Residues in Transmembrane Segments of the P2X4 Receptor Contribute to Channel Function and Ethanol Sensitivity


 ,
, 
Abstract
1. Introduction
2. Results
2.1. TM1 Alanine Scan Revealed Residues That Are Important for Receptor Function
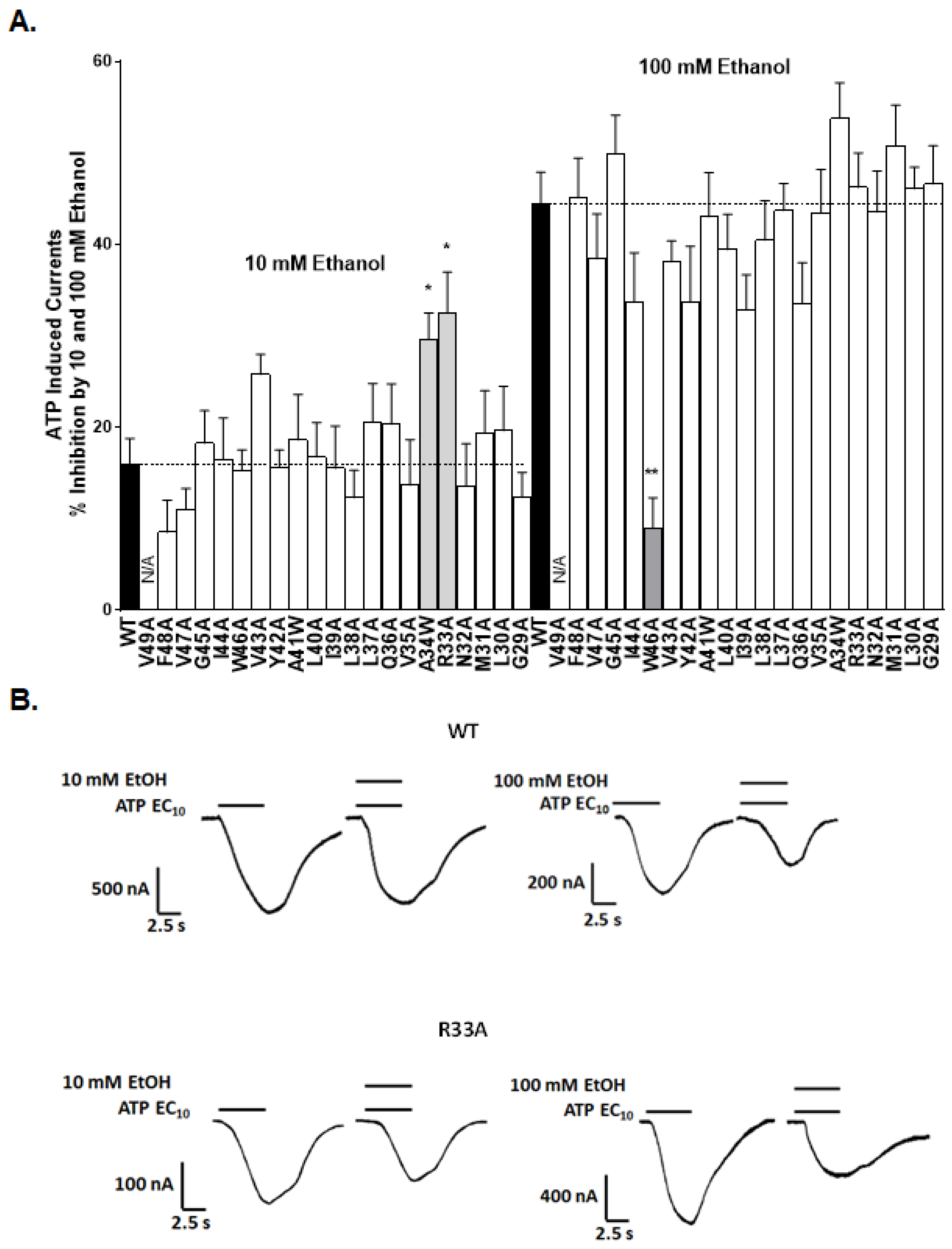
2.2. Alanine Scan of the TM1 Revealed Sites for Ethanol Action
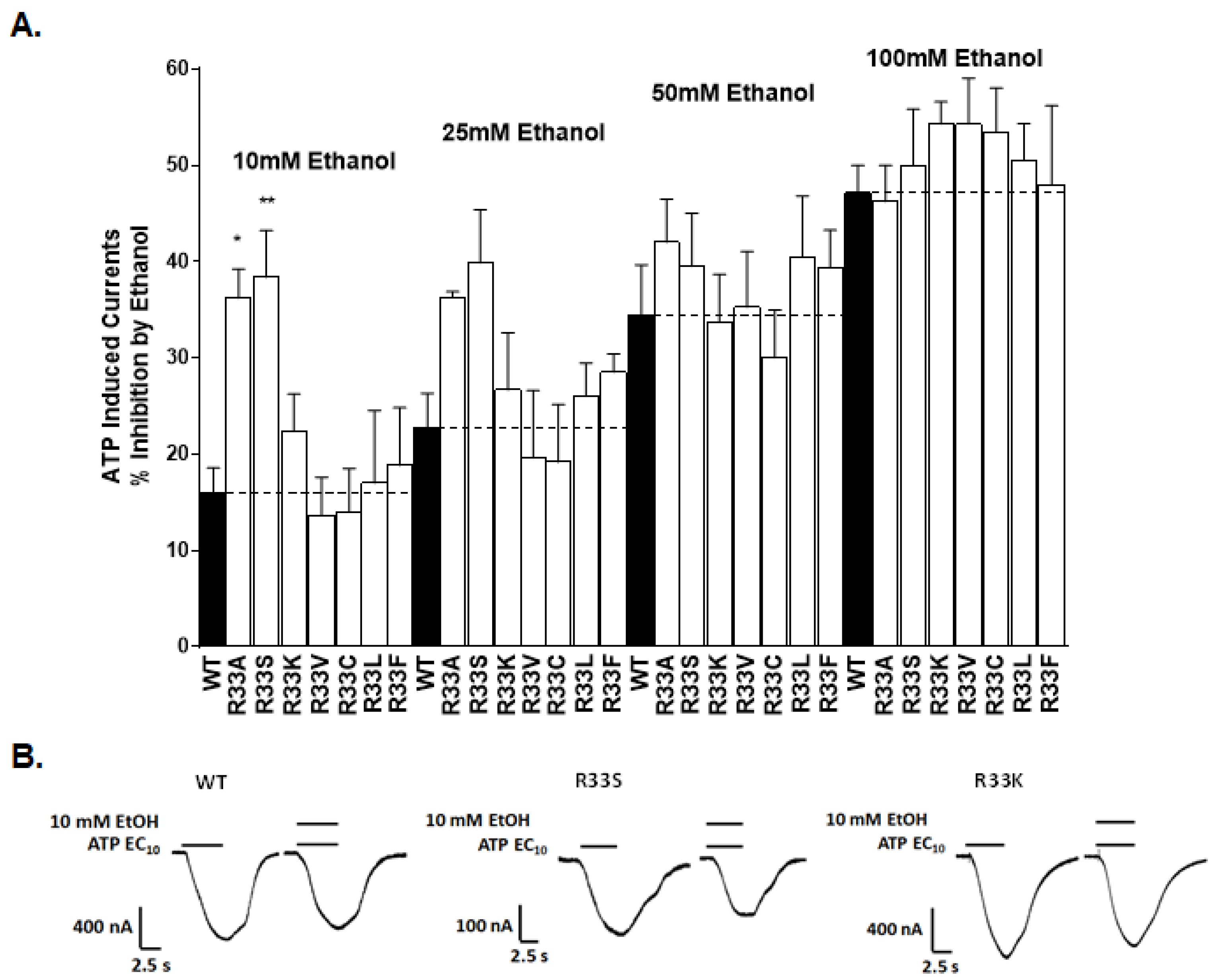
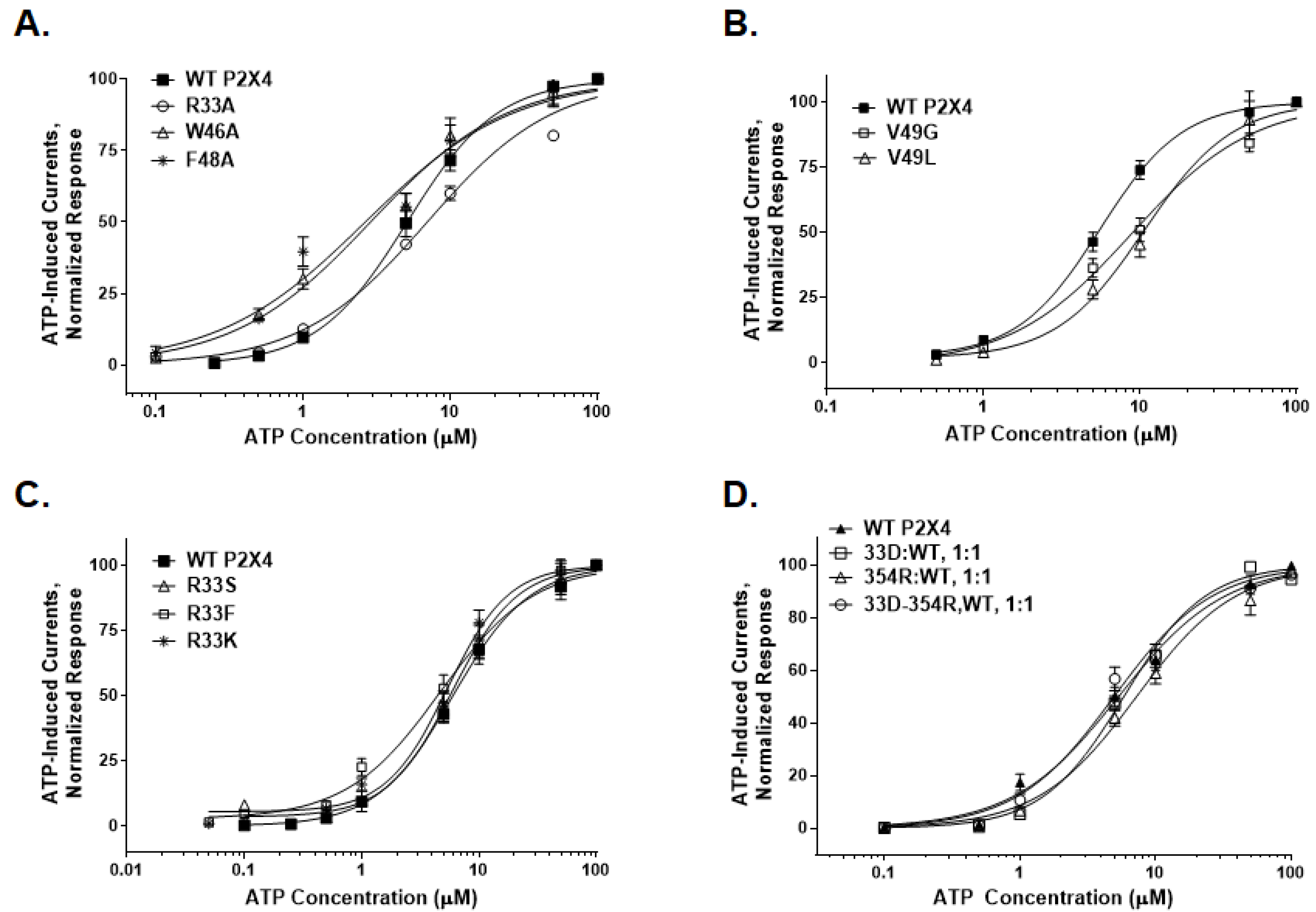
2.3. Physical-Chemical Properties of Residues at Position 33 Determine Receptor Function and Ethanol Sensitivity
2.3.1. Receptor Function
2.3.2. Ethanol Response
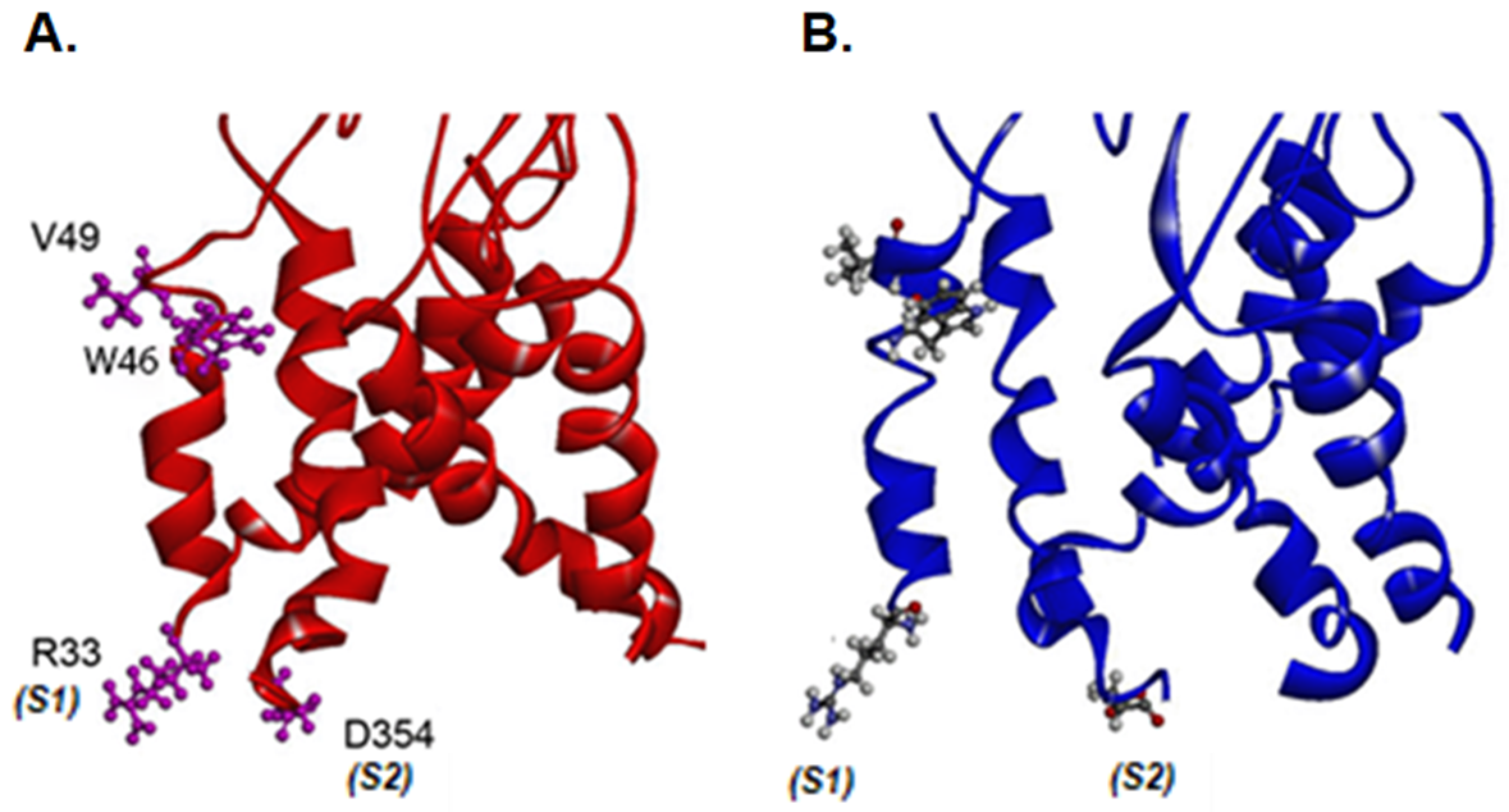
2.4. Residues That Are Important for Agonist Sensitivity and Ethanol Effects Identified in Homology Models
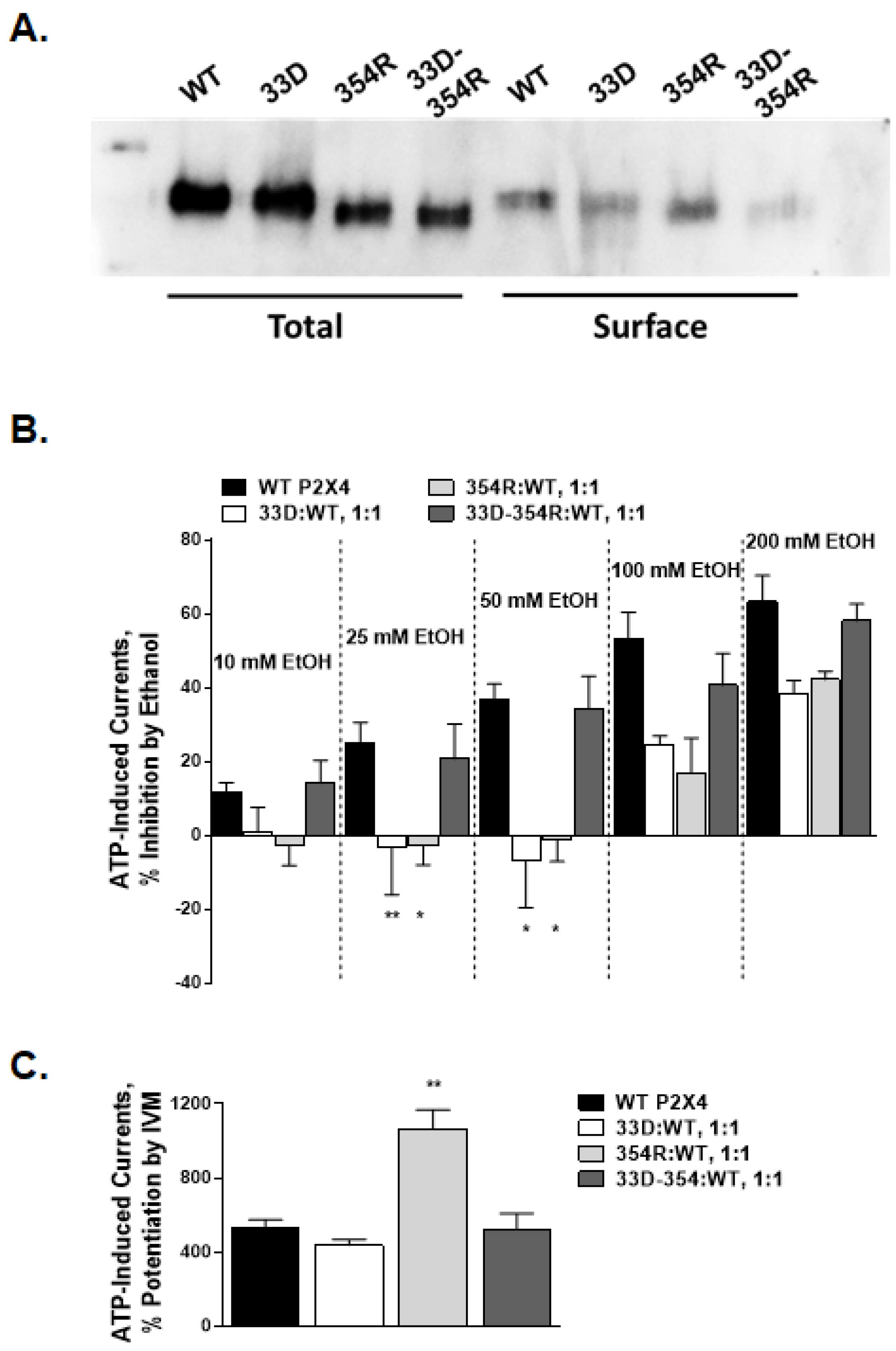
2.5. Arginine 33 and Aspartic Acid 354 Interactions
3. Discussion
3.1. Function of Valine at Position 49 in the upper Portion of the TM1 Segment
3.2. Arginine at Position 33 in the TM1 Segment in Ethanol Action
3.3. Interaction between TM1 Arginine at Position 33 and TM2 Aspartic Acid at Position 354 in Channel Functioning
4. Materials and Methods
4.1. Materials
4.2. Isolation of Xenopus Laevis Oocytes and cRNA Injections
4.3. Site-Directed Mutagenesis and cRNA Synthesis
4.4. Whole-Cell Voltage Clamp Recordings
4.5. Experimental Procedures
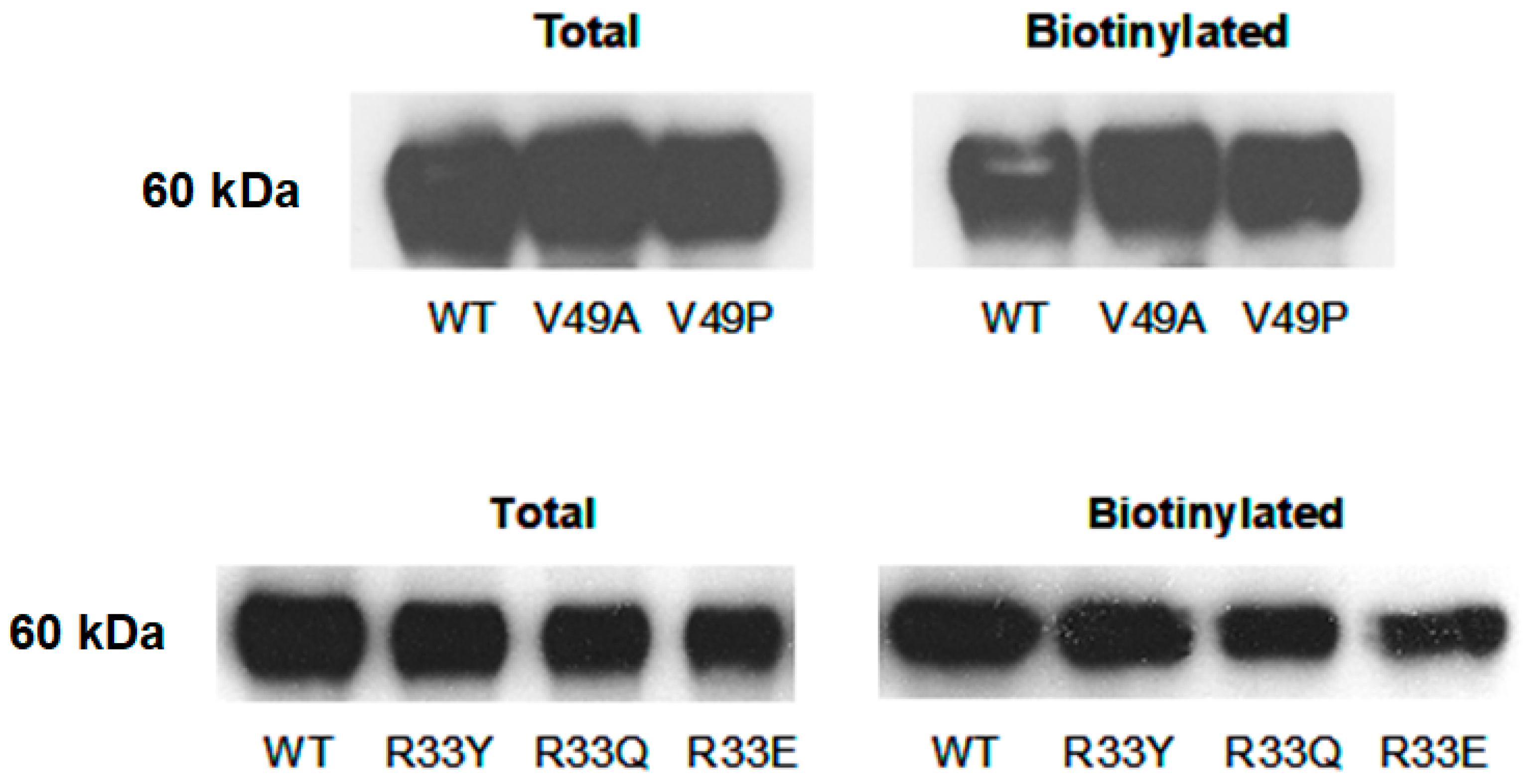
4.6. Cell Surface Biotinylation and Immunoblotting
4.7. Homology Modeling
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AUD | alcohol use disorder |
| ATP | adenosine triphosphate |
| EC | effective concentration |
| P2 × 4R | purinergic P2X4receