Inhibition of Proinflammatory Enzymes and Attenuation of IL-6 in LPS-Challenged RAW 264.7 Macrophages Substantiates the Ethnomedicinal Use of the Herbal Drug Homalium bhamoense Cubitt & W.W.Sm

 , ,
, , 
 ,
, 
Abstract
1. Introduction
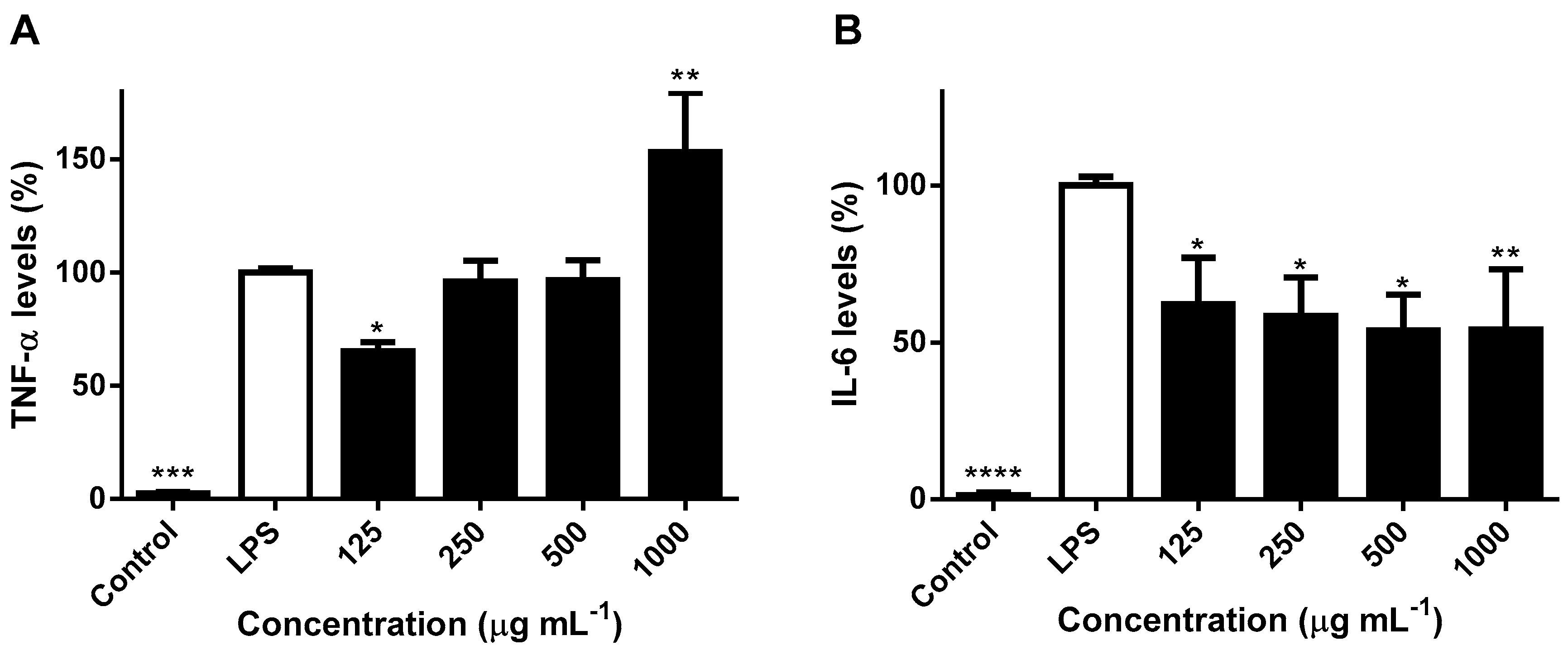
2. Results and Discussion
2.1. HPLC-DAD Characterization of the Phenolic Profile of H. bhamoense Trunk Bark Extract
2.2. H. bhamoense Trunk Bark Extract Significantly Inhibits Hyaluronidase and 5-Lipoxygenase
2.3. Effects on NO Levels and Inflammatory Cytokines in LPS-Stimulated RAW 264.7 Macrophages
2.4. Effects in the Viability of Human Keratinocyte HaCaT Cells
3. Materials and Methods
3.1. General Chemicals and Materials
3.2. Collection of Plant Material and Preparation of Crude Extract
3.3. HPLC-DAD Qualitative and Quantitative Analysis
3.4. Enzymatic Assays
3.4.1. Inhibition of Hyaluronidase Activity
3.4.2. Inhibition of 5-Lipoxygenase Activity
3.5. Nitric Oxide Radical Scavening Activity
3.6. Interference with LPS-Induced NO Production by RAW 264.7 Cells
3.6.1. Cell Viability
3.6.2. Nitrite Assay
3.7. Interference with LPS-Induced TNF-α and IL-6 Production by RAW 264.7 Cells
3.7.1. Enzyme-Linked Immunosorbent Assay (ELISA)
3.7.2. Total Protein Quantification
3.8. Interference with HaCaT Cells’ Viability
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-LOX | 5-Lipoxygenase |
| ATCC® | American Type Culture Collection |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| iNOS | Inducible nitric oxide synthase |
| IL-6 | Interleukin-6 |
| LPS | Lipopolysaccharide |
| NF-κB | Nuclear factor kappa B |
| NO | Nitric oxide |
| TNF-α | Tumor necrosis factor α |
References
- Mahapatra, A.K.; Pani, S.S.; Sahoo, A.K. Free radical-scavenging activities of Homalium species—An endangered medicinal plant of Eastern Ghats of India. Nat. Prod. Res. 2015, 29, 2112–2116. [Google Scholar] [CrossRef] [PubMed]
- Prescott, T.A.K.; Homot, P.; Lundy, F.T.; Fang, R.; Patrick, S.; Cámara-Leret, R.; Kiapranis, R. Tropical ulcer plant treatments used by Papua New Guinea’s Apsokok nomads. J. Ethnopharmacol. 2017, 205, 240–245. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sahoo, A.K.; Dash, U.C.; Kanhar, S.; Mahapatra, A.K. In vitro biological assessment of Homalium zeylanicum and isolation of lucidenic acid A triterpenoid. Toxicol. Rep. 2017, 4, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Okokon, J.E.; Okokon, P.J.; Dar Farooq, A.; Choudhary, M.I. Anti-inflammatory and antinociceptive activities of Homalium letestui. Pharm. Biol. 2013, 51, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, S.; Sai Kumar, P.; Vinod, K.R.; Banji, D.; Kumar, K. Plants as potent anti-diabetic and wound healing agents—A review. Hygeia J. D. Med. 2011, 3, 11–19. [Google Scholar]
- Prescott, T.A.K.; Briggs, M.; Kiapranis, R.; Simmonds, M.S.J. Medicinal plants of Papua New Guinea’s Miu speaking population and a focus on their use of plant–slaked lime mixtures. J. Ethnopharmacol. 2015, 174, 217–223. [Google Scholar] [CrossRef]
- Sleumer, H. The Flacourtiaceae of Thailand. Blumea 1985, 30, 217–250. [Google Scholar]
- Wiart, C. Medicinal Plants Classified in the Family Flacourtiaceae. In Medicinal Plants of Asia and the Pacific; Wiart, C., Ed.; CRC Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 97–98. [Google Scholar]
- Maytin, E.V. Hyaluronan: More than just a wrinkle filler. Glycobiology 2016, 26, 553–559. [Google Scholar] [CrossRef]
- Guimarães, F.R.; Sales-Campos, H.; Nardini, V.; da Costa, T.A.; Fonseca, M.T.C.; Júnior, V.R.; Sorgi, C.A.; da Silva, J.S.; Chica, J.E.L.; Faccioli, L.H.; et al. The inhibition of 5-lipoxygenase (5-LO) products leukotriene B4 (LTB4) and cysteinyl leukotrienes (cysLTs) modulated the inflammatory response and improves cutaneous wound healing. Clin. Immunol. 2018, 190, 74–83. [Google Scholar] [CrossRef]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef]
- Liang, J.; Jiang, D.; Noble, P.W. Hyaluronan as a therapeutic target in human diseases. Adv. Drug Deliv. Rev. 2016, 97, 186–203. [Google Scholar] [CrossRef] [PubMed]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune regulation of skin wound healing: Mechanisms and novel therapeutic targets. Adv. Wound Care (New Rochelle) 2018, 7, 209–231. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.F.; Bártolo, P.J. Traditional therapies for skin wound healing. Adv. Wound Care (New Rochelle) 2016, 5, 208–229. [Google Scholar] [CrossRef] [PubMed]
- Manca, M.L.; Zaru, M.; Bacchetta, G.; Biggio, T.; Cappai, N.; Cabras, A.; Falchi, A.M.; Manconi, M.; Fadda, A.M. A new technological approach to improve the efficacy of a traditional herbal medicinal product in wound healing. Ind. Crop. Prod. 2015, 63, 71–78. [Google Scholar] [CrossRef]
- Castangia, I.; Caddeo, C.; Manca, M.L.; Casu, L.; Latorre, A.C.; Díez-Sales, O.; Ruiz-Saurí, A.; Bacchetta, G.; Fadda, A.M.; Manconi, M. Delivery of liquorice extract by liposomes and hyalurosomes to protect the skin against oxidative stress injuries. Carbohydr. Polym. 2015, 134, 657–663. [Google Scholar] [CrossRef]
- Manca, M.L.; Castangia, I.; Zaru, M.; Nácher, A.; Valenti, D.; Fernàndez-Busquets, X.; Fadda, A.M.; Manconi, M. Development of curcumin loaded sodium hyaluronate immobilized vesicles (hyalurosomes) and their potential on skin inflammation and wound restoring. Biomaterials 2015, 71, 100–109. [Google Scholar] [CrossRef]
- Ekabo, O.A.; Farnsworth, N.R.; Santisuk, T.; Reutrakul, V. Phenolic, iridoid and ionyl glycosides from Homalium ceylanicum. Phytochemistry 1993, 32, 747–754. [Google Scholar] [CrossRef]
- Liu, L.; Guo, Z.; Chai, X.; Zhao, M.; Lu, Y.; Tu, P. Phenolic glycosides from the stems of Homalium ceylanicum (Gardner) Bentham (Flacourtiaceae/Salicaceae sensu lato). Biochem. Syst. Ecol. 2013, 46, 55–58. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Zheng, C.J.; Bai, M.; Li, X.B.; Song, X.P.; Han, C.R. Cytotoxic constituents of the twigs of Homalium stenophyllum. Chem. Nat. Compd. 2017, 53, 362–364. [Google Scholar] [CrossRef]
