Protective Effect of the Aqueous Extract of Deschampsia antarctica (EDAFENCE®) on Skin Cells against Blue Light Emitted from Digital Devices

 ,
,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. EDA Promotes Cell Viability Reduced by Artificial Blue Light Exposure
2.2. EDA Prevents Oxidative Stress Induced by Artificial Blue Light Irradiation
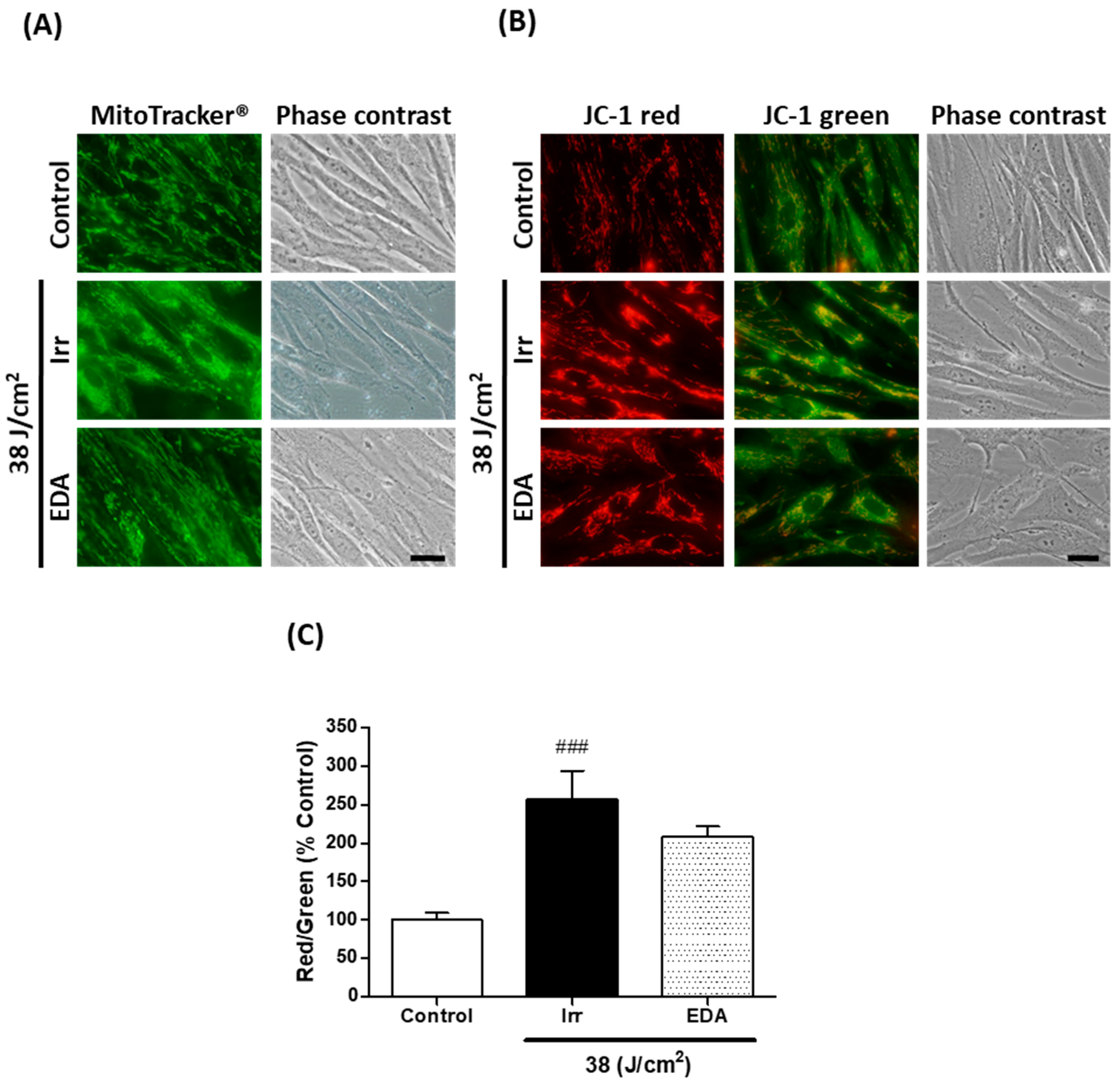
2.3. EDA Prevents Blue Light-Induced Alterations on Mitochondrial Morphology and Membrane Potential
2.4. EDA Regulates Artificial Blue Light-Induced Pigmentation through p38 Melanogenic Signalling Pathway Modulation
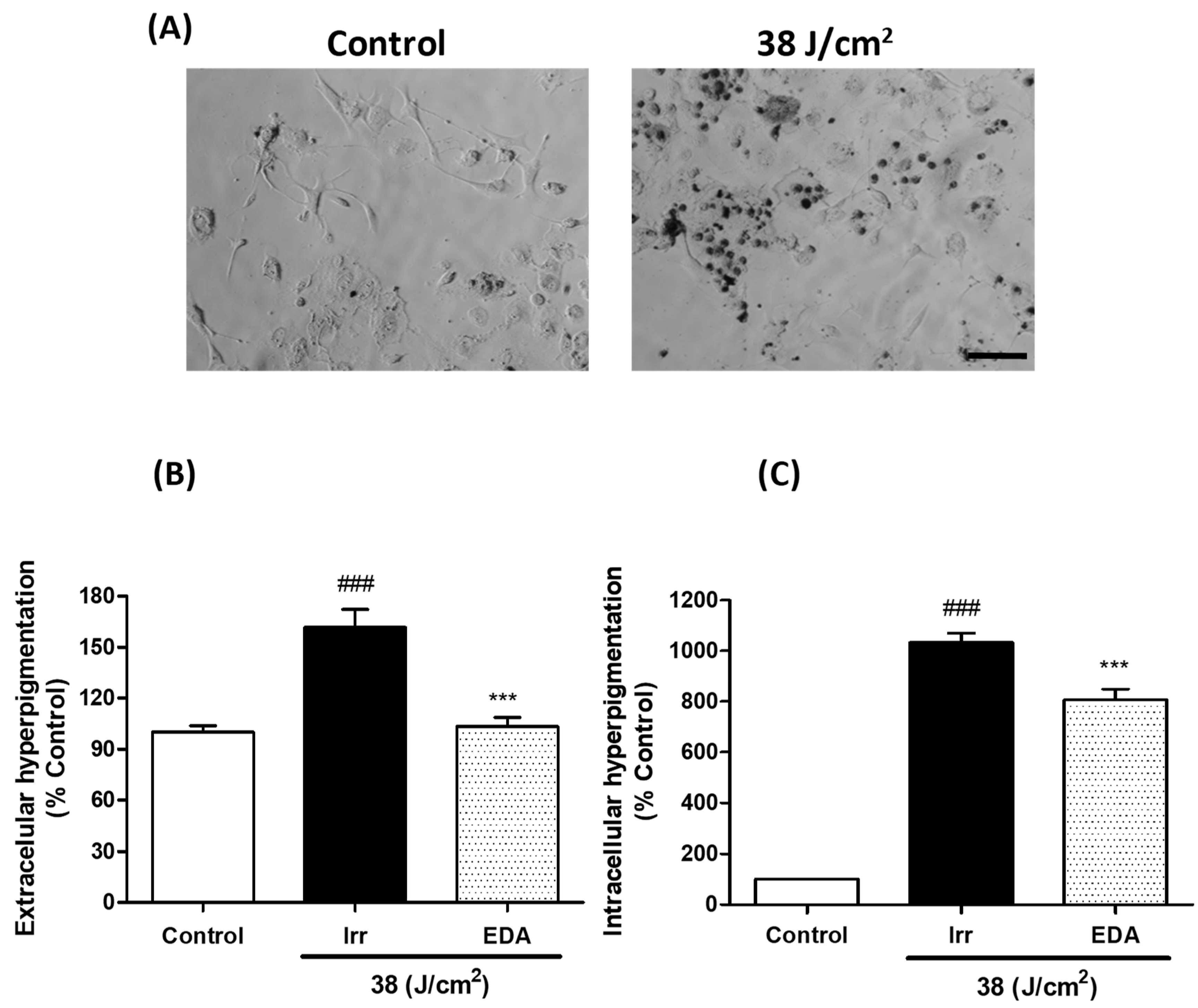
2.5. EDA Prevents Hyperpigmentation Induced by Artificial Blue Light
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Natural Extract and Cell Treatment
4.3. Irradiation
4.4. MTT Cell Viability Assay
4.5. Reactive Oxygen Species Measurement
4.6. Mitochondrial Morphology and Membrane Potential
4.7. Protein Electrophoresis and Western Blot
4.8. Melanin Content Determination
4.9. Microscopic Observation and Quantification
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CPD | Cyclobutane pyrimidine dimers |
| DHFDA | Dihydrofluorescein diacetate |
| DMEM | Dulbecco’s modified eagle medium |
| EDA | Extract from Deschampsia antarctica, EDAFENCE® |
| FBS | Fetal bovine serum |
| HDF | Human dermal fibroblasts |
| Irr | Irradiation |
| JC-1 | 5,5,6,6’-tetrachloro-1,1’,3,3’-tetraethylbenzimi-dazoylcarbocyanine iodide |
| LED | Light-emitted diode |
| MAPK | Mitogen-activated protein kinase |
| MITF | Microphthalmia-associated transcription factor |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazoliumbromide |
| ROS | Reactive oxygen species |
| SEM | Standard error of the mean |
| Trxs | Thioredoxins |
| UV | Ultraviolet |
| VIS | Visible light |
References
- Nguyen, A.V.; Soulika, A.M. The dynamics of the skin’s immune system. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef] [Green Version]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Parrado, C.; Mercado-Saenz, S.; Pérez-Davó, A.; Gilaberte, Y.; González, S.; Juarranz, A. Environmental Stressors on Skin Aging. Mechanistic Insights. Front. Pharmacol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rascalou, A.; Lamartine, J.; Poydenot, P.; Demarne, F.; Bechetoille, N. Mitochondrial damage and cytoskeleton reorganization in human dermal