Exposure to Chemotherapy During Childhood or Adulthood and Consequences on Spermatogenesis and Male Fertility

 , and
, and
Abstract
:1. Introduction
2. Consequences of Chemotherapy Exposure During Childhood on Future Male Fertility
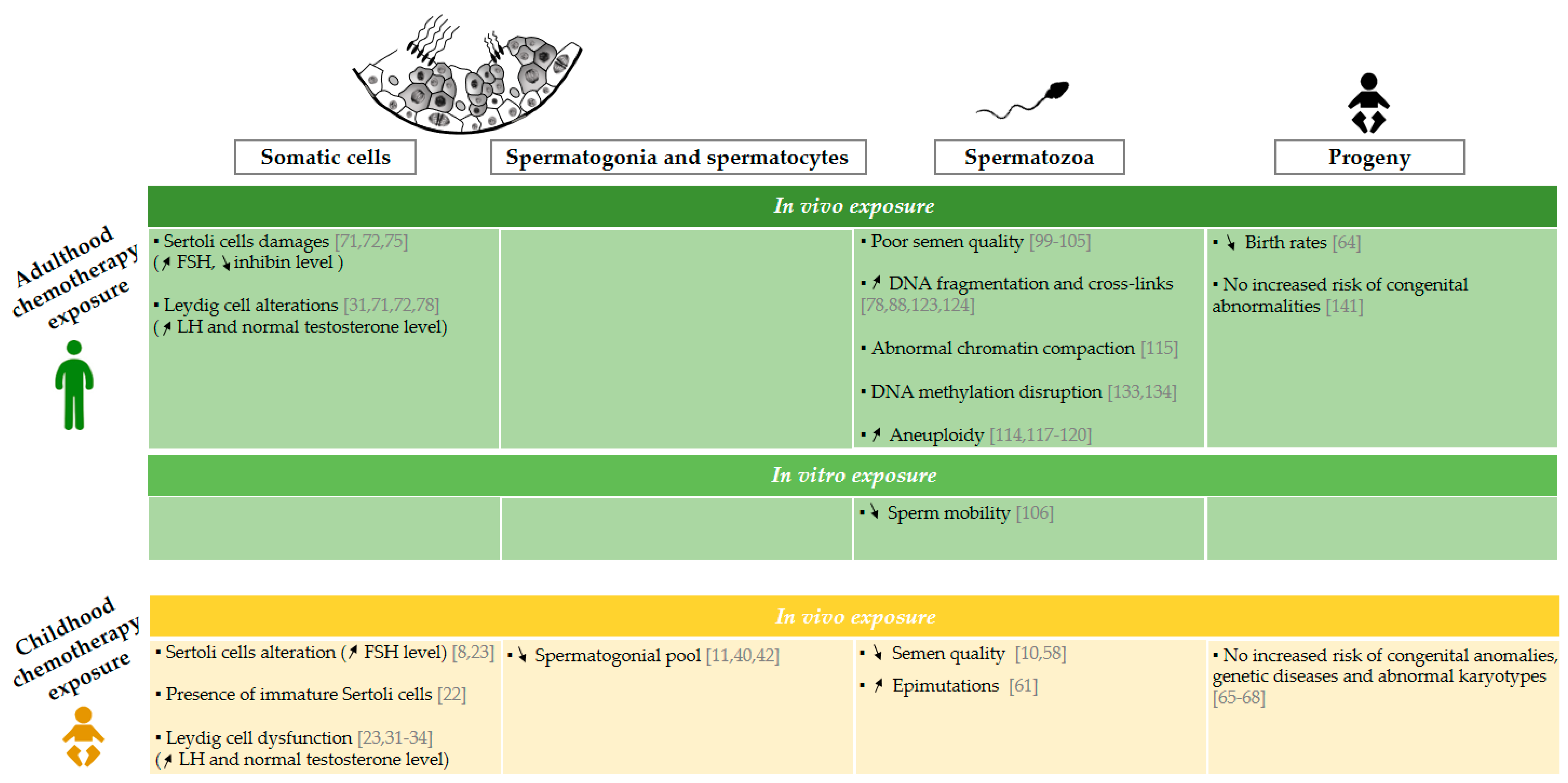
2.1. Effects of Chemotherapy on Somatic Cells and Spermatogenic Progression in Adulthood
2.1.1. Effects of Chemotherapy on Somatic Cells
2.1.2. Effects of Chemotherapy on Spermatogonia and Spermatocytes
2.2. Effects of Chemotherapy on Spermatozoa and Offspring
2.2.1. Sperm Production and Recovery Period
2.2.2. Sperm Nuclear Abnormalities
2.2.3. Effects of Chemotherapy on Cancer Survivors’ Offspring
3. Consequences of Chemotherapy Exposure During Adulthood on Male Fertility
3.1. Effects of Chemotherapy on Somatic Cells and Spermatogenesis Recovery
3.1.1. Effects of Chemotherapy on Somatic Cells
3.1.2. Effects of Chemotherapy on Spermatogonia and Spermatocytes
3.2. Effects of Chemotherapy on Spermatozoa and Offspring
3.2.1. Sperm Production and Recovery Period
3.2.2. Sperm Nuclear Abnormalities
3.2.3. Effects of Chemotherapy on Cancer Survivors’ Offspring
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MOPP | Mechloretamine, Vincristine, Procarbazine, and Prednisone |
| ABVD | Adriamycin, Bleomycin, Vinblastine, and Dacarbazine |
| CHOP | Cyclophosphamide, Doxorubicin, Vincristine, and Prednisolone |
| DNA | DeoxyriboNucleic Acid |
| SSCs | Spermatogonial Stem Cells |
| FSH | Follicle Stimulating Hormone |
| ABP | Androgen Binding Protein |
| LH | Luteinizing Hormone |
| γH2AX | γ Histone 2AX |
| dpp | day post-partum |
| CED | Cyclophosphamide Equivalent Dose |
| BEP | Bleomycin, Etoposide, and cisPlatin |
| CYP19A1 | Cytochrome P450 Family 19 Subfamily A Member 1 |
| AMH | Anti-Müllerian Hormone |
| CDKN1a | Cyclin-Dependent Kinase Inhibitor 1a |
| ROS | Reactive Oxygen Species |
| FISH | Fluorescence In Situ Hybridization |
| ABV | Adriamycin, Bleomycin, and Vinblastine |
| MOPP-ABV | Mechlorethamine, Vincristine, Procarbazine, Prednisone-Doxorubicin, Bleomycin, Vinblastine |
| TUNEL | TdT-Mediated dUTP Nickend Labelling Assay |
| BRDT | Testis-Specific Bromodomain-Protein |
| H3K9me | Histone H3 Monomethylated On Lysine 9 |
| H2B | Histone 2B |
| H3 | Histone 3 |
| H4 | Histone 4 |
| H19D | Histone 19 |
| ICSI | Intracytoplasmic Sperm Injection |
References
- DeSantis, C.E.; Lin, C.C.; Mariotto, A.B.; Siegel, R.L.; Stein, K.D.; Kramer, J.L.; Alteri, R.; Robbins, A.S.; Jemal, A. Cancer treatment and survivorship statistics, 2014. CA Cancer J. Clin. 2014, 64, 252–271. [Google Scholar] [CrossRef] [PubMed]
- Steliarova-Foucher, E.; Colombet, M.; Ries, L.A.G.; Moreno, F.; Dolya, A.; Bray, F.; Hesseling, P.; Shin, H.Y.; Stiller, C.A. IICC-3 contributors International incidence of childhood cancer, 2001–2010: A population-based registry study. Lancet Oncol. 2017, 18, 719–731. [Google Scholar] [CrossRef]
- Desandes, E.; Lacour, B.; Clavel, J. Les cancers des adolescents et des jeunes patients: Vision épidémiologique et organisations des soins en France. Bull. Du Cancer 2016, 103, 957–965. [Google Scholar] [CrossRef]
- Lacour, B.; Guyot-Goubin, A.; Guissou, S.; Bellec, S.; Désandes, E.; Clavel, J. Incidence of childhood cancer in France: National Children Cancer Registries, 2000–2004. Eur. J. Cancer Prev. 2010, 19, 173–181. [Google Scholar] [CrossRef]
- Benedict, C.; Shuk, E.; Ford, J. Fertility Issues in Adolescent and Young Adult Cancer Survivors. J. Adolesc. Young Adult Oncol. 2016, 5, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Brougham, M.F.H.; Wallace, W.H.B. Subfertility in children and young people treated for solid and haematological malignancies. Br. J. Haematol. 2005, 131, 143–155. [Google Scholar] [CrossRef]
- Wasilewski-Masker, K.; Seidel, K.D.; Leisenring, W.; Mertens, A.C.; Shnorhavorian, M.; Ritenour, C.W.; Stovall, M.; Green, D.M.; Sklar, C.A.; Armstrong, G.T.; et al. Male infertility in long-term survivors of pediatric cancer: A report from the childhood cancer survivor study. J. Cancer Surviv. 2014, 8, 437–447. [Google Scholar] [CrossRef] [Green Version]
- van Casteren, N.J.; van der Linden, G.H.M.; Hakvoort-Cammel, F.G.A.J.; Hählen, K.; Dohle, G.R.; van den Heuvel-Eibrink, M.M. Effect of childhood cancer treatment on fertility markers in adult male long-term survivors. Pediatr. Blood Cancer 2009, 52, 108–112. [Google Scholar] [CrossRef]
- Kenney, L.B.; Laufer, M.R.; Grant, F.D.; Grier, H.; Diller, L. High risk of infertility and long term gonadal damage in males treated with high dose cyclophosphamide for sarcoma during childhood. Cancer 2001, 91, 613–621. [Google Scholar] [CrossRef]
- Green, D.M.; Liu, W.; Kutteh, W.H.; Ke, R.W.; Shelton, K.C.; Sklar, C.A.; Chemaitilly, W.; Pui, C.-H.; Klosky, J.L.; Spunt, S.L.; et al. Cumulative alkylating agent exposure and semen parameters in adult survivors of childhood cancer: A report from the St Jude Lifetime Cohort Study. Lancet Oncol. 2014, 15, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Poganitsch-Korhonen, M.; Masliukaite, I.; Nurmio, M.; Lähteenmäki, P.; van Wely, M.; van Pelt, A.M.M.; Jahnukainen, K.; Stukenborg, J.-B. Decreased spermatogonial quantity in prepubertal boys with leukaemia treated with alkylating agents. Leukemia 2017, 31, 1460–1463. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Schover, L.R.; Partridge, A.H.; Patrizio, P.; Wallace, W.H.; Hagerty, K.; Beck, L.N.; Brennan, L.V.; Oktay, K. American Society of Clinical Oncology Recommendations on Fertility Preservation in Cancer Patients. JCO 2006, 24, 2917–2931. [Google Scholar] [CrossRef] [PubMed]
- Wyns, C.; Curaba, M.; Vanabelle, B.; Van Langendonckt, A.; Donnez, J. Options for fertility preservation in prepubertal boys. Hum. Reprod. Updat. 2010, 16, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.; Lopes, F.; Mitchell, R.T.; Spears, N. How does chemotherapy treatment damage the prepubertal testis? Reproduction 2018, 156, R209–R233. [Google Scholar] [CrossRef] [Green Version]
- Jahnukainen, K.; Ehmcke, J.; Hou, M.; Schlatt, S. Testicular function and fertility preservation in male cancer patients. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 287–302. [Google Scholar] [CrossRef]
- Heinrich, A.; DeFalco, T. Essential roles of interstitial cells in testicular development and function. Andrologia 2019. [Google Scholar] [CrossRef] [Green Version]
- Clermont, Y. Kinetics of spermatogenesis in mammals: Seminiferous epithelium cycle and spermatogonial renewal. Physiol. Rev. 1972, 52, 198–236. [Google Scholar] [CrossRef]
- Clermont, Y. Two classes of spermatogonial stem cells in the monkey (Cercopithecus aethiops). Am. J. Anat. 1969, 126, 57–71. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Shinohara, T. Spermatogonial stem cell self-renewal and development. Annu. Rev. Cell Dev. Biol. 2013, 29, 163–187. [Google Scholar] [CrossRef]
- Chemes, H.E. Infancy is not a quiescent period of testicular development. Int. J. Androl. 2001, 24, 2–7. [Google Scholar] [CrossRef]
- Stukenborg, J.-B.; Jahnukainen, K.; Hutka, M.; Mitchell, R.T. Cancer treatment in childhood and testicular function: The importance of the somatic environment. Endocr. Connect. 2018, 7, R69–R87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maymon, B.B.-S.; Yogev, L.; Marks, A.; Hauser, R.; Botchan, A.; Yavetz, H. Sertoli cell inactivation by cytotoxic damage to the human testis after cancer chemotherapy. Fertil. Steril. 2004, 81, 1391–1394. [Google Scholar] [CrossRef] [PubMed]
- Brämswig, J.H.; Heimes, U.; Heiermann, E.; Schlegel, W.; Nieschlag, E.; Schellong, G. The effects of different cumulative doses of chemotherapy on testicular function. Results in 75 patients treated for Hodgkin’s disease during childhood or adolescence. Cancer 1990, 65, 1298–1302. [Google Scholar] [CrossRef]
- Smart, E.; Lopes, F.; Rice, S.; Nagy, B.; Anderson, R.; Mitchell, R.; Spears, N. Chemotherapy drugs cyclophosphamide, cisplatin and doxorubicin induce germ cell loss in an in vitro model of the prepubertal testis. Sci. Rep. 2018, 8, 1773. [Google Scholar] [CrossRef] [Green Version]
- Aslani, F.; Sebastian, T.; Keidel, M.; Fröhlich, S.; Elsässer, H.-P.; Schuppe, H.-C.; Klug, J.; Mahavadi, P.; Fijak, M.; Bergmann, M.; et al. Resistance to apoptosis and autophagy leads to enhanced survival in Sertoli cells. MHR Basic Sci. Reprod. Med. 2017, 23, 370–380. [Google Scholar] [CrossRef]
- Tremblay, A.R.; Delbes, G. In vitro study of doxorubicin-induced oxidative stress in spermatogonia and immature sertoli cells. Toxicol. Appl. Pharmacol. 2018, 348, 32–42. [Google Scholar] [CrossRef]
- Brilhante, O.; Okada, F.K.; Sasso-Cerri, E.; Stumpp, T.; Miraglia, S.M. Late morfofunctional alterations of the Sertoli cell caused by doxorubicin administered to prepubertal rats. Reprod. Biol. Endocrinol. 2012, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Stumpp, T.; Freymuller, E.; Miraglia, S.M. Sertoli cell morphological alterations in albino rats treated with etoposide during prepubertal phase. Microsc. Microanal. 2008, 14, 225–235. [Google Scholar] [CrossRef]
- Velez de la Calle, J.F.; Soufir, J.C.; Chodorge, F.; Boisseau, C.; Kercret, H.; Jegou, B. Reproductive effects of the anti-cancer drug procarbazine in male rats at different ages. J. Reprod. Fertil. 1988, 84, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Stumpp, T.; Freymüller, E.; Miraglia, S.M. Sertoli cell function in albino rats treated with etoposide during prepubertal phase. Histochem. Cell. Biol. 2006, 126, 353–361. [Google Scholar] [CrossRef]
- Nurmio, M.; Toppari, J.; Kallio, J.; Hou, M.; Söder, O.; Jahnukainen, K. Functional in vitro model to examine cancer therapy cytotoxicity in maturing rat testis. Reprod. Toxicol. 2009, 27, 28–34. [Google Scholar] [CrossRef]
- Gerl, A.; Mühlbayer, D.; Hansmann, G.; Mraz, W.; Hiddemann, W. The impact of chemotherapy on Leydig cell function in long term survivors of germ cell tumors. Cancer 2001, 91, 1297–1303. [Google Scholar] [CrossRef]
- Heikens, J.; Behrendt, H.; Adriaanse, R.; Berghout, A. Irreversible gonadal damage in male survivors of pediatric Hodgkin’s disease. Cancer 1996, 78, 2020–2024. [Google Scholar] [CrossRef]
- Gerres, L.; Brämswig, J.H.; Schlegel, W.; Jürgens, H.; Schellong, G. The effects of etoposide on testicular function in boys treated for Hodgkin’s disease. Cancer 1998, 83, 2217–2222. [Google Scholar] [CrossRef]
- Talbot, J.A.; Shalet, S.M.; Tsatsoulis, A.; Grabinski, M.; Robertson, W.R. Luteinizing hormone pulsatility in men with damage to the germinal epithelium. Int. J. Androl. 1990, 13, 223–231. [Google Scholar] [CrossRef]
- Isaksson, S.; Bogefors, K.; Ståhl, O.; Eberhard, J.; Giwercman, Y.L.; Leijonhufvud, I.; Link, K.; Øra, I.; Romerius, P.; Bobjer, J.; et al. High risk of hypogonadism in young male cancer survivors. Clin. Endocrinol. 2018, 88, 432–441. [Google Scholar] [CrossRef]
- Pietila, S.; Mäkipernaa, A.; Koivisto, A.; Lenko, H. Growth impairment and gonadal axis abnormalities are common in survivors of paediatric brain tumours. Acta Paediatr. 2017, 106, 1684–1693. [Google Scholar] [CrossRef]
- Rafsanjani, K.A.; Faranoush, M.; Hedayatiasl, A.A.; Vossough, P. Gonadal function and fertility in male survivors treated for Hodgkin’s disease in Iran. Saudi Med. J. 2007, 28, 1690–1693. [Google Scholar]
- Hensle, T.; Burbige, K.; Shepard, B.; Marboe, C.; Blanc, W.; Wigger, J. Chemotherapy and its effect on testicular morphology in children. J. Urol. 1984, 131, 1142–1144. [Google Scholar] [CrossRef]
- Hyman, L.; Gilbert, E. Testicular atrophy in a prepubescent male after cyclophosphamide therapy. Lancet 1972, 2, 426–427. [Google Scholar] [CrossRef]
- Stukenborg, J.; Alves-Lopes, J.; Kurek, M.; Albalushi, H.; Reda, A.; Keros, V.; Töhönen, V.; Bjarnason, R.; Romerius, P.; Sundin, M.; et al. Spermatogonial quantity in human prepubertal testicular tissue collected for fertility preservation prior to potentially sterilizing therapy. Hum. Reprod. 2018, 33, 1677–1683. [Google Scholar] [CrossRef]
- Meistrich, M.L. Relationship between spermatogonial stem cell survival and testis function after cytotoxic therapy. Br. J. Cancer Suppl. 1986, 7, 89–101. [Google Scholar]
- Nurmio, M.; Keros, V.; Lähteenmäki, P.; Salmi, T.; Kallajoki, M.; Jahnukainen, K. Effect of Childhood Acute Lymphoblastic Leukemia Therapy on Spermatogonia Populations and Future Fertility. J. Clin. Endocrinol. Metab. 2009, 94, 2119–2122. [Google Scholar] [CrossRef] [Green Version]
- Freitas, F.E.L.; Cordeiro-mori, F.; Sasso-Cerri, E.; Lucas, S.R.R.; Miraglia, S.M. Alterations of spermatogenesis in etoposide-treated rats: A stereological study. Intercienca 2002, 27, 227–235. [Google Scholar]
- Clermont, Y.; Perey, B. Quantitative study of the cell population of the seminiferous tubules in immature rats. Am. J. Anat. 1957, 100, 241–267. [Google Scholar] [CrossRef]
- Drumond, A.L.; Meistrich, M.L.; Chiarini-Garcia, H. Spermatogonial morphology and kinetics during testis development in mice: A high-resolution light microscopy approach. Reproduction 2011, 142, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Lopes, F.; Smith, R.; Nash, S.; Mitchell, R.T.; Spears, N. Irinotecan metabolite SN38 results in germ cell loss in the testis but not in the ovary of prepubertal mice. Mol. Hum. Reprod. 2016, 22, 745–755. [Google Scholar] [CrossRef]
- Basciani, S.; De Luca, G.; Dolci, S.; Brama, M.; Arizzi, M.; Mariani, S.; Rosano, G.; Spera, G.; Gnessi, L. Platelet-Derived Growth Factor Receptor β-Subtype Regulates Proliferation and Migration of Gonocytes. Endocrinology 2008, 149, 6226–6235. [Google Scholar] [CrossRef] [Green Version]
- Nurmio, M.; Toppari, J.; Zaman, F.; Andersson, A.-M.; Paranko, J.; Söder, O.; Jahnukainen, K. Inhibition of tyrosine kinases PDGFR and C-Kit by imatinib mesylate interferes with postnatal testicular development in the rat. Int. J. Androl. 2007, 30, 366–376. [Google Scholar] [CrossRef]
- Culty, M. Gonocytes, the forgotten cells of the germ cell lineage. Birth Defects Res. Part C Embryo Today Rev. 2009, 87, 1–26. [Google Scholar] [CrossRef]
- Huckins, C.; Clermont, Y. Evolution of gonocytes in the rat testis during late embryonic and early post-natal life. Arch. Anat. Histol. Embryol. 1968, 51, 341–354. [Google Scholar] [PubMed]
- Heim, C.; Minniear, K.; Dann, C.T. Imatinib has deleterious effects on differentiating spermatogonia while sparing spermatogonial stem cell self renewal. Reprod. Toxicol. 2011, 31, 454–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaud, H.; van Pelt, A.; Delbes, G. Doxorubicin and vincristine affect undifferentiated rat spermatogonia. Reproduction 2017, 153, 725–735. [Google Scholar] [CrossRef]
- Liu, M.; Hales, B.F.; Robaire, B. Effects of four chemotherapeutic agents, bleomycin, etoposide, cisplatin, and cyclophosphamide, on DNA damage and telomeres in a mouse spermatogonial cell line. Biol. Reprod. 2014, 90, 72. [Google Scholar] [CrossRef] [Green Version]
- de Lange, T. Protection of mammalian telomeres. Oncogene 2002, 21, 532–540. [Google Scholar] [CrossRef] [Green Version]
- Stumpp, T.; Sasso-Cerri, E.; Freymuller, E.; Miraglia, S. Apoptosis and testicular alterations in albino rats treated with etoposide during the prepubertal phase. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2004, 279, 611–622. [Google Scholar] [CrossRef]
- Vendramini, V.; Robaire, B.; Miraglia, S.M. Amifostine-doxorubicin association causes long-term prepubertal spermatogonia DNA damage and early developmental arrest. Hum. Reprod. 2012, 27, 2457–2466. [Google Scholar] [CrossRef] [Green Version]
- Green, D.M.; Nolan, V.G.; Goodman, P.J.; Whitton, J.A.; Srivastava, D.; Leisenring, W.M.; Neglia, J.P.; Sklar, C.A.; Kaste, S.C.; Hudson, M.M.; et al. The cyclophosphamide equivalent dose as an approach for quantifying alkylating agent exposure: A report from the Childhood Cancer Survivor Study. Pediatr. Blood Cancer 2014, 61, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Romerius, P.; Ståhl, O.; Moëll, C.; Relander, T.; Cavallin Ståhl, E.; Wiebe, T.; Giwercman, Y.L.; Giwercman, A. High risk of azoospermia in men treated for childhood cancer. Int. J. Androl. 2011, 34, 69–76. [Google Scholar] [CrossRef]
- Romerius, P.; Stahl, O.; Moell, C.; Relander, T.; Cavallin-Stahl, E.; Gustafsson, H.; Lofvander Thapper, K.; Jepson, K.; Spano, M.; Wiebe, T.; et al. Sperm DNA Integrity in Men Treated for Childhood Cancer. Clin. Cancer Res. 2010, 16, 3843–3850. [Google Scholar] [CrossRef] [Green Version]
- Thomson, A.; Campbell, A.; Irvine, D.; Anderson, R.; Kelnar, C.J.; Wallace, W.H. Semen quality and spermatozoal DNA integrity in survivors of childhood cancer: A case-control study. Lancet 2002, 360, 361–367. [Google Scholar] [CrossRef]
- Shnorhavorian, M.; Schwartz, S.M.; Stansfeld, B.; Sadler-Riggleman, I.; Beck, D.; Skinner, M.K. Differential DNA Methylation Regions in Adult Human Sperm following Adolescent Chemotherapy: Potential for Epigenetic Inheritance. PLoS ONE 2017, 12, e0170085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, G.; Vadinkar, A.; Nair, S.; Kalthur, S.; D’Souza, A.; Shetty, P.; Mutalik, S.; Shetty, M.; Kalthur, G.; Adiga, S. Sperm abnormalities induced by pre-pubertal exposure to cyclophosphamide are effectively mitigated by Moringa oleifera leaf extract. Andrologia 2016, 48, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Chow, E.J.; Stratton, K.L.; Leisenring, W.M.; Oeffinger, K.C.; Sklar, C.A.; Donaldson, S.S.; Ginsberg, J.P.; Kenney, L.B.; Levine, J.M.; Robison, L.L.; et al. Pregnancy after chemotherapy in male and female survivors of childhood cancer treated between 1970 and 1999: A report from the Childhood Cancer Survivor Study cohort. Lancet Oncol. 2016, 17, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.-W.; Liu, J.; Juay, L.; Czene, K.; Miao, H.; Salim, A.; Verkooijen, H.M.; Hartman, M. Birth rates among male cancer survivors and mortality rates among their offspring: A population-based study from Sweden. BMC Cancer 2016, 16, 196. [Google Scholar] [CrossRef] [Green Version]
- Meistrich, M.L.; Byrne, J. Genetic disease in offspring of long-term survivors of childhood and adolescent cancer treated with potentially mutagenic therapies. Am. J. Hum. Genet. 2002, 70, 1069–1071. [Google Scholar] [CrossRef] [Green Version]
- Seppänen, V.; Artama, M.; Malila, N.; Pitkäniemi, J.; Rantanen, M.; Ritvanen, A.; Madanat-Harjuoja, L. Risk for congenital anomalies in offspring of childhood, adolescent and young adult cancer survivors. Int. J. Cancer 2016, 139, 1721–1730. [Google Scholar] [CrossRef]
- Signorello, L.B.; Mulvihill, J.J.; Green, D.M.; Munro, H.M.; Stovall, M.; Weathers, R.E.; Mertens, A.C.; Whitton, J.A.; Robison, L.L.; Boice, J.D. Congenital anomalies in the children of cancer survivors: A report from the childhood cancer survivor study. J. Clin. Oncol. 2012, 30, 239–245. [Google Scholar] [CrossRef]
- Winther, J.; Boice, J.J.; Christensen, J.; Frederiksen, K.; Mulvihill, J.J.; Stovall, M.; Olsen, J. Hospitalizations among children of survivors of childhood and adolescent cancer: A population-based cohort study. Int. J. Cancer 2010, 127, 2879–2887. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, A.; Beaud, H.; Delbès, G. Effets transgénérationnels des chimiothérapies: l’exposition du père influence-t-elle la santé des générations futures ? Gynécologie Obstétrique Fertilité Sénologie 2017, 45, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Blasco, M.; Trimarchi, J.; Keefe, D. An essential role for functional telomeres in mouse germ cells during fertilization and early development. Dev. Biol. 2002, 249, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remenár, E.; Számel, I.; Budai, B.; Vincze, B.; Gaudi, I.; Gundy, S.; Kásler, M. Increase of hypophyseal hormone levels in male head and neck cancer patients. Pathol. Oncol. Res. 2007, 13, 341–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spermon, J.; Ramos, L.; Wetzels, A.; Sweep, C.; Braat, D.; Kiemeney, L.; Witjes, J. Sperm integrity pre- and post-chemotherapy in men with testicular germ cell cancer. Hum. Reprod. 2006, 21, 1781–1786. [Google Scholar] [CrossRef] [PubMed]
- Giagulli, V.A.; Vermeulen, A. Leydig cell function in infertile men with idiopathic oligospermic infertility. J. Clin. Endocrinol. Metab. 1988, 66, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Franchimont, P.; Millet, D.; Vendrely, E.; Letawe, J.; Legros, J.J.; Netter, A. Relationship between Spermatogenesis and Serum Gonadotropin Levels in Azoospermia and Oligospermia. J. Clin. Endocrinol. Metab. 1972, 34, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Tsatsoulis, A.; Shalet, S.M.; Morris, I.D.; de Kretser, D.M. Immunoactive Inhibin as a Marker of Sertoli Cell Function following Cytotoxic Damage to the Human Testis. HRP 1990, 34, 254–259. [Google Scholar] [CrossRef]
- Wallace, E.M.; Groome, N.P.; Riley, S.C.; Parker, A.C.; Wu, F.C.W. Effects of Chemotherapy-Induced Testicular Damage on Inhibin, Gonadotropin, and Testosterone Secretion: A Prospective Longitudinal Study. J. Clin. Endocrinol. Metab. 1997, 82, 3111–3115. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, X.; Li, S.; Wang, H.; Yu, L.; Wang, P. The Effects of l-Carnitine Against Cyclophosphamide-Induced Injuries in Mouse Testis. Basic Clin. Pharmacol. Toxicol. 2017, 120, 152–158. [Google Scholar] [CrossRef]
- Howell, S.J.; Radford, J.A.; Ryder, W.D.J.; Shalet, S.M. Testicular Function After Cytotoxic Chemotherapy: Evidence of Leydig Cell Insufficiency. J. Clin. Oncol. 1999, 17, 1493–1498. [Google Scholar] [CrossRef]
- Setchell, B.; Galil, K. Limitations imposed by testicular blood flow on the function of Leydig cells in rats in vivo. Aust. J. Biol. Sci. 1983, 36, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Howell, S.J.; Shalet, S.M. Testicular function following chemotherapy. Hum. Reprod. Updat. 2001, 7, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bader, M.; Kilarkaje, N. Effects of bleomycin, etoposide and cisplatin treatment on Leydig cell structure and transcription of steroidogenic enzymes in rat testis. Eur. J. Pharmacol. 2015, 747, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Mossadegh-Keller, N.; Sieweke, M.H. Testicular macrophages: Guardians of fertility. Cell Immunol. 2018, 330, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Sasso-Cerri, E.; Oliveira, B.; de Santi, F.; Beltrame, F.L.; Caneguim, B.H.; Cerri, P.S. The antineoplastic busulphan impairs peritubular and Leydig cells, and vitamin B 12 stimulates spermatogonia proliferation and prevents busulphan-induced germ cell death. Biomed. Pharmacother. 2017, 95, 1619–1630. [Google Scholar] [CrossRef] [Green Version]
- Levi, M.; Hasky, N.; Stemmer, S.M.; Shalgi, R.; Ben-Aharon, I. Anti-Müllerian Hormone Is a Marker for Chemotherapy-Induced Testicular Toxicity. Endocrinology 2015, 156, 3818–3827. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, C.; Hamdane, M.; Mitchell, V.; Mayo, K.; Devisme, L.; Rigot, J.M.; Beauvillain, J.C.; Hermand, E.; Defossez, A. Immunolocalization of Inhibin and Activin α and βB Subunits and Expression of Corresponding Messenger RNAs in the Human Adult Testis. Biol. Reprod. 2003, 68, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Steger, K.; Rey, R.; Kliesch, S.; Louis, F.; Schleicher, G.; Bergmann, M. Immunohistochemical detection of immature Sertoli cell markers in testicular tissue of infertile adult men: A preliminary study. Int. J. Androl. 1996, 19, 122–128. [Google Scholar] [CrossRef]
