; Thierry, B.; Kempson, I.M. Cross-Correlative Single-Cell Analysis Reveals Biological Mechanisms of Nanoparticle Radiosensitization. ACS Nano 2019, 13, 5077–5090. [Google Scholar] [CrossRef]
- Pagáčová, E.; Štefančíková, L.; Schmidt-Kaler, F.; Hildenbrand, G.; Vičar, T.; Depeš, D.; Lee, J.-H.; Bestvater, F.; Lacombe, S.; Porcel, E.; et al. Challenges and Contradictions of Metal Nano-Particle Applications for Radio-Sensitivity Enhancement in Cancer Therapy. Int. J. Mol. Sci. 2019, 20, 588. [Google Scholar] [CrossRef] [Green Version]
- Forkink, M.; Smeitink, J.A.M.; Brock, R.; Willems, P.H.G.M.; Koopman, W.J.H. Detection and manipulation of mitochondrial reactive oxygen species in mammalian cells. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 1034–1044. [Google Scholar] [CrossRef] [Green Version]
- Manke, A.; Wang, L.; Rojanasakul, Y. Mechanisms of Nanoparticle-Induced Oxidative Stress and Toxicity. BioMed Res. Int. 2013, 2013, 942916. [Google Scholar] [CrossRef] [Green Version]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Borrego-Soto, G.; Ortiz-López, R.; Rojas-Martínez, A. Ionizing radiation-induced DNA injury and damage detection in patients with breast cancer. Genet. Mol. Biol. 2015, 38, 420–432. [Google Scholar] [CrossRef]
- Liou, G.-Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Lee, S.; Seo, D.; Kim, D.; Kim, K.; Kim, E.; Kang, J.; Seong, K.M.; Youn, H.; Youn, B. Cellular Stress Responses in Radiotherapy. Cells 2019, 8, 1105. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxidative Med. Cell. Longev. 2016, 2016, 18. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.H.; Hanagata, N.; Ikoma, T.; Huang, J.Y.; Li, K.Y.; Lin, C.P.; Lin, F.H. Hafnium-doped hydroxyapatite nanoparticles with ionizing radiation for lung cancer treatment. Acta Biomater. 2016, 37, 165–173. [Google Scholar] [CrossRef]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Yang, J.; Xu, Z.-P.; Huang, Y.; Hamrick, H.E.; Duerksen-Hughes, P.J.; Yu, Y.-N. ATM and ATR: Sensing DNA damage. World J. Gastroenterol. 2004, 10, 155–160. [Google Scholar] [CrossRef]
- Bartek, J.; Lukas, J. Chk1 and Chk2 kinases in checkpoint control and cancer. Cancer Cell 2003, 3, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Menon, V.; Povirk, L. Involvement of p53 in the repair of DNA double strand breaks: Multifaceted Roles of p53 in homologous recombination repair (HRR) and non-homologous end joining (NHEJ). Subcell Biochem. 2014, 85, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Howard, S.M.; Yanez, D.A.; Stark, J.M. DNA Damage Response Factors from Diverse Pathways, Including DNA Crosslink Repair, Mediate Alternative End Joining. PLoS Genet. 2015, 11, e1004943. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.K.; Kumar, S.; Choi, E.-H.; Sharma, P.; Misra, S.; Kim, M.-H. Insight Into the Molecular Dynamic Simulation Studies of Reactive Oxygen Species in Native Skin Membrane. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Schaur, R.J.; Siems, W.; Bresgen, N.; Eckl, P.M. 4-Hydroxy-nonenal-A Bioactive Lipid Peroxidation Product. Biomolecules 2015, 5, 2247–2337. [Google Scholar] [CrossRef] [Green Version]
- Siwecka, N.; Rozpędek, W.; Pytel, D.; Wawrzynkiewicz, A.; Dziki, A.; Dziki, Ł.; Diehl, J.A.; Majsterek, I. Dual role of Endoplasmic Reticulum Stress-Mediated Unfolded Protein Response Signaling Pathway in Carcinogenesis. Int. J. Mol. Sci. 2019, 20, 4354. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.-H.; Yen, J.-H.; Weng, C.-Y.; Wang, L.; Ha, C.-L.; Wu, M.-J. Lipid peroxidation end product 4-hydroxy-trans-2-nonenal triggers unfolded protein response and heme oxygenase-1 expression in PC12 cells: Roles of ROS and MAPK pathways. Toxicology 2014, 315, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Kolesnick, R. The therapeutic potential of modulating the ceramide/sphingomyelin pathway. J. Clin. Investig. 2002, 110, 3–8. [Google Scholar] [CrossRef]
- Liao, W.-C.; Haimovitz-Friedman, A.; Persaud, R.S.; McLoughlin, M.; Ehleiter, D.; Zhang, N.; Gatei, M.; Lavin, M.; Kolesnick, R.; Fuks, Z. Ataxia Telangiectasia-mutated Gene Product Inhibits DNA Damage-induced Apoptosis via Ceramide Synthase. J. Biol. Chem. 1999, 274, 17908–17917. [Google Scholar] [CrossRef] [Green Version]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Tamma, G.; Valenti, G.; Grossini, E.; Donnini, S.; Marino, A.; Marinelli, R.A.; Calamita, G. Aquaporin membrane channels in oxidative stress, cell signaling, and aging: Recent advances and research trends. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Kodiha, M.; Stochaj, U. Nuclear Transport: A Switch for the Oxidative Stress—Signaling Circuit? J. Signal Transduct. 2012, 2012, 208650. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Gabaldón, T.; Pittis, A.A. Origin and evolution of metabolic sub-cellular compartmentalization in eukaryotes. Biochimie 2015, 119, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Shinoda, W. Permeability across lipid membranes. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 2254–2265. [Google Scholar] [CrossRef]
- Bienert, G.P.; Schjoerring, J.K.; Jahn, T.P. Membrane transport of hydrogen peroxide. Biochim. Biophys. Acta (BBA) Biomembr. 2006, 1758, 994–1003. [Google Scholar] [CrossRef] [Green Version]