fibroblasts exposed to artificial visible light similar to screen-emitted light. J. Dermatol. Sci. 2018, 91, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Duteil, L.; Cardot-Leccia, N.; Queille-Roussel, C.; Maubert, Y.; Harmelin, Y.; Boukari, F.; Ambrosetti, D.; Lacour, J.P.; Passeron, T. Differences in visible light-induced pigmentation according to wavelengths: A clinical and histological study in comparison with UVB exposure. Pigment Cell Melanoma Res. 2014, 27, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Zamarrón, A.; Lorrio, S.; González, S.; Juarranz, A. Fernblock Prevents Dermal Cell Damage Induced by Visible and Infrared A Radiation. Int. J. Mol. Sci. 2018, 19, 2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennet, D.; Viswanath, B.; Kim, S.; An, J.H. An ultra-sensitive biophysical risk assessment of light effect on skin cells. Oncotarget 2017, 8, 47861–47875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godley, B.F.; Shamsi, F.A.; Liang, F.-Q.; Jarrett, S.G.; Davies, S.; Boulton, M. Blue Light Induces Mitochondrial DNA Damage and Free Radical Production in Epithelial Cells. J. Biol. Chem. 2005, 280, 21061–21066. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, Y.; Ohta, S.; Wolf, A.M. Blue light-induced oxidative stress in live skin. Free Radic. Biol. Med. 2017, 108, 300–310. [Google Scholar] [CrossRef]
- Liebel, F.; Kaur, S.; Ruvolo, E.; Kollias, N.; Southall, M.D. Irradiation of skin with visible light induces reactive oxygen species and matrix-degrading enzymes. J. Invest. Dermatol. 2012, 132, 1901–1907. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Zhang, Y.; Fong, C.-C.; Tsang, C.-H.; Yang, Z.; Yang, M. cDNA Microarray Analysis of Gene Expression Profiles in Human Fibroblast Cells Irradiated with Red Light. J. Invest. Dermatol. 2003, 120, 849–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamunuwa, G.; Karunaratne, V.; Karunaratne, D.N. Effect of Lipid Composition on In Vitro Release and Skin Deposition of Curcumin Encapsulated Liposomes. J. Nanomater. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liebmann, J.; Born, M.; Kolb-Bachofen, V. Blue-light irradiation regulates proliferation and differentiation in human skin cells. J. Invest. Dermatol. 2010, 130, 259–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opländer, C.; Hidding, S.; Werners, F.B.; Born, M.; Pallua, N.; Suschek, C.V. Effects of blue light irradiation on human dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2011, 103, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Denda, M.; Fuziwara, S. Visible radiation affects epidermal permeability barrier recovery: Selective effects of red and blue light. J. Invest. Dermatol. 2008, 128, 1335–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorbellini, E.; Rucco, M.; Rinaldi, F. Photodynamic and photobiological effects of light-emitting diode (LED) therapy in dermatological disease: an update. Lasers Med. Sci. 2018, 33, 1431–1439. [Google Scholar] [CrossRef] [Green Version]
- Passeron, T. The key question of irradiance when it comes to the effects of visible light in the skin. J. Dermatol. Sci. 2019, 93, 69–70. [Google Scholar] [CrossRef] [Green Version]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the Mitochondrial Membrane Potential Using the Cationic JC-1 Dye as a Sensitive Fluorescent Probe. BIO-PROTOCOL 2019, 9. [Google Scholar] [CrossRef]
- Wang, Y.; Viennet, C.; Robin, S.; Berthon, J.Y.; He, L.; Humbert, P. Precise role of dermal fibroblasts on melanocyte pigmentation. J. Dermatol. Sci. 2017, 88, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Stiglic, N.; Viner, R.M. Effects of screentime on the health and well-being of children and adolescents: a systematic review of reviews. BMJ Open 2019, 9, e023191. [Google Scholar] [CrossRef] [Green Version]
- Hölker, F.; Moss, T.; Griefahn, B.; Kloas, W.; Voigt, C.; Henckel, D.; Hänel, A.; Kappeler, P.M.; Völker, S.; Schwope, A.; et al. The Dark Side of Light: A Transdisciplinary Research Agenda for Light Pollution Policy. Ecol. Soc. 2010, 15, 13. [Google Scholar] [CrossRef]
- Flies, E.J.; Mavoa, S.; Zosky, G.R.; Mantzioris, E.; Williams, C.; Eri, R.; Brook, B.W.; Buettel, J.C. Urban-associated diseases: Candidate diseases, environmental risk factors, and a path forward. Environ. Int. 2019, 133, 105187. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; Liñares, G.G.; Bujjamer, J.M.; Gorojod, R.M.; Alcon, S.P.; Martínez, J.H.; Baldessari, A.; Grecco, H.E.; Kotler, M.L. Toxicity of blue led light and A2E is associated to mitochondrial dynamics impairment in ARPE-19 cells: implications for age-related macular degeneration. Arch. Toxicol. 2019, 93, 1401–1415. [Google Scholar] [CrossRef] [PubMed]
- Premi, S.; Wallisch, S.; Mano, C.M.; Weiner, A.B.; Bacchiocchi, A.; Wakamatsu, K.; Bechara, E.J.; Halaban, R.; Douki, T.; Brash, D.E. Photochemistry. Chemiexcitation of melanin derivatives induces DNA photoproducts long after UV exposure. Science 2015, 347, 842–847. [Google Scholar] [CrossRef]
- Regazzetti, C.; Sormani, L.; Debayle, D.; Bernerd, F.; Tulic, M.K.; De Donatis, G.M.; Chignon-Sicard, B.; Rocchi, S.; Passeron, T. Melanocytes Sense Blue Light and Regulate Pigmentation through Opsin-3. J. Invest. Dermatol. 2018, 138, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Hatao, M. Involvement of photooxidation of melanogenic precursors in prolonged pigmentation induced by ultraviolet A. J. Invest. Dermatol. 2004, 122, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Zamarrón, A.; Morel, E.; Lucena, S.R.; Mataix, M.; Pérez-Davó, A.; Parrado, C.; González, S. Extract of Deschampsia antarctica (EDA) Prevents Dermal Cell Damage Induced by UV Radiation and 2,3,7,8-Tetrachlorodibenzo-p-dioxin. Int. J. Mol. Sci. 2019, 20, 1356. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Davó, A.; Truchuelo, M.T.; Vitale, M.; González-Castro, J. Efficacy of an Antiaging Treatment Against Environmental Factors. J. Clin. Aesthet. Dermatol. 2019, 12, 65–70. [Google Scholar]
- Milani, M.; Hashtroody, B.; Piacentini, M.; Celleno, L. Skin protective effects of an antipollution, antioxidant serum containing Deschampsia antartica extract, ferulic acid and vitamin C: A controlled single-blind, prospective trial in women living in urbanized, high air pollution area. Clin. Cosmet. Investig. Dermatol. 2019, 12, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Espín, A.; Morel, E.; Juarranz, A.; Guerrero, A.; González, S.; Jiménez, A.; Sevilla, F. An Extract from the Plant Deschampsia antarctica Protects Fibroblasts from Senescence Induced by Hydrogen Peroxide. Oxidative Med. Cell. Longevity. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Viña, J.; Borras, C.; Abdelaziz, K.M.; García-Valles, R.; Gómez-Cabrera, M.C. The free radical theory of aging revisited: The cell signaling disruption theory of aging. Antioxid. Redox Signal. 2013, 19, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Zhou, H.J.; Zhang, H.; Huang, Y.; Hinojosa-Kirschenbaum, F.; Fan, P.; Yao, L.; Belardinelli, L.; Tellides, G.; Giordano, F.J.; et al. Thioredoxin-2 inhibits mitochondrial reactive oxygen species generation and apoptosis stress kinase-1 activity to maintain cardiac function. Circulation 2015, 131, 1082–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smalley, K.; Eisen, T. The involvement of p38 mitogen-activated protein kinase in the alpha-melanocyte stimulating hormone (alpha-MSH)-induced melanogenic and anti-proliferative effects in B16 murine melanoma cells. FEBS Lett. 2000, 476, 198–202. [Google Scholar] [CrossRef] [Green Version]
- Hirata, N.; Naruto, S.; Ohguchi, K.; Akao, Y.; Nozawa, Y.; Iinuma, M.; Matsuda, H. Mechanism of the melanogenesis stimulation activity of (-)-cubebin in murine B16 melanoma cells. Bioorganic Med. Chem. 2007, 15, 4897–4902. [Google Scholar] [CrossRef]
- Hempel, S.L.; Buettner, G.R.; O’Malley, Y.Q.; Wessels, D.A.; Flaherty, D.M. Dihydrofluorescein diacetate is superior for detecting intracellular oxidants: Comparison with 2’,7’-dichlorodihydrofluorescein diacetate, 5(and 6)-carboxy-2’,7’-dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic. Biol. Med. 1999, 27, 146–159. [Google Scholar] [CrossRef]
- Bellei, B.; Flori, E.; Izzo, E.; Maresca, V.; Picardo, M. GSK3β inhibition promotes melanogenesis in mouse B16 melanoma cells and normal human melanocytes. Cell. Signal. 2008, 20, 1750–1761. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorrio, S.; Rodríguez-Luna, A.; Delgado-Wicke, P.; Mascaraque, M.; Gallego, M.; Pérez-Davó, A.; González, S.; Juarranz, Á. Protective Effect of the Aqueous Extract of Deschampsia antarctica (EDAFENCE®) on Skin Cells against Blue Light Emitted from Digital Devices. Int. J. Mol. Sci. 2020, 21, 988. https://doi.org/10.3390/ijms21030988
Lorrio S, Rodríguez-Luna A, Delgado-Wicke P, Mascaraque M, Gallego M, Pérez-Davó A, González S, Juarranz Á. Protective Effect of the Aqueous Extract of Deschampsia antarctica (EDAFENCE®) on Skin Cells against Blue Light Emitted from Digital Devices. International Journal of Molecular Sciences. 2020; 21(3):988. https://doi.org/10.3390/ijms21030988
Chicago/Turabian StyleLorrio, Silvia, Azahara Rodríguez-Luna, Pablo Delgado-Wicke, Marta Mascaraque, María Gallego, Azahara Pérez-Davó, Salvador González, and Ángeles Juarranz. 2020. "Protective Effect of the Aqueous Extract of Deschampsia antarctica (EDAFENCE®) on Skin Cells against Blue Light Emitted from Digital Devices" International Journal of Molecular Sciences 21, no. 3: 988. https://doi.org/10.3390/ijms21030988