- Wu, S.Y.; Fu, Y.H.; Zhou, Q.; Bai, M.; Chen, G.Y.; Han, C.R.; Song, X.P. A new dihydrochalcone glycoside from the stems of Homalium stenophyllum. Nat. Prod. Res. 2018, 32, 953–958. [Google Scholar] [CrossRef]
- Ferreres, F.; Grosso, C.; Gil-Izquierdo, A.; Valentão, P.; Andrade, P.B. Ellagic acid and derivatives from Cochlospermum angolensis Welw. extracts: HPLC-DAD-ESI/MSn profiling, quantification and in vitro anti-depressant, anti-cholinesterase and anti-oxidant activities. Phytochem. Anal. 2013, 24, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Maibach, H.I. Hyaluronan in skin: Aspects of aging and its pharmacologic modulation. Clin. Dermatol. 2008, 26, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Jedrzejas, M.J. The hyaluronidases: Their genomics, structures, and mechanisms of action. Chem. Rev. 2008, 106, 818–839. [Google Scholar] [CrossRef] [PubMed]
- Kuppusami, U.R.; Das, N.P. Inhibitory effects of flavonoids on several venom hyaluronidases. Experientia 1991, 47, 1196–1200. [Google Scholar] [CrossRef]
- Sgariglia, M.A.; Soberón, J.R.; Cabanes, A.P.; Sampietro, D.A.; Vattuone, M.A. Anti-inflammatory properties of phenolic lactones isolated from Caesalpinia paraguariensis stem bark. J. Ethnopharmacol. 2013, 147, 63–73. [Google Scholar] [CrossRef]
- Cho, K.J.; Seo, J.M.; Kim, J.H. Bioactive lipoxygenase metabolites stimulation of NADPH oxidases and reactive oxygens species. Mol. Cells 2011, 32, 1–5. [Google Scholar] [CrossRef]
- Brogliato, A.R.; Moor, A.N.; Kesl, S.L.; Guilherme, R.F.; Georgii, J.L.; Peters-Golden, M.; Canetti, C.; Gould, L.J.; Benjamim, C.F. Critical role of 5-lipoxygenase and heme oxygenase-1 in wound healing. J. Investig. Dermatol. 2014, 134, 1436–1445. [Google Scholar] [CrossRef]
- Kendall, A.C.; Nicolaou, A. Bioactive lipid mediators in skin inflammation and immunity. Prog. Lipid Res. 2013, 52, 141–164. [Google Scholar] [CrossRef]
- El-Shitany, N.A.; El-Bastawissy, E.A.; El-Desoky, K. Ellagic acid protects against carrageenan-induced acute inflammation through inhibition of nuclear factor kappa B, inducible cyclooxygenase and proinflammatory cytokines and enhancement of interleukin-10 via an antioxidant mechanism. Int. Immunopharmacol. 2014, 19, 290–299. [Google Scholar] [CrossRef]
- Kawakami, Y.; Hosokawa, T.; Morinaka, T.; Irino, S.; Hirano, S.; Kobayashi, H.; Yoshioka, A.; Suzuki-Yamamoto, T.; Yokoro, M.; Kimoto, M.; et al. Antiatherogenic effect of guava leaf extracts inhibiting leucocyte-type 12-lipoxygenase activity. Food Chem. 2012, 131, 1069–1075. [Google Scholar] [CrossRef]
- McKee, C.M.; Lowenstein, C.J.; Horton, M.R.; Wu, J.; Bao, C.; Chin, B.Y.; Choi, A.M.; Noble, P.W. Hyaluronan fragments induce nitric-oxide synthase in murine macrophages through a nuclear factor kappaB-dependent mechanism. J. Biol. Chem. 1997, 272, 8013–8018. [Google Scholar] [CrossRef] [PubMed]
- Trachootham, D.; Lu, W.; Ogasawara, M.A.; Nilsa, R.D.; Huang, P. Redox regulation of cell survival. Antioxid. Redox Signal. 2008, 10, 1343–1374. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, J.; Zhang, X.; Wang, L.; Zhang, W. Pomegranate peel polyphenols inhibits inflammation in LPS-induced RAW 264.7 macrophages via the suppression of MAPKs activation. J. Funct. Foods 2018, 43, 62–69. [Google Scholar] [CrossRef]
- Verotta, L.; Panzella, L.; Antenucci, S.; Calvenzani, V.; Tomay, F.; Petroni, K.; Caneva, E.; Napolitano, A. Fermented pomegranate wastes as sustainable source of ellagic acid: Antioxidant properties, anti-inflammatory action, and controlled release under simulated digestion conditions. Food Chem. 2018, 246, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.S.; Jeong, S.J.; Yoo, S.R.; Lee, N.R.; Shin, H.K. Quantitative analysis and in vitro anti-inflammatory effects of gallic acid, ellagic acid, and quercetin from Radix Sanguisorbae. Pharmacogn. Mag. 2016, 12, 104–108. [Google Scholar] [CrossRef]
- Landen, N.X.; Li, D.; Stahle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef]
- Schwentker, A.; Vodovotz, Y.; Weller, R.; Billiar, T.R. Nitric oxide and wound repair: Role of cytokines? Nitric Oxide 2002, 7, 1–10. [Google Scholar] [CrossRef]
- Kalliolias, G.D.; Ivashkiv, L.B. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef]
- Eigler, A.; Moeller, J.; Endres, S. Exogenous and endogenous nitric oxide attenuates tumor necrosis factor synthesis in the murine macrophage cell line RAW264.7. J. Immunol. 1995, 154, 4048–4054. [Google Scholar]
- Marcinkiewicz, J.; Grabowska, A.; Chain, B. Nitric oxide up-regulates the release of inflammatory mediators by mouse macrophages. Eur. J. Immunol. 1995, 25, 947–951. [Google Scholar] [CrossRef]
- Deakin, A.M.; Payne, A.N.; Whittle, B.J.R.; Moncada, S. The modulation of IL-6 and TNF-α release by nitric oxide following stimulation of J774 cells with LPS and IFN-γ. Cytokine 1995, 7, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Grellner, W.; Georg, T.; Wilske, J. Quantitative analysis of proinflammatory cytokines (IL-1β, IL-6, TNF-α) in human skin wounds. Forensic Sci. Int. 2000, 113, 251–264. [Google Scholar] [CrossRef]
- Guan, S.; Zheng, Y.; Yu, X.; Han, B.; Lu, J. Ellagic acid protects against LPS-induced acute lung injury through inhibition of nuclear factor kappa B, proinflammatory cytokines and enhancement of interleukin-10. Food Agr. Immunol. 2017, 28, 1347–1361. [Google Scholar] [CrossRef]
- Barker, J.; Griffiths, C.; Mitra, R.; Dixit, V.; Nickoloff, B.; Griffiths, C. Keratinocytes as initiators of inflammation. Lancet 1991, 337, 211–214. [Google Scholar] [CrossRef]
- Wiegand, C.; Hipler, U.C. Evaluation of biocompatibility and cytotoxicity using keratinocyte and fibroblast cultures. Skin Pharmacol. Physiol. 2009, 22, 74–82. [Google Scholar] [CrossRef]
- Pinho, B.R.; Sousa, C.; Valentão, P.; Oliveira, J.M.A.; Andrade, P.B. Modulation of basophils’ degranulation and allergy-related enzymes by monomeric and dimeric naphthoquinones. PLoS ONE 2014, 9, e90122. [Google Scholar] [CrossRef]
- Macedo, T.; Ribeiro, V.; Oliveira, A.P.; Pereira, D.M.; Fernandes, F.; Gomes, N.G.M.; Araújo, L.; Valentão, P.; Andrade, P.B. Anti-inflammatory properties of Xylopia aethiopica leaves: Interference with pro-inflammatory cytokines in THP-1-derived macrophages and flavonoid profiling. J. Ethnopharmacol. 2020, 248, 112312. [Google Scholar] [CrossRef]
- Ferreres, F.; Duangsrisai, S.; Gomes, N.G.M.; Suksungworn, R.; Pereira, D.M.; Gil-Izquierdo, A.; Valentão, P.; Choowongkomon, K.; Andrade, P.B. Anti-inflammatory properties of the stem bark from the herbal drug Vitex peduncularis Wall. ex Schauer and characterization of its polyphenolic profile. Food Chem. Toxicol. 2017, 106, 8–16. [Google Scholar] [CrossRef]
- Gomes, N.G.M.; Oliveira, A.P.; Cunha, D.; Pereira, D.M.; Valentão, P.; Pinto, E.; Araújo, L.; Andrade, P.B. Flavonoid composition of Salacia senegalensis (Lam.) DC. leaves, evaluation of antidermatophytic effects, and potential amelioration of the associated inflammatory response. Molecules 2019, 24, 2530. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Rt | mg Kg−1 (Dry Extract) |
|---|---|---|
| 1 Ellagic acid | 34.6 | 2363.30 ± 53.89 |
| 2 Methyl ellagic acid pentoside | 42.4 | 1590.60 ± 25.51 |
| 3 Methyl ellagic acid deoxyhexoside | 47.0 | 551.52 ± 4.38 |
| 4 Methyl ellagic acid | 47.5 | 1029.36 ± 10.45 |
| Total | 5534.78 ± 94.23 | |
| Regression Equation | ||||||
|---|---|---|---|---|---|---|
| Slope (σ) | Intercept (b) | R2 (n = 3) | Linearity Range (µg mL−1) | LOD (µg mL−1) | LOQ (µg mL−1) | |
| Ellagic acid | 164.35 | −3443 | 0.9997 | 26.88–430 | 3.55 | 10.74 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suksungworn, R.; Andrade, P.B.; Oliveira, A.P.; Valentão, P.; Duangsrisai, S.; Gomes, N.G.M. Inhibition of Proinflammatory Enzymes and Attenuation of IL-6 in LPS-Challenged RAW 264.7 Macrophages Substantiates the Ethnomedicinal Use of the Herbal Drug Homalium bhamoense Cubitt & W.W.Sm. Int. J. Mol. Sci. 2020, 21, 2421. https://doi.org/10.3390/ijms21072421
Suksungworn R, Andrade PB, Oliveira AP, Valentão P, Duangsrisai S, Gomes NGM. Inhibition of Proinflammatory Enzymes and Attenuation of IL-6 in LPS-Challenged RAW 264.7 Macrophages Substantiates the Ethnomedicinal Use of the Herbal Drug Homalium bhamoense Cubitt & W.W.Sm. International Journal of Molecular Sciences. 2020; 21(7):2421. https://doi.org/10.3390/ijms21072421
Chicago/Turabian StyleSuksungworn, Rungcharn, Paula B. Andrade, Andreia P. Oliveira, Patrícia Valentão, Sutsawat Duangsrisai, and Nelson G. M. Gomes. 2020. "Inhibition of Proinflammatory Enzymes and Attenuation of IL-6 in LPS-Challenged RAW 264.7 Macrophages Substantiates the Ethnomedicinal Use of the Herbal Drug Homalium bhamoense Cubitt & W.W.Sm" International Journal of Molecular Sciences 21, no. 7: 2421. https://doi.org/10.3390/ijms21072421
APA StyleSuksungworn, R., Andrade, P. B., Oliveira, A. P., Valentão, P., Duangsrisai, S., & Gomes, N. G. M. (2020). Inhibition of Proinflammatory Enzymes and Attenuation of IL-6 in LPS-Challenged RAW 264.7 Macrophages Substantiates the Ethnomedicinal Use of the Herbal Drug Homalium bhamoense Cubitt & W.W.Sm. International Journal of Molecular Sciences, 21(7), 2421. https://doi.org/10.3390/ijms21072421