- Madeira, A.; Moura, T.F.; Soveral, G. Detecting Aquaporin Function and Regulation. Front. Chem. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Bienert, G.P.; Møller, A.L.B.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific Aquaporins Facilitate the Diffusion of Hydrogen Peroxide across Membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Henzler, T.; Steudle, E. Transport and metabolic degradation of hydrogen peroxide in Chara corallina: Model calculations and measurements with the pressure probe suggest transport of H2O2 across water channels. J. Exp. Bot. 2000, 51, 2053–2066. [Google Scholar] [CrossRef] [Green Version]
- Provost, C.; Choufani, F.; Avedanian, L.; Bkaily, G.; Gobeil, F.; Jacques, D. Nitric oxide and reactive oxygen species in the nucleus revisited. Can. J. Physiol. Pharmacol. 2010, 88, 296–304. [Google Scholar] [CrossRef]
- Chittiboyina, S.; Bai, Y.; Lelièvre, S.A. Microenvironment-Cell Nucleus Relationship in the Context of Oxidative Stress. Front. Cell Dev. Biol. 2018, 6, 23. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, Z.; Min, W. Mitochondria, Oxidative Stress and Innate Immunity. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Babu, K.R.; Tay, Y. The Yin-Yang Regulation of Reactive Oxygen Species and MicroRNAs in Cancer. Int. J. Mol. Sci. 2019, 20, 5335. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Xu, Y. p53, oxidative stress, and aging. Antioxid. Redox Signal. 2011, 15, 1669–1678. [Google Scholar] [CrossRef]
- Shi, Y.; Nikulenkov, F.; Zawacka-Pankau, J.; Li, H.; Gabdoulline, R.; Xu, J.; Eriksson, S.; Hedström, E.; Issaeva, N.; Kel, A.; et al. ROS-dependent activation of JNK converts p53 into an efficient inhibitor of oncogenes leading to robust apoptosis. Cell Death Differ. 2014, 21, 612–623. [Google Scholar] [CrossRef] [Green Version]
- Faraonio, R.; Vergara, P.; Di Marzo, D.; Pierantoni, M.G.; Napolitano, M.; Russo, T.; Cimino, F. p53 Suppresses the Nrf2-dependent Transcription of Antioxidant Response Genes. J. Biol. Chem. 2006, 281, 39776–39784. [Google Scholar] [CrossRef] [Green Version]
- Macip, S.; Igarashi, M.; Berggren, P.; Yu, J.; Lee, S.W.; Aaronson, S.A. Influence of induced reactive oxygen species in p53-mediated cell fate decisions. Mol. Cell Biol. 2003, 23, 8576–8585. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Qi, F.; Kobayashi, J. Potential relationship between the biological effects of low-dose irradiation and mitochondrial ROS production. J. Radiat. Res. 2018, 59, ii91–ii97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shawgo, M.E.; Shelton, S.N.; Robertson, J.D. Caspase-mediated Bak activation and cytochrome c release during intrinsic apoptotic cell death in Jurkat cells. J. Biol. Chem. 2008, 283, 35532–35538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, X.; Wang, Y.; Ma, X.; Li, Y.; Zhang, Z.; Xiao, Z.; Liu, L.; Gao, X.; Liu, J. Mitochondria-targeting Au nanoclusters enhance radiosensitivity of cancer cells. J. Mater. Chem. B 2017, 5, 4190–4197. [Google Scholar] [CrossRef]
- Zeeshan, H.M.A.; Lee, G.H.; Kim, H.-R.; Chae, H.-J. Endoplasmic Reticulum Stress and Associated ROS. Int. J. Mol. Sci. 2016, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [Green Version]
- Haeri, M.; Knox, B.E. Endoplasmic Reticulum Stress and Unfolded Protein Response Pathways: Potential for Treating Age-related Retinal Degeneration. J. Ophthalmic Vis. Res. 2012, 7, 45–59. [Google Scholar]
- Márton, M.; Kurucz, A.; Lizák, B.; Margittai, É.; Bánhegyi, G.; Kapuy, O. A Systems Biological View of Life-and-Death Decision with Respect to Endoplasmic Reticulum Stress-The Role of PERK Pathway. Int. J. Mol. Sci. 2017, 18, 58. [Google Scholar] [CrossRef] [Green Version]
- Suh, D.H.; Kim, M.-K.; Kim, H.S.; Chung, H.H.; Song, Y.S. Unfolded protein response to autophagy as a promising druggable target for anticancer therapy. Ann. N. Y. Acad. Sci. 2012, 1271, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Butterworth, K.T.; McMahon, S.J.; Currell, F.J.; Prise, K.M. Physical basis and biological mechanisms of gold nanoparticle radiosensitization. Nanoscale 2012, 4, 4830–4838. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, L.; Fu, J.; Shi, C.; Xu, J.; Zhu, Y. Efficient free radical generation against cancer cells by low-dose X-ray irradiation with a functional SPC delivery nanosystem. J. Mater. Chem. B 2016, 4, 5863–5872. [Google Scholar] [CrossRef]
- Liu, R. Adsorption and dissociation of H2O on Au (111) surface: A DFT study. Comput. Theor. Chem. 2013, 1019, 141–145. [Google Scholar] [CrossRef]
- Gilles, M.B.; Brun, E.; Sicard-Roselli, C. Quantification of hydroxyl radicals and solvated electrons produced by irradiated gold nanoparticles suggests a crucial role of interfacial water. J. Colloid Interface Sci. 2018, 525, 31–38. [Google Scholar] [CrossRef]
- Misawa, M.; Takahashi, J. Generation of reactive oxygen species induced by gold nanoparticles under X-ray and UV Irradiations. Nanomed. Nanotechnol. Biol. Med. 2011, 7. [Google Scholar] [CrossRef]
- Nikolaos, M.; Dimitriou, G.T.; Evangelos, C.; Balanikas, E.; Pavlopoulou, A.; Mitsiogianni, M.; Mantso, T.; Pashos, G.; Boudouvis, A.G.; Lykakis, I.N.; et al. Gold nanoparticles, radiations and the immune system: Current insights into the physical mechanisms and the biological interactions of this new alliance towards cancer therapy. Pharmacol. Ther. 2017, 1–17. [Google Scholar]
- Wu, H.; Yin, J.-J.; Wamer, W.G.; Zeng, M.; Lo, Y.M. Reactive oxygen species-related activities of nano-iron metal and nano-iron oxides. J. Food Drug Anal. 2014, 22, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.; Sommer, A.; Distel, L.V.R.; Hazemann, J.-L.; Kröner, W.; Neuhuber, W.; Müller, P.; Proux, O.; Kryschi, C. Superparamagnetic Iron Oxide Nanoparticles as Novel X-ray Enhancer for Low-Dose Radiation Therapy. J. Phys. Chem. B 2014, 118, 6159–6166. [Google Scholar] [CrossRef]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. The Role of Reactive Oxygen Species (ROS) in the Biological Activities of Metallic Nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [Green Version]
- Peukert, D.; Incerti, S.; Kempson, I.; Douglass, M.; Karamitros, M.; Baldacchino, G.; Bezak, E. Validation and investigation of reactive species yields of Geant4-DNA chemistry models. Med. Phys. 2019, 46, 983–998. [Google Scholar] [CrossRef]
- Caputo, F.; de Nicola, M.; Ghibelli, L. Pharmacological potential of bioactive engineered nanomaterials. Biochem. Pharmacol. 2014, 92, 112–130. [Google Scholar] [CrossRef]
- Wardman, P.; Candeias, L.P. Fenton Chemistry: An Introduction. Radiat. Res. 1996, 145, 523–531. [Google Scholar] [CrossRef]
- Goyal, M.M.; Basak, A. Hydroxyl radical generation theory: A possible explanation of unexplained actions of mammalian catalase. Int. J. Biochem. Mol. Biol. 2012, 3, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Khaing Oo, M.K.; Yang, Y.; Hu, Y.; Gomez, M.; Du, H.; Wang, H. Gold Nanoparticle-Enhanced and Size-Dependent Generation of Reactive Oxygen Species from Protoporphyrin IX. ACS Nano 2012, 6, 1939–1947. [Google Scholar] [CrossRef]
- Lord, M.S.; Jung, M.; Teoh, W.Y.; Gunawan, C.; Vassie, J.A.; Amal, R.; Whitelock, J.M. Cellular uptake and reactive oxygen species modulation of cerium oxide nanoparticles in human monocyte cell line U937. Biomaterials 2012, 33, 7915–7924. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Z.; Huang, Y.; Feng, S.; Zheng, Z.; Liu, X.; Liu, M. Label-free detection of hydrogen peroxide-induced oxidative stress in human retinal pigment epithelium cells via laser tweezers Raman spectroscopy. Biomed. Opt. Express 2019, 10, 500–513. [Google Scholar] [CrossRef]
- Panikkanvalappil, S.R.; Mahmoud, M.A.; Mackey, M.A.; El-Sayed, M.A. Surface-Enhanced Raman Spectroscopy for Real-Time Monitoring of Reactive Oxygen Species-Induced DNA Damage and Its Prevention by Platinum Nanoparticles. ACS Nano 2013, 7, 7524–7533. [Google Scholar] [CrossRef]
- Setsukinai, K.-I.; Urano, Y.; Kakinuma, K.; Majima, H.J.; Nagano, T. Development of Novel Fluorescence Probes That Can Reliably Detect Reactive Oxygen Species and Distinguish Specific Species. J. Biol. Chem. 2003, 278, 3170–3175. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.; Fernandes, E.; Lima, J.L.F.C. Fluorescence probes used for detection of reactive oxygen species. J. Biochem. Biophys. Methods 2005, 65, 45–80. [Google Scholar] [CrossRef]
- Hempel, S.L.; Buettner, G.R.; O’Malley, Y.Q.; Wessels, D.A.; Flaherty, D.M. Dihydrofluorescein diacetate is superior for detecting intracellular oxidants: Comparison with 2′,7′-dichlorodihydrofluorescein diacetate, 5(and 6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic. Biol. Med. 1999, 27, 146–159. [Google Scholar] [CrossRef]
- Kalyanaraman, B. Oxidative chemistry of fluorescent dyes: Implications in the detection of reactive oxygen and nitrogen species. Biochem. Soc. Trans. 2011, 39, 1221. [Google Scholar] [CrossRef] [Green Version]
- Gollmer, A.; Arnbjerg, J.; Blaikie, F.H.; Pedersen, B.W.; Breitenbach, T.; Daasbjerg, K.; Glasius, M.; Ogilby, P.R. Singlet Oxygen Sensor Green®: Photochemical Behavior in Solution and in a Mammalian Cell. Photochem. Photobiol. 2011, 87, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, X.H.; Hu, X.D.; Zhang, W.; Lou, Z.C.; Xie, L.H.; Liu, P.D.; Zhang, H.Q. Enhancement of radiotherapy by ceria nanoparticles modified with neogambogic acid in breast cancer cells. Int. J. Nanomed. 2015, 10, 4957–4969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhong, Z.; Xu, Z.; Chen, L.; Wang, Y. 2′,7′-Dichlorodihydrofluorescein as a fluorescent probe for reactive oxygen species measurement: Forty years of application and controversy. Free Radic. Res. 2010, 44, 587–604. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, T.; Mizutani, A.; Murakami, A.; Kojo, S.; Ishii, T.; Taketani, S. Rapid oxidation of dichlorodihydrofluorescin with heme and hemoproteins: Formation of the fluorescein is independent of the generation of reactive oxygen species. FEBS Lett. 2002, 511, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Taggart, L.E.; McMahon, S.J.; Currell, F.J.; Prise, K.M.; Butterworth, K.T. The role of mitochondrial function in gold nanoparticle mediated radiosensitisation. Cancer Nanotechnol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Khalil, T.T.; Bazzi, R.; Roux, S.; Fromm, M. The contribution of hydrogen peroxide to the radiosensitizing effect of gold nanoparticles. Colloids Surf. B Biointerfaces 2019, 175, 606–613. [Google Scholar] [CrossRef]
- Li, S.; Penninckx, S.; Karmani, L.; Heuskin, A.C.; Watillon, K.; Marega, R.; Zola, J.; Corvaglia, V.; Genard, G.; Gallez, B.; et al. LET-dependent radiosensitization effects of gold nanoparticles for proton irradiation. Nanotechnology 2016, 27, 455101. [Google Scholar] [CrossRef]
- Swanner, J.; Mims, J.; Carroll, D.L.; Akman, S.A.; Furdui, C.M.; Torti, S.V.; Singh, R.N. Differential cytotoxic and radiosensitizing effects of silver nanoparticles on triple-negative breast cancer and non-triple-negative breast cells. Int. J. Nanomed. 2015, 10, 3937–3953. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.T.; He, W.; Lo, Y.M.; Hu, X.; Wu, X.; Yin, J.J. Effect of silver nanomaterials on the activity of thiol-containing antioxidants. J. Agric. Food Chem. 2013, 61, 7855–7862. [Google Scholar] [CrossRef]
- Abdul Rashid, R.; Zainal Abidin, S.; Khairil Anuar, M.A.; Tominaga, T.; Akasaka, H.; Sasaki, R.; Kie, K.; Abdul Razak, K.; Pham, B.T.T.; Hawkett, B.S.; et al. Radiosensitization effects and ROS generation by high Z metallic nanoparticles on human colon carcinoma cell (HCT116) irradiated under 150 MeV proton beam. OpenNano 2019, 4, 100027. [Google Scholar] [CrossRef]
- Adams, W.T.; Nolan, M.W.; Ivanisevic, A. Ga Ion-Enhanced and Particle Shape-Dependent Generation of Reactive Oxygen Species in X-ray-Irradiated Composites. ACS Omega 2018, 3, 5252–5259. [Google Scholar] [CrossRef]
- Bouras, A.; Kaluzova, M.; Hadjipanayis, C.G. Radiosensitivity enhancement of radioresistant glioblastoma by epidermal growth factor receptor antibody-conjugated iron-oxide nanoparticles. J. Neuro-Oncol. 2015, 124. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.J.; Jung, K.O.; Graves, E.E.; Pratx, G. A gold nanoparticle system for the enhancement of radiotherapy and simultaneous monitoring of reactive-oxygen-species formation. Nanotechnology 2018, 29, 504001. [Google Scholar] [CrossRef]
- Colon, J.; Hsieh, N.; Ferguson, A.; Kupelian, P.; Seal, S.; Jenkins, D.W.; Baker, C.H. Cerium oxide nanoparticles protect gastrointestinal epithelium from radiation-induced damage by reduction of reactive oxygen species and upregulation of superoxide dismutase 2. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 698–705. [Google Scholar] [CrossRef]
- Molina Higgins, M.C.; Rojas, J.V. X-ray radiation enhancement of gold-TiO2 nanocomposites. Appl. Surf. Sci. 2019, 480, 1147–1155. [Google Scholar] [CrossRef]
- Jiang, Y.-W.; Gao, G.; Jia, H.-R.; Zhang, X.; Zhao, J.; Ma, N.; Liu, J.-B.; Liu, P.; Wu, F.-G. Copper Oxide Nanoparticles Induce Enhanced Radiosensitizing Effect via Destructive Autophagy. ACS Biomater. Sci. Eng. 2019, 5, 1569–1579. [Google Scholar] [CrossRef]
- Klein, S.; dell’Arciprete, M.L.; Wegmann, M.; Distel, L.V.R.; Neuhuber, W.; Gonzalez, M.C.; Kryschi, C. Oxidized silicon nanoparticles for radiosensitization of cancer and tissue cells. Biochem. Biophys. Res. Commun. 2013, 434, 217–222. [Google Scholar] [CrossRef]
- Lu, V.M.; Crawshay-Williams, F.; White, B.; Elliot, A.; Hill, M.A.; Townley, H.E. Cytotoxicity, dose-enhancement and radiosensitization of glioblastoma cells with rare earth nanoparticles. Artif. Cells Nanomed. Biotechnol. 2019, 47, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Wu, F.G.; Zhang, X.; Jiang, Y.W.; Jia, H.R.; Wang, H.Y.; Li, Y.H.; Liu, P.; Gu, N.; Chen, Z. Shape-Dependent Radiosensitization Effect of Gold Nanostructures in Cancer Radiotherapy: Comparison of Gold Nanoparticles, Nanospikes, and Nanorods. ACS Appl. Mater. Interfaces 2017, 9, 13037–13048. [Google Scholar] [CrossRef]
- Ma, S.; Miao, H.; Luo, Y.; Sun, Y.; Tian, X.; Wang, F.; You, C.; Peng, S.; Tang, G.; Yang, C.; et al. FePt/GO Nanosheets Suppress Proliferation, Enhance Radiosensitization and Induce Autophagy of Human Non-Small Cell Lung Cancer Cells. Int. J. Biol. Sci. 2019, 15, 999–1009. [Google Scholar] [CrossRef] [Green Version]
- Morita, K.; Miyazaki, S.; Numako, C.; Ikeno, S.; Sasaki, R.; Nishimura, Y.; Ogino, C.; Kondo, A. Characterization of titanium dioxide nanoparticles modified with polyacrylic acid and H2O2 for use as a novel radiosensitizer. Free Radic. Res. 2016, 50, 1319–1328. [Google Scholar] [CrossRef]
- Nakayama, M.; Sasaki, R.; Ogino, C.; Tanaka, T.; Morita, K.; Umetsu, M.; Ohara, S.; Tan, Z.; Nishimura, Y.; Akasaka, H.; et al. Titanium peroxide nanoparticles enhanced cytotoxic effects of X-ray irradiation against pancreatic cancer model through reactive oxygen species generation in vitro and in vivo. Radiat. Oncol. 2016, 11, 91. [Google Scholar] [CrossRef] [Green Version]
- Nicol, J.R.; Harrison, E.; O’Neill, S.M.; Dixon, D.; McCarthy, H.O.; Coulter, J.A. Unraveling the cell-type dependent radiosensitizing effects of gold through the development of a multifunctional gold nanoparticle. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.J.; Han, S.M.; Cho, J.H.; Hyodo, K.; Zaboronok, A.; You, H.; Peach, K.; Hill, M.A.; Kim, J.K. Enhanced production of reactive oxygen species by gadolinium oxide nanoparticles under core-inner-shell excitation by proton or monochromatic X-ray irradiation: Implication of the contribution from the interatomic de-excitation-mediated nanoradiator effect to dose enhancement. Radiat. Environ. Biophys. 2015, 54, 423–431. [Google Scholar] [CrossRef]
- Vasilieva, V.V.; Alyakov, M.; Apostolova, M.D. Nanoparticles Containing a Copper Chelator: A Possible Instrument for Radiation Protection. In Nanoscience Advances in CBRN Agents Detection, Information and Energy Security; Springer: Dordrecht, The Netherlands, 2015; pp. 423–436. [Google Scholar]
- Wu, H.; Lin, J.; Liu, P.; Huang, Z.; Zhao, P.; Jin, H.; Ma, J.; Wen, L.; Gu, N. Reactive oxygen species acts as executor in radiation enhancement and autophagy inducing by AgNPs. Biomaterials 2016, 101, 1–9. [Google Scholar] [CrossRef]
- Yong, Y.; Zhang, C.; Gu, Z.; Du, J.; Guo, Z.; Dong, X.; Xie, J.; Zhang, G.; Liu, X.; Zhao, Y. Polyoxometalate-Based Radiosensitization Platform for Treating Hypoxic Tumors by Attenuating Radioresistance and Enhancing Radiation Response. ACS Nano 2017, 11, 7164–7176. [Google Scholar] [CrossRef]
- Youkhana, E.Q.; Feltis, B.; Blencowe, A.; Geso, M. Titanium Dioxide Nanoparticles as Radiosensitisers: An In vitro and Phantom-Based Study. Int. J. Med. Sci. 2017, 14, 602–614. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Liu, T.; Du, Y.; Luo, Z.; Zheng, W.; Chen, T. X-ray-responsive selenium nanoparticles for enhanced cancer chemo-radiotherapy. Colloids Surf. B Biointerfaces 2016, 139, 180–189. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, H.; Yang, Y.; Zhang, C.; Dong, X.; Du, J.; Yan, L.; Zhang, G.; Gu, Z.; Zhao, Y. Tumor microenvironment-manipulated radiocatalytic sensitizer based on bismuth heteropolytungstate for radiotherapy enhancement. Biomaterials 2019, 189, 11–22. [Google Scholar] [CrossRef]
- Li, M.; Yin, J.-J.; Wamer, W.G.; Lo, Y.M. Mechanistic characterization of titanium dioxide nanoparticle-induced toxicity using electron spin resonance. J. Food Drug Anal. 2014, 22, 76–85. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Liu, Y.; Wamer, W.G.; Yin, J.-J. Electron spin resonance spectroscopy for the study of nanomaterial-mediated generation of reactive oxygen species. J. Food Drug Anal. 2014, 22, 49–63. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.; Manchester, L.C.; Qi, W. Biochemical reactivity of melatonin with reactive oxygen and nitrogen species. Cell Biochem. Biophys. 2001, 34, 237–256. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, S.; Zhou, P.; Liu, W.; Ni, F. Effects of Ligustrazine on DNA damage and apoptosis induced by irradiation. Environ. Toxicol. Pharmacol. 2013, 36, 1197–1206. [Google Scholar] [CrossRef]
- Babu, S.; Velez, A.; Wozniak, K.; Szydlowska, J.; Seal, S. Electron paramagnetic study on radical scavenging properties of ceria nanoparticles. Chem. Phys. Lett. 2007, 442, 405–408. [Google Scholar] [CrossRef]
- Yin, J.-J.; Lao, F.; Meng, J.; Fu, P.P.; Zhao, Y.; Xing, G.; Gao, X.; Sun, B.; Wang, P.C.; Chen, C.; et al. Inhibition of Tumor Growth by Endohedral Metallofullerenol Nanoparticles Optimized as Reactive Oxygen Species Scavenger. Mol. Pharmacol. 2008, 74, 1132–1140. [Google Scholar] [CrossRef]
- Zhang, H.; Meng, D.; Fu, B.; Fan, H.; Cai, R.; Fu, P.P.; Wu, X. Separation of charge carriers and generation of reactive oxygen species by TiO2 nanoparticles mixed with differently-coated gold nanorods under light irradiation. J. Environ. Sci. Health Part C 2019, 37, 81–98. [Google Scholar] [CrossRef]
- Yu, K.-N.; Yoon, T.-J.; Minai-Tehrani, A.; Kim, J.-E.; Park, S.J.; Jeong, M.S.; Ha, S.-W.; Lee, J.-K.; Kim, J.S.; Cho, M.-H. Zinc oxide nanoparticle induced autophagic cell death and mitochondrial damage via reactive oxygen species generation. Toxicol. In Vitro 2013, 27, 1187–1195. [Google Scholar] [CrossRef]
- Maggiorella, L.; Barouch, G.; Devaux, C.; Pottier, A.; Deutsch, E.; Bourhis, J.; Borghi, E.; Levy, L. Nanoscale radiotherapy with hafnium oxide nanoparticles. Future Oncol. 2012, 8, 1167–1181. [Google Scholar] [CrossRef]
- Marill, J.; Anesary, N.M.; Zhang, P.; Vivet, S.; Borghi, E.; Levy, L.; Pottier, A. Hafnium oxide nanoparticles: Toward an in vitropredictive biological effect? Radiat. Oncol. 2014, 9, 150. [Google Scholar] [CrossRef] [Green Version]
- Su, X.-Y.; Liu, P.-D.; Wu, H.; Gu, N. Enhancement of radiosensitization by metal-based nanoparticles in cancer radiation therapy. Cancer Biol. Med. 2014, 11, 86–91. [Google Scholar] [CrossRef]
- Yamada, M.; Foote, M.; Prow, T.W. Therapeutic gold, silver, and platinum nanoparticles. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 428–445. [Google Scholar] [CrossRef]
- Kwatra, D.; Venugopal, A.; Anant, S. Nanoparticles in radiation therapy: A summary of various approaches to enhance radiosensitization in cancer. Transl. Cancer Res. 2013, 2, 330–342. [Google Scholar]
- Liu, P.; Jin, H.; Guo, Z.; Ma, J.; Zhao, J.; Li, D.; Wu, H.; Gu, N. Silver nanoparticles outperform gold nanoparticles in radiosensitizing U251 cells in vitro and in an intracranial mouse model of glioma. Int. J. Nanomed. 2016, 11, 5003–5014. [Google Scholar] [CrossRef] [Green Version]
- Fakhimikabir, H.; Tavakoli, M.B.; Zarrabi, A.; Amouheidari, A.; Rahgozar, S. The role of folic acid-conjugated polyglycerol coated iron oxide nanoparticles on radiosensitivity with clinical electron beam (6 MeV) on human cervical carcinoma cell line: In vitro study. J. Photochem. Photobiol. B Biol. 2018, 182, 71–76. [Google Scholar] [CrossRef]
- Verry, C.; Dufort, S.; Barbier, E.L.; Montigon, O.; Peoc’h, M.; Chartier, P.; Lux, F.; Balosso, J.; Tillement, O.; Sancey, L.; et al. MRI-guided clinical 6-MV radiosensitization of glioma using a unique gadolinium-based nanoparticles injection. Nanomedicine 2016, 11, 2405–2417. [Google Scholar] [CrossRef] [Green Version]
- Bouziotis, P.; Stellas, D.; Thomas, E.; Truillet, C.; Tsoukalas, C.; Lux, F.; Tsotakos, T.; Xanthopoulos, S.; Paravatou-Petsotas, M.; Gaitanis, A.; et al. 68Ga-radiolabeled AGuIX nanoparticles as dual-modality imaging agents for PET/MRI-guided radiation therapy. Nanomedicine 2017, 12, 1561–1574. [Google Scholar] [CrossRef] [Green Version]
- Li H, Y.Z.; Liu, C.; Zeng, Y.P.; Hao, Y.H.; Gu, Y.; Wang, W.D.; Li, R. PEGylated ceria nanoparticles used for radioprotection on human liver cells under γ-ray irradiation. Free Radic. Biol. Med. 2015, 87, 26–35. [Google Scholar] [CrossRef]
- Asati, A.; Santra, S.; Kaittanis, C.; Perez, J.M. Surface-Charge-Dependent Cell Localization and Cytotoxicity of Cerium Oxide Nanoparticles. ACS Nano 2010, 4, 5321–5331. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Z.; Mainali, M.K.; Sinha, N.; Strack, G.; Altundal, Y.; Hao, Y.; Winningham, T.A.; Sajo, E.; Celli, J.; Ngwa, W. Potential of using cerium oxide nanoparticles for protecting healthy tissue during accelerated partial breast irradiation (APBI). Phys. Med. 2016, 32, 631–635. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Chen, K.; Ma, J.; Gao, F. Cerium oxide nanoparticles in cancer. Oncotargets Ther. 2014, 2014, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Her, S.; Jaffray, D.A.; Allen, C. Gold nanoparticles for applications in cancer radiotherapy: Mechanisms and recent advancements. Adv. Drug Deliv. Rev. 2017, 109, 84–101. [Google Scholar] [CrossRef]
- Cui, L.; Her, S.; Borst, G.R.; Bristow, R.G.; Jaffray, D.A.; Allen, C. Radiosensitization by gold nanoparticles: Will they ever make it to the clinic? Radiother. Oncol. 2017, 124, 344–356. [Google Scholar] [CrossRef]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-Dependent Cytotoxicity of Gold Nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef]
- Peukert, D.; Kempson, I.; Douglass, M.; Bezak, E. Gold nanoparticle enhanced proton therapy: A Monte Carlo simulation of the effects of proton energy, nanoparticle size, coating material, and coating thickness on dose and radiolysis yield. Med. Phys. 2019. [Google Scholar] [CrossRef]
- Jeremic, B.; Aguerri, A.R.; Filipovic, N. Radiosensitization by gold nanoparticles. Clin. Transl. Oncol. 2013, 15, 593–601. [Google Scholar] [CrossRef]
- Chithrani, B.D.; Ghazani, A.A.; Chan, W.C. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006, 6. [Google Scholar] [CrossRef]
- Yin, J.-J.; Liu, J.; Ehrenshaft, M.; Roberts, J.E.; Fu, P.P.; Mason, R.P.; Zhao, B. Phototoxicity of Nano Titanium Dioxides in HaCaT Keratinocytes—Generation of Reactive Oxygen Species and Cell Damage. Toxicol. Appl. Pharmacol. 2012, 263, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Albanese, A.; Chan, W.C.W. Effect of Gold Nanoparticle Aggregation on Cell Uptake and Toxicity. ACS Nano 2011, 5, 5478–5489. [Google Scholar] [CrossRef]
- Wang, M.; Thanou, M. Targeting nanoparticles to cancer. Pharmacol. Res. 2010, 62, 90–99. [Google Scholar] [CrossRef]
- Gilles, M.; Brun, E.; Sicard-Roselli, C. Gold nanoparticles functionalization notably decreases radiosensitization through hydroxyl radical production under ionizing radiation. Colloids Surf. B Biointerfaces 2014, 123, 770–777. [Google Scholar] [CrossRef]
- Yang, P.-H.; Sun, X.; Chiu, J.-F.; Sun, H.; He, Q.-Y. Transferrin-Mediated Gold Nanoparticle Cellular Uptake. Bioconjugate Chem. 2005, 16, 494–496. [Google Scholar] [CrossRef]
- Cheng, N.N.; Starkewolf, Z.; Davidson, R.A.; Sharmah, A.; Lee, C.; Lien, J.; Guo, T. Chemical Enhancement by Nanomaterials under X-ray Irradiation. J. Am. Chem. Soc. 2012, 134, 1950–1953. [Google Scholar] [CrossRef]
- Munaweera, I.; Koneru, B.; Shi, Y.; Pasqua, A.J.D.; Kenneth, J.; Balkus, J. Chemoradiotherapeutic wrinkled mesoporous silica nanoparticles for use in cancer therapy. Appl. Mater. 2014, 2, 113315. [Google Scholar] [CrossRef]
- Jain, S.; Hirst, D.G.; O’Sullivan, J.M. Gold nanoparticles as novel agents for cancer therapy. Br. J. Radiol. 2012, 85, 101–113. [Google Scholar] [CrossRef]
- Patra, C.R.; Bhattacharya, R.; Mukhopadhyay, D.; Mukherjee, P. Fabrication of gold nanoparticles for targeted therapy in pancreatic cancer. Adv. Drug Deliv. Rev. 2010, 62, 346–361. [Google Scholar] [CrossRef] [Green Version]
- Gargioni, E.; Schulz, F.; Raabe, A.; Burdak-Rothkamm, S.; Rieckmann, T.; Rothkamm, K. Targeted nanoparticles for tumour radiotherapy enhancement—The long dawn of a golden era? Ann. Transl. Med. 2016, 4, 523. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, T.; Thierry, B.; Kempson, I. A quantitative study of intercellular heterogeneity in gold nanoparticle uptake across multiple cell lines. Anal. Bioanal. Chem. 2019, 411, 7529–7538. [Google Scholar] [CrossRef]


{kind=link}
| Assay | Specificity |
|---|---|
| 2′,7′-dichlorofluorescein diacetate (DCFDA) | Non-specific for most ROS or nitrogen species [88,89] |
| 7-hydroxycoumarin | Hydroxyl radical from hydrogen peroxide [89] |
| Dihydrorhodamine (DHR) | Superoxide radical, peroxynitrite anion and hydroxyl radical [90,91] |
| 3′-(p-aminophenyl) fluorescein (APF) | Hydroxyl radical, hypochlorite or peroxynitrite anion [88,89] |
| Dihydroethidium (DHE) | Superoxide radical and hydroxyl radical [89,91] |
| Singlet oxygen sensor green | Singlet oxygen [92] |
| MitoSOX | Superoxide radical [91] |
| Authors | Type of Nanoparticle and Size | Measurement Method | Radiation Dose | Key Observations |
|---|---|---|---|---|
| Abdul Rashid et al. [101] | Gold nanospheres, superparamagnetic iron oxide NPs, platinum nanodiamonds and bismuth oxide nanorods Size: 1.9 nm, 15 nm, 42 nm, 70 nm respectively | DCFDA with HCT 116 cell line | 4 Gy from a 150 MeV proton beam | In order of sensitization enhancer ratio; SPIONs < AuNPs < PtNDs < BiNRs. This was reflected in ROS generation and suggested to be the main variable between different NPs |
| Adams et al. [102] | Gallium oxyhydroxide in an anisotropic and “orzo” shape Size: 53 nm and 49 nm respectively | DCFDA in vitro with PC12 cell line | Up to 10 Gy from a 6 MeV LINAC | Generation of ROS was related to the stability and structure of NPs. The less stable the NP, the greater ROS generation due to an increased number of metal ions and chemical interactions |
| Bouras et al. [103] | Superparamagnetic iron oxide conjugated with cetuximab Size: 10 nm core | DCFDA in vitro with U87MG cell line | 10 Gy from a 320 keV X-ray source | Cetuximab coated iron NPs had higher internalization and ROS generation compared to non-coated NPs |
| Chen et al. [33] | Hafnium-doped hydroxyapatite nanocrystal Size: 100 nm | DCFDA in vitro with A549 cell line | 5 Gy from a 662 keV gamma source | Radiolysis enhancement due to physical mechanisms. Suggested hafnium ions near intracellular organelles to promote ROS generation |
| Chen et al. [93] | Ceria coated with neogambogic acid Size: 3–5 nm before coating | DCFDA in vitro with MCF-7 cell line | 6 Gy from a 6 MeV LINAC | Ceria NPs promoted autophagy of tumour cells, while also contributing to radioprotection by inhibiting ROS due to Cs4+ |
| Choi et al. [104] | Pegylated gold NPs Size: 20 nm | Dihydrorhodamine in vitro with MDA-MB-231 cell line and in vivo in a murine model | 2–10 Gy from 320 kV X-ray source | Gold NPs functionalized with dihydrorhodamine was used to analyse ROS on the surface of the NP. |
| Colon et al. [105] | Cerium oxide NPs Size: Possibly 3–5 nm or 10–50 nm aggregates | DCFDA in vitro with CRL-1541 cell line | 20 Gy from a 160 keV X-ray source | Increased radioprotection by scavenging and regulating ROS by the increased ratio of Ce4+ and upregulation of superoxide dismutase 2 |
| Fang et al. [65] | Peptide templated gold nanoclusters Size: 3 nm | DCFDA in vitro with MCF-7 cell line | 4 Gy from a 160 keV X-ray source | Increased ROS generation and radiosensitization when NPs were targeted to mitochondria |
| Gilles et al. [74] | Uncoated gold NPs Size: 32.4 nm | 7-hydroxycoumarin in solution | 15 Gy from a 17.5 keV X-ray source | Physical mechanisms do not govern radiosensitization. Physico-chemical mechanisms and the interfacial water around NPs is important for ROS production and radiosensitization |
| Higgins et al. [106] | Titania NPs loaded with gold Size: 6.5 nm and 21.6 nm | Methylene Blue degradation in solution | 35 Gy/min from a 225 kV X-ray source | NPs displayed radiosensitization by radical generation. Smaller NPs have increased surface area and more catalytic sites for chemical interactions |
| Jeynes et al. [3] | Gold NPs conjugated with fetal bovine serum or TAT peptide Size: 60 nm and 80 nm respectively | DMSO in vitro with RT112 cell line | 5 Gy from a 250 kVp X-ray source and 5 Gy from a 3 MeV proton source | During X-ray irradiation with NPs, DMSO scavenged ROS. This was not seen with a proton experiment |
| Jiang et al. [107] | Copper oxide NPs Size: 5.4 nm | DCFDA with MCF-7 cell line | 6 MV X-ray source | Copper oxide NPs contributed to ROS generation and autophagy |
| Khalil et al. [97] | Citrate-coated gold NPs Size: 9 nm, 21 nm and 30 nm | DMSO and 2-amino-2-hydroxymethyl-1-3-propanediol in water | 11–89 Gy with a 1.5 keV cathode source | H2O2 was crucial in production of hydroxyl radicals, mediated by gold NPs. Radical scavengers confirmed higher ROS production with smaller gold core |
| Klein et al. [108] | Silicon coated with amino-silane Size: 1 nm | DCFDA in vitro with MCF-7 and 3T3 cell lines | 3 Gy from a 120 keV X-ray source | NPs enhanced mitochondrial membrane depolarization, provoking oxidative stress |
| Klein et al. [78] | Superparamagnetic iron oxide NPs uncoated and coated with citric or malic acid Size: 9–20 nm uncoated, 7–17 nm with citric acid coat and 6–16 nm for malic acid coat | DCFDA in vitro with MCF-7, Caco-2 and 3T3 cell lines | 1 Gy or 3 Gy from a 120 keV X-ray source | Internalization of the NPs into the mitochondria provoked oxidative stress under irradiation |
| Liu et al. [4] | Gold NPs with different coatings Size: 13 nm gold | DCFDA in vitro with A431 cell line | 10 Gy from a 6 MeV LINAC | NPs released nitrite ions upon irradiation to increase ROS generation due to nanoparticle coating |
| Lu et al. [109] | La2O3, CeO2, CeO2-Gd, Nd2O3, Nd2O3-Si, Gd2O3 Size: <100 nm for all | DCFDA with U-87 MG and Mo59K cell lines | 3 Gy from 250 keV source | Cell lines responded differently to NPs incubation and irradiation. Gd and Ce based NPs generated ROS |
| Ma et al. [110] | Gold nanospheres, nanospikes and nanorods Size: 53.2 nm nanospheres, 54.0 nm nanospikes and 50.2 nm nanorods | DCFDA in vitro with KB cell line | 4 Gy from a 6 MeV LINAC | Shape affected internalization. Unclear if increases in ROS generation were shape dependent or due to difference in internalization. Spheres were the most effective |
| Ma et al. [111] | FePt NPs in nanosheets Size: 3.05 nm particles and 500 nm nanosheet | DCFDA with H1975 cell line | 4 Gy from a 204 kV photon beam | The nanosheet inhibited cell proliferation and increased ROS generation. Once in the cytoplasm, FePt NPs were internalized in the mitochondria and lysosome |
| Misawa et al. [75] | Citrate-coated gold Size: 5–250 nm | 3′-(p-aminophenyl) fluorescein and dihydroethidium respectively and in solution | Up to 10 Gy from a 100 keV X-ray source | ROS generation was proportional with the inverse of the diameter of the nanoparticle |
| Morita et al. [112] | Polyacrylic acid-modified titanium dioxide with H2O2 Size: 124 nm | 3′-(p-aminophenyl) fluorescein in solution | Up to 18 Gy from an 80 keV X-ray source | H2O2 bound to surface and gradually released from nanoparticle surface, adding ROS |
| Nakayama et al. [113] | Titanium peroxide with coating of polyacrylic acid Size: 50–70 nm | 3′-(p-aminophenyl) fluorescein, DCFDA and dihydroethidium. Measured in solution and in vitro with MIA PaCa-2 cell line | Up to 30 Gy from a 150 keV X-ray source | Nanoparticle coating peroxidised into H2O2, catalysing ROS generation |
| Nicol et al. [114] | Gold NPs functionalized with peptides Size: 28.7 nm before peptides and 45.9 nm after | DCFDA in vitro with MDA-MB-231 and MCF-7 cell lines | 2 Gy from a 160 keV X-ray source | Nanoparticle coating inhibited SOD-2 expression and promotes cellular uptake, leaving cells susceptible to increased levels of ROS |
| Seo et al. [115] | Gadolinium oxide and gadolinium-chelate NPs Size: 40–45 nm | Dihydrorhodamine in vitro with CT26 cell line | Up to 15 Gy from a 45 MeV proton source | Gd ions from Gd-Gd de-excitation promoted ROS generation for radiosensitization |
| Shao et al. [72] | Hollow mesoporous silica NPs with sodium percarbonate in the cavity and coated with polyacrylic acid Size: 290 nm with 80 nm core | DCFDA in vitro with ZR-75-30 cell line | Unknown dose from a 60 keV X-ray source | NPs transported sodium percarbonate to the cancer microenvironment, increasing oxygen and generation of ROS |
| Taggart et al. [96] | Aurovist™ gold nanoparticles. Size: 1.9 nm | Nonyl-Acridine Orange in vitro with MDA-MB-231 and DU145 cell lines | 2 Gy with a 225 kV X-ray generator | Gold NPs and irradiation increased levels of ROS, leading to reduced mitochondrial membrane polarization |
| Vasilieva et al. [116] | Nanodiamonds conjugated with neocuproine Size: 6 nm | DCFDA in vitro with HepG2 cell line | 3 Gy from a 137Cs gamma source | NPs scavenged ROS but mechanisms are not well known |
| Wu et al. [117] | Silver coated with polyvinylpyrroliodone Size: 15.38 nm | DCFDA in vitro and MitoSOX (mitochondrial probe) with U251 cell line | No irradiation source used for ROS generation | Silver NPs increased inhibition of protective autophagy and ROS generation was increased |
| Yong et al. [118] | Gadolinium-containing polyoxometalates-conjugated chitosan Size: 30 nm | DCFDA in solution and in vitro with BEL-7402 cell line | 2 Gy from an unknown X-ray source | NPs reduced glutathione levels by redox reaction. Reduction of antioxidants lead to increased levels of ROS and oxidative stress |
| Youkhana et al. [119] | Anatase titanium oxide coated with aminopropyl trimethoxysilane Size: 30 nm | DCFDA in vitro with HaCaT and DU145 cell lines | 15 Gy and 14 Gy from a 6 MeV LINAC | ROS generation was dependent on the nanoparticle concentration |
| Yu et al. [120] | Selenium NPs coated with PEG Size: 500 nm | DCFDA in solution | 8 Gy from an unknown X-ray source | ROS generation using DCFDA was time dependent, decreasing intensity after 40 min. NPs contributed to ROS generation and degraded in cells |
| Zhou et al. [121] | Bismuth heteropolytungstate (BiP5W30) nanocluster Size: 1.5 nm | Terephthalic acid in solution. ELISA kit with human hydroxyl radical capture antibody in HeLa cell line. DCFDA was also used. | 50 kV with unknown X-ray source | Nanocluster promoted radiosensitization through physical and physico-chemical mechanisms. Depletion of glutathione by redox reactions, further promoting oxidative stress |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howard, D.; Sebastian, S.; Le, Q.V.-C.; Thierry, B.; Kempson, I. Chemical Mechanisms of Nanoparticle Radiosensitization and Radioprotection: A Review of Structure-Function Relationships Influencing Reactive Oxygen Species. Int. J. Mol. Sci. 2020, 21, 579. https://doi.org/10.3390/ijms21020579
Howard D, Sebastian S, Le QV-C, Thierry B, Kempson I. Chemical Mechanisms of Nanoparticle Radiosensitization and Radioprotection: A Review of Structure-Function Relationships Influencing Reactive Oxygen Species. International Journal of Molecular Sciences. 2020; 21(2):579. https://doi.org/10.3390/ijms21020579
Chicago/Turabian StyleHoward, Douglas, Sonia Sebastian, Quy Van-Chanh Le, Benjamin Thierry, and Ivan Kempson. 2020. "Chemical Mechanisms of Nanoparticle Radiosensitization and Radioprotection: A Review of Structure-Function Relationships Influencing Reactive Oxygen Species" International Journal of Molecular Sciences 21, no. 2: 579. https://doi.org/10.3390/ijms21020579