- Peigné, M.; Decanter, C. Serum AMH level as a marker of acute and long-term effects of chemotherapy on the ovarian follicular content: A systematic review. Reprod. Biol. Endocrinol. 2014, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, C.; Hales, B.F.; Chan, P.; Robaire, B. Impact of chemotherapeutics and advanced testicular cancer or Hodgkin lymphoma on sperm deoxyribonucleic acid integrity. Fertil. Steril. 2010, 94, 1374–1379. [Google Scholar] [CrossRef]
- Marcon, L.; Zhang, X.; Hales, B.F.; Robaire, B.; Nagano, M.C. Effects of chemotherapeutic agents for testicular cancer on rat spermatogonial stem/progenitor cells. J. Androl. 2011, 32, 432–443. [Google Scholar] [CrossRef]
- van Pelt, A.; Roepers-Gajadien, H.; Gademan, I.; Creemers, L.; de Rooij, D.; van Dissel-Emiliani, F. Establishment of cell lines with rat spermatogonial stem cell characteristics. Endocrinology 2002, 143, 1845–1850. [Google Scholar] [CrossRef] [PubMed]
- Kotzur, T.; Benavides-Garcia, R.; Mecklenburg, J.; Sanchez, J.R.; Reilly, M.; Hermann, B.P. Granulocyte colony-stimulating factor (G-CSF) promotes spermatogenic regeneration from surviving spermatogonia after high-dose alkylating chemotherapy. Reprod. Biol. Endocrinol. 2017, 15, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadnejad, D.; Abedelahi, A.; Soleimani-rad, J.; Mohammadi-roshandeh, A.; Rashtbar, M.; Azami, A. Degenerative Effect of Cisplatin on Testicular Germinal Epithelium. Adv. Pharm. Bull. 2012, 2, 173–177. [Google Scholar] [PubMed]
- Azab, S.S.; kamel, I.; Ismail, N.N.; El Din Hosni, H.; El Fatah, M.A. The defensive role of taurine against gonadotoxicity and testicular apoptosis effects induced by cisplatin in rats. J. Infect. Chemother. 2020, 26, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.A.; Refaie, M.M.M.; Abdelghany, M.I. Naringenin palliates cisplatin and doxorubicin gonadal toxicity in male rats. Toxicol. Mech. Methods 2019, 29, 67–73. [Google Scholar] [CrossRef]
- Yeh, Y.-C.; Lai, H.-C.; Ting, C.-T.; Lee, W.-L.; Wang, L.-C.; Wang, K.-Y.; Lai, H.-C.; Liu, T.-J. Protection by doxycycline against doxorubicin-induced oxidative stress and apoptosis in mouse testes. Biochem. Pharmacol. 2007, 74, 969–980. [Google Scholar] [CrossRef]
- Cai, L.; Hales, B.F.; Robaire, B. Induction of Apoptosis in the Germ Cells of Adult Male Rats after Exposure to Cyclophosphamide1. Biol. Reprod. 1997, 56, 1490–1497. [Google Scholar] [CrossRef] [Green Version]
- Delbès, G.; Chan, D.; Pakarinen, P.; Trasler, J.M.; Hales, B.F.; Robaire, B. Impact of the Chemotherapy Cocktail Used to Treat Testicular Cancer on the Gene Expression Profile of Germ Cells from Male Brown-Norway Rats. Biol. Reprod. 2009, 80, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, F.; Bishop, J.B.; Lowe, X.; Generoso, W.M.; Hozier, J.; Wyrobek, A.J. Etoposide induces heritable chromosomal aberrations and aneuploidy during male meiosis in the mouse. Proc. Natl. Acad. Sci. USA 2001, 98, 3952–3957. [Google Scholar] [CrossRef] [Green Version]
- Bahadur, G.; Ozturk, O.; Muneer, A.; Wafa, R.; Ashraf, A.; Jaman, N.; Patel, S.; Oyede, A.W.; Ralph, D.J. Semen quality before and after gonadotoxic treatment. Hum. Reprod. 2005, 20, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Paoli, D.; Rizzo, F.; Fiore, G.; Pallotti, F.; Pulsoni, A.; Annechini, G.; Lombardo, F.; Lenzi, A.; Gandini, L. Spermatogenesis in Hodgkin’s lymphoma patients: A retrospective study of semen quality before and after different chemotherapy regimens. Hum. Reprod. 2016, 31, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Bujan, L.; Walschaerts, M.; Moinard, N.; Hennebicq, S.; Saias, J.; Brugnon, F.; Auger, J.; Berthaut, I.; Szerman, E.; Daudin, M.; et al. Impact of chemotherapy and radiotherapy for testicular germ cell tumors on spermatogenesis and sperm DNA: A multicenter prospective study from the CECOS network. Fertil. Steril. 2013, 100, 673–680.e2. [Google Scholar] [CrossRef] [PubMed]
- Meistrich, M.L.; Wilson, G.; Brown, B.W.; da Cunha, M.F.; Lipshultz, L.I. Impact of cyclophosphamide on long-term reduction in sperm count in men treated with combination chemotherapy for Ewing and soft tissue sarcomas. Cancer 1992, 70, 2703–2712. [Google Scholar] [CrossRef] [Green Version]
- Chapman, R.M.; Sutcliffe, S.B.; Rees, L.H.; Edwards, C.R.; Malpas, J.S. Cyclical combination chemotherapy and gonadal function. Retrospective study in males. Lancet 1979, 1, 285–289. [Google Scholar] [CrossRef]
- Marmor, D.; Duyck, F. Male reproductive potential after MOPP therapy for Hodgkin’s disease: A long-term survey. Andrologia 1994, 27, 99–106. [Google Scholar] [CrossRef]
- Tal, R.; Botchan, A.; Hauser, R.; Yogev, L.; Paz, G.; Yavetz, H. Follow-up of sperm concentration and motility in patients with lymphoma. Hum. Reprod. 2000, 15, 1985–1988. [Google Scholar] [CrossRef] [Green Version]
- Hong, C.Y.; Chiang, B.N.; Ku, J.; Wu, P. Screening the in vitro Sperm-immobilizing Effect of some Anticancer Drugs. Hum. Toxicol. 1985, 4, 461–464. [Google Scholar] [CrossRef]
- Calamera, J.C.; Morgenfeld, M.C.; Mancini, R.E.; Vilar, O. Biochemical Changes of the Human Semen Produced by Chlorambucil, Testosterone Propionate and Human Chorionic Gonadotropin Administration. Andrologia 1979, 11, 43–50. [Google Scholar] [CrossRef]
- Bieber, A.M.; Marcon, L.; Hales, B.F.; Robaire, B. Effects of chemotherapeutic agents for testicular cancer on the male rat reproductive system, spermatozoa, and fertility. J. Androl. 2006, 27, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Makino, S.; Kimura, H.; Ota, T.; Furuhashi, T.; Nagamura, Y. Sperm motion analysis in rats treated with adriamycin and its applicability to male reproductive toxicity studies. J. Toxicol. Sci. 2001, 26, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaisheva, F.; Delbes, G.; Hales, B.F.; Robaire, B. Effects of the chemotherapeutic agents for non-Hodgkin lymphoma, cyclophosphamide, doxorubicin, vincristine, and prednisone (CHOP), on the male rat reproductive system and progeny outcome. J. Androl. 2007, 28, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Delbès, G.; Vaisheva, F.; Luu, T.; Marcon, L.; Hales, B.F.; Robaire, B. Reversibility of the effects of the chemotherapeutic regimen for non-Hodgkin lymphoma, cyclophosphamide, doxorubicin, vincristine, and prednisone, on the male rat reproductive system and progeny outcome. Reprod. Toxicol. 2010, 29, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Beaud, H.; Tremblay, A.R.; Chan, P.T.K.; Delbes, G. Sperm DNA Damage in Cancer Patients. Adv. Exp. Med. Biol. 2019, 1166, 189–203. [Google Scholar]
- Bujan, L.; Walschaerts, M.; Brugnon, F.; Daudin, M.; Berthaut, I.; Auger, J.; Saias, J.; Szerman, E.; Moinard, N.; Rives, N.; et al. Impact of lymphoma treatments on spermatogenesis and sperm deoxyribonucleic acid: A multicenter prospective study from the CECOS network. Fertil. Steril. 2014, 102, 667–674. [Google Scholar] [CrossRef]
- Rives, N.; Walschaerts, M.; Setif, V.; Hennebicq, S.; Saias, J.; Brugnon, F.; Auger, J.; Berthaut, I.; Szerman, E.; Daudin, M.; et al. Sperm aneuploidy after testicular cancer treatment: Data from a prospective multicenter study performed within the French Centre d’Étude et de Conservation des Oeufs et du Sperme network. Fertil. Steril. 2017, 107, 580–588. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, C.M.; Chan, P.T.; Hales, B.F.; Robaire, B. Sperm chromatin structure components are differentially repaired in cancer survivors. J. Androl. 2012, 33, 629–636. [Google Scholar] [CrossRef]
- Meistrich, M.L. Risks of Genetic Damage in Offspring Conceived Using Sperm Produced during Chemotherapy or Radiotherapy. Andrology 2019. [Google Scholar] [CrossRef] [Green Version]
- Burrello, N.; Vicari, E.; Vignera, S.L.; Romeo, G.; Campagna, C.; Magro, E.; Giuffrida, D.; D’Agata, R.; Calogero, A.E. Effects of anti-neoplastic treatment on sperm aneuploidy rate in patients with testicular tumor: A longitudinal study. J. Endocrinol. Investig. 2011, 34, 121–125. [Google Scholar] [CrossRef]
- De Mas, P.; Daudin, M.; Vincent, M.-C.; Bourrouillou, G.; Calvas, P.; Mieusset, R.; Bujan, L. Increased aneuploidy in spermatozoa from testicular tumour patients after chemotherapy with cisplatin, etoposide and bleomycin. Hum. Reprod. 2001, 16, 1204–1208. [Google Scholar] [CrossRef] [Green Version]
- Tempest, H.G.; Ko, E.; Chan, P.; Robaire, B.; Rademaker, A.; Martin, R.H. Sperm aneuploidy frequencies analysed before and after chemotherapy in testicular cancer and Hodgkin’s lymphoma patients. Hum. Reprod. 2008, 23, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Martinez, G.; Walschaerts, M.; Le Mitouard, M.; Borye, R.; Thomas, C.; Auger, J.; Berthaut, I.; Brugnon, F.; Daudin, M.; Moinard, N.; et al. Impact of Hodgkin or non-Hodgkin lymphoma and their treatments on sperm aneuploidy: A prospective study by the French CECOS network. Fertil. Steril. 2017, 107, 341–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, F.; Pearson, F.S.; Bishop, J.B.; Wyrobek, A.J. Etoposide induces chromosomal abnormalities in mouse spermatocytes and stem cell spermatogonia. Hum. Reprod. 2006, 21, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, M.; van Casteren, N.; Wildhagen, M.; Romijn, J.; Dohle, G. Sperm DNA integrity in cancer patients before and after cytotoxic treatment. Hum. Reprod. 2010, 25, 1877–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Delft, J.H.M.; van den Ende, A.M.C.; Keizer, H.J.; Ouwerkerk, J.; Baan, R.A. Determination of N7-methylguanine in DNA of white blood cells from cancer patients treated with dacarbazine. Carcinogenesis 1992, 13, 1257–1259. [Google Scholar] [CrossRef]
- Stornetta, A.; Zimmermann, M.; Cimino, G.; Henderson, P.; Sturla, S. DNA Adducts from Anticancer Drugs as Candidate Predictive Markers for Precision Medicine. Chem. Res. Toxicol. 2017, 30, 388–409. [Google Scholar] [CrossRef]
- Poirier, M.C.; Reed, E.; Litterst, C.L.; Katz, D.; Gupta-Burt, S. Persistence of platinum-ammine-DNA adducts in gonads and kidneys of rats and multiple tissues from cancer patients. Cancer Res. 1992, 52, 149–153. [Google Scholar]
- Codrington, A.M.; Hales, B.F.; Robaire, B. Spermiogenic Germ Cell Phase—Specific DNA Damage Following Cyclophosphamide Exposure. J. Androl. 2014, 25, 354–362. [Google Scholar] [CrossRef]
- Manicardi, G.C.; Tombacco, A.; Bizzaro, D.; Bianchi, U.; Bianchi, P.G.; Sakkas, D. DNA strand breaks in ejaculated human spermatozoa: Comparison of susceptibility to the nick translation and terminal transferase assays. Histochem. J. 1998, 30, 33–39. [Google Scholar] [CrossRef]
- Sailer, B.L.; Jost, L.K.; Evenson, D.P. Mammalian sperm DNA susceptibility to in situ denaturation associated with the presence of DNA strand breaks as measured by the terminal deoxynucleotidyl transferase assay. J. Androl. 1995, 16, 80–87. [Google Scholar]
- Maselli, J.; Hales, B.F.; Chan, P.; Robaire, B. Exposure to Bleomycin, Etoposide, and Cis-Platinum Alters Rat Sperm Chromatin Integrity and Sperm Head Protein Profile. Biol. Reprod. 2012, 86, 1–10. [Google Scholar] [CrossRef]
- Maselli, J.; Hales, B.F.; Robaire, B. The Effects of Chemotherapy with Bleomycin, Etoposide, and Cis-Platinum (BEP) on Rat Sperm Chromatin Remodeling, Fecundity and Testicular Gene Expression in the Progeny. Biol. Reprod. 2013, 89, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bagheri-Sereshki, N.; Hales, B.F.; Robaire, B. The Effects of Chemotherapeutic Agents, Bleomycin, Etoposide, and Cisplatin, on Chromatin Remodeling in Male Rat Germ Cells. Biol. Reprod. 2016, 94, 81. [Google Scholar] [CrossRef] [PubMed]
- González-Marín, C.; Gosálvez, J.; Roy, R.; Types, C. Detection and Repair of DNA Fragmentation in Animal and Human Sperm Cells. Int. J. Mol. Sci. 2012, 13, 14026–14052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.J.; Costa, P.; Vaz, B.; Carvalho, F.; Fernandes, S.; Barros, A.; Sousa, M. Abnormal methylation of imprinted genes in human sperm is associated with oligozoospermia. Mol. Hum. Reprod. 2008, 14, 67–74. [Google Scholar] [CrossRef]
- Berthaut, I.; Montjean, D.; Dessolle, L.; Morcel, K.; Deluen, F.; Poirot, C.; Bashamboo, A.; Mcelreavey, K.; Ravel, C. Effect of temozolomide on male gametes: An epigenetic risk to the offspring? J. Assist. Reprod. Genet. 2013, 30, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.; Delbès, G.; Landry, M.; Robaire, B.; Trasler, J.M. Epigenetic Alterations in Sperm DNA Associated with Testicular Cancer Treatment. Toxicol. Sci. 2012, 125, 532–543. [Google Scholar] [CrossRef]
- Sakai, K.; Ideta-Otsuka, M.; Saito, H.; Hiradate, Y.; Hara, K.; Igarashi, K.; Tanemura, K. Effects of doxorubicin on sperm DNA methylation in mouse models of testicular toxicity. Biochem. Biophys. Res. Commun. 2018, 498, 674–679. [Google Scholar] [CrossRef]
- Barton, T.S.; Robaire, B.; Hales, B.F. Epigenetic programming in the preimplantation rat embryo is disrupted by chronic paternal cyclophosphamide exposure. Proc. Natl. Acad. Sci. USA 2005, 102, 7865–7870. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.L.J.; Li, E.; Trasler, J.M. 5-Aza-2’-Deoxycytidine Induces Alterations in Murine Spermatogenesis and Pregnancy Outcome. J. Androl. 2003, 24, 822–830. [Google Scholar] [CrossRef]
- La Salle, S.; Oakes, C.C.; Neaga, O.R.; Bourc’his, D.; Bestor, T.H.; Trasler, J.M. Loss of spermatogonia and wide-spread DNA methylation defects in newborn male mice deficient in DNMT3L. BMC Dev. Biol. 2007, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- van Otterdijk, S.; Michels, K. Transgenerational epigenetic inheritance in mammals: How good is the evidence? FASEB J. 2016, 30, 2457–2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stensheim, H.; Klungsøyr, K.; Skjaerven, R.; Grotmol, T.; Fosså, S. Birth outcomes among offspring of adult cancer survivors: A population-based study. Int. J. Cancer 2013, 33, 2696–2705. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, G.V.; Bielski, C.M.; Samocha, K.; Fromer, M.; Seepo, S.; Gentry, C.; Neale, B.; Garraway, L.A.; Sweeney, C.J.; Taplin, M.-E.; et al. Genetic Effect of Chemotherapy Exposure in Children of Testicular Cancer Survivors. Clin. Cancer Res. 2016, 22, 2183–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcon, L.; Hales, B.F.; Robaire, B. Reversibility of the effects of subchronic exposure to the cancer chemotherapeutics bleomycin, etoposide, and cisplatin on spermatogenesis, fertility, and progeny outcome in the male rat. J. Androl. 2008, 29, 408–417. [Google Scholar] [CrossRef]
- Harrouk, W.; Khatabaksh, S.; Robaire, B.; Hales, B.F. Paternal exposure to cyclophosphamide dysregulates the gene activation program in rat preimplantation embryos. Mol. Reprod. Dev. 2000, 57, 214–223. [Google Scholar] [CrossRef]
- Auroux, M.R.; Dulioust, E.J.B.; Nawar, N.N.; Yacoub, S.G.; Mayaux, M.J.; Schwartz, D.; David, G. Antimitotic drugs in the male rat. Behavioral abnormalities in the second generation. J. Androl. 1988, 9, 153–159. [Google Scholar] [CrossRef]
- Seethalakshmi, L.; Flores, C.; Kinkead, T.; Carboni, A.A.; Malhotra, R.K.; Menon, M. Effects of subchronic treatment with cis-platinum on testicular function, fertility, pregnancy outcome, and progeny. J. Androl. 1992, 13, 65–74. [Google Scholar]
- e Silva, P.V.; da Silva, R.F.; Borges, C.D.S.; Cavariani, M.M.; de Almeida Francia, C.C.D.; Júnior, F.B.; De Grava Kempinas, W. Sexual differentiation and reproductive development of female rat offspring after paternal exposure to the anti-tumor pharmaceutical cisplatin. Reprod. Toxicol. 2016, 60, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Favareto, A.P.A.; de Toledo, F.C.; Kempinas, W.D.G. Paternal treatment with cisplatin impairs reproduction of adult male offspring in rats. Reprod. Toxicol. 2011, 32, 425–433. [Google Scholar] [CrossRef]
- Hales, B.; Crosman, K.; Robaire, B. Increased postimplantation loss and malformations among the F2 progeny of male rats chronically treated with cyclophosphamide. Teratology 1992, 45, 671–678. [Google Scholar] [CrossRef]
- Auroux, M.R.; Dulioust, E.M.; Nawar, N.Y.; Yacoub, S.G.; Kempf, E.H.; Ebel, A.B. Cyclophosphamide in the male rat: Cerebral biochemical changes in progeny. Biomed. Pharmacother. 1990, 44, 519–523. [Google Scholar] [CrossRef]
- Kläver, R.; Sánchez, V.; Damm, O.; Redmann, K.; Lahrmann, E.; Sandhowe-Klaverkamp, R.; Rohde, C.; Wistuba, J.; Ehmcke, J.; Schlatt, S.; et al. Direct but no transgenerational effects of decitabine and vorinostat on male fertility. PLoS ONE 2015, 10, e0117839. [Google Scholar] [CrossRef] [PubMed]
- Ntemou, E.; Alexandri, C.; Lybaert, P.; Goossens, E.; Demeestere, I. Oncofertility: Pharmacological Protection and Immature Testicular Tissue (ITT)-Based Strategies for Prepubertal and Adolescent Male Cancer Patients. Int. J. Mol. Sci. 2019, 20, 5223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Michele, F.; Vermeulen, M.; Wyns, C. Fertility restoration with spermatogonial stem cells. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 424–431. [Google Scholar] [CrossRef]
- Zarandi, N.P.; Galdon, G.; Kogan, S.; Atala, A.; Sadri-Ardekani, H. Cryostorage of immature and mature human testis tissue to preserve spermatogonial stem cells (SSCs): A systematic review of current experiences toward clinical applications. Stem Cells Cloning 2018, 11, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Perrard, M.-H.; Sereni, N.; Schluth-Bolard, C.; Blondet, A.; d’Estaing, S.G.; Plotton, I.; Morel-Journel, N.; Lejeune, H.; David, L.; Durand, P. Complete Human and Rat Ex Vivo Spermatogenesis from Fresh or Frozen Testicular Tissue. Biol. Reprod. 2016, 95, 89. [Google Scholar] [CrossRef]
- Abu Elhija, M.; Lunenfeld, E.; Schlatt, S.; Huleihel, M. Differentiation of murine male germ cells to spermatozoa in a soft agar culture system. Asian J. Androl. 2012, 14, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In vitro production of functional sperm in cultured neonatal mouse testes. Nature 2011, 471, 504–507. [Google Scholar] [CrossRef]
- Oblette, A.; Rives, N.; Dumont, L.; Rives, A.; Verhaeghe, F.; Jumeau, F.; Rondanino, C. Assessment of sperm nuclear quality after in vitro maturation of fresh or frozen/thawed mouse pre-pubertal testes. Mol. Hum. Reprod. 2017, 23, 674–684. [Google Scholar] [CrossRef]
- AbuMadighem, A.; Solomon, R.; Stepanovsky, A.; Kapelushnik, J.; Shi, Q.; Meese, E.; Lunenfeld, E.; Huleihel, M. Development of Spermatogenesis In Vitro in Three-Dimensional Culture from Spermatogonial Cells of Busulfan-Treated Immature Mice. Int. J. Mol. Sci. 2018, 19, 3804. [Google Scholar] [CrossRef] [Green Version]
- Yokonishi, T.; Sato, T.; Komeya, M.; Katagiri, K.; Kubota, Y.; Nakabayashi, K.; Hata, K.; Inoue, K.; Ogonuki, N.; Ogura, A.; et al. Offspring production with sperm grown in vitro from cryopreserved testis tissues. Nat. Commun. 2014, 5, 4320. [Google Scholar] [CrossRef] [PubMed]
- Abofoul-Azab, M.; AbuMadighem, A.; Lunenfeld, E.; Kapelushnik, J.; Shi, Q.; Pinkas, H.; Huleihel, M. Development of Postmeiotic Cells In Vitro from Spermatogonial Cells of Prepubertal Cancer Patients. Stem Cells Dev. 2018, 27, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- de Michele, F.; Poels, J.; Vermeulen, M.; Ambroise, J.; Gruson, D.; Guiot, Y.; Wyns, C. Haploid Germ Cells Generated in Organotypic Culture of Testicular Tissue From Prepubertal Boys. Front. Physiol. 2018, 9, 1413. [Google Scholar] [CrossRef] [PubMed]
- Medrano, J.V.; Vilanova-Pérez, T.; Fornés-Ferrer, V.; Navarro-Gomezlechon, A.; Martínez-Triguero, M.L.; García, S.; Gómez-Chacón, J.; Povo, I.; Pellicer, A.; Andrés, M.M.; et al. Influence of temperature, serum, and gonadotropin supplementation in short- and long-term organotypic culture of human immature testicular tissue. Fertil. Steril. 2018, 110, 1045–1057.e3. [Google Scholar] [CrossRef] [Green Version]
- Portela, J.M.D.; de Winter-Korver, C.M.; van Daalen, S.K.M.; Meißner, A.; de Melker, A.A.; Repping, S.; van Pelt, A.M.M. Assessment of fresh and cryopreserved testicular tissues from (pre)pubertal boys during organ culture as a strategy for in vitro spermatogenesis. Hum. Reprod. 2019, 34, 2443–2455. [Google Scholar] [CrossRef]
- Alexandri, C.; Stamatopoulos, B.; Rothé, F.; Bareche, Y.; Devos, M.; Demeestere, I. MicroRNA profiling and identification of let-7a as a target to prevent chemotherapy-induced primordial follicles apoptosis in mouse ovaries. Sci. Rep. 2019, 9, 9636. [Google Scholar] [CrossRef] [Green Version]
- Carmely, A.; Meirow, D.; Peretz, A.; Albeck, M.; Bartoov, B.; Sredni, B. Protective effect of the immunomodulator AS101 against cyclophosphamide-induced testicular damage in mice. Hum. Reprod. 2009, 24, 1322–1329. [Google Scholar] [CrossRef] [Green Version]
- Ilbey, Y.O.; Ozbek, E.; Simsek, A.; Otunctemur, A.; Cekmen, M.; Somay, A. Potential chemoprotective effect of melatonin in cyclophosphamide- and cisplatin-induced testicular damage in rats. Fertil. Steril. 2009, 92, 1124–1132. [Google Scholar] [CrossRef]
- Benavides-Garcia, R.; Joachim, R.; Pina, N.A.; Mutoji, K.N.; Reilly, M.A.; Hermann, B.P. Granulocyte colony-stimulating factor prevents loss of spermatogenesis after sterilizing busulfan chemotherapy. Fertil. Steril. 2015, 103, 270–280. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Agents (Cumulative Dose for Effect) | Class of Anticancer Drugs | Risk on Fertility after Adulthood Exposure [12] | Risk on Fertility after Childhood Exposure [13] |
|---|---|---|---|
| Chlorambucil (1.4 g/m2) | Alkylating agent | High | |
| Cyclophosphamide (19 g/m2) | High | High (7.5 g/m2) | |
| Procarbazine (4 g/m2) | (Prolonged azoospermia) | High | |
| Melphalan (140 mg/m2) | High | ||
| Cisplatin (500 mg/m2) | High | ||
| Busulfan (600 mg/kg) | Alkylating agent | High | |
| Ifosfamide (42 g/m2) | Moderate | High (4 g/m2) | |
| Carmustin (300 mg/m2) | (Likelihood of azoospermia, but always given with other sterilizing agents) | Low | |
| Dactinomycin | DNA Intercalating | Low | |
| Carboplatin (2 g/m2) | Alkylating agent | Moderate | |
| Thiotepa (400 mg/m2) | Moderate | ||
| Doxorubicin (770 mg/m2) | DNA Intercalating | Moderate | |
| Cytarabine (1 g/m2) | Antimetabolite | Moderate | |
| Vinblastine (50 g/m2) | Spindle poison | Low | |
| Vincristine (8 g/m2) | Low | ||
| Low | |||
| Dacarbazine | Alkylating agent | (Temporary reduction in sperm counts) | Moderate |
| Daunorubicin | DNA Intercalating | Moderate | |
| Mitoxantrone | Moderate | ||
| Bleomycin | DNA strand breaks inducer | Low | |
| Etoposide | Topoisomerase II inhibitor | Low | |
| Fludarabine | Antimetabolite | Unknown | |
| Fluorouracil | Low | ||
| Mercaptopurine | Low | ||
| Methotrexate | Low | ||
| Thioguanine | Unknown |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delessard, M.; Saulnier, J.; Rives, A.; Dumont, L.; Rondanino, C.; Rives, N. Exposure to Chemotherapy During Childhood or Adulthood and Consequences on Spermatogenesis and Male Fertility. Int. J. Mol. Sci. 2020, 21, 1454. https://doi.org/10.3390/ijms21041454
Delessard M, Saulnier J, Rives A, Dumont L, Rondanino C, Rives N. Exposure to Chemotherapy During Childhood or Adulthood and Consequences on Spermatogenesis and Male Fertility. International Journal of Molecular Sciences. 2020; 21(4):1454. https://doi.org/10.3390/ijms21041454
Chicago/Turabian StyleDelessard, Marion, Justine Saulnier, Aurélie Rives, Ludovic Dumont, Christine Rondanino, and Nathalie Rives. 2020. "Exposure to Chemotherapy During Childhood or Adulthood and Consequences on Spermatogenesis and Male Fertility" International Journal of Molecular Sciences 21, no. 4: 1454. https://doi.org/10.3390/ijms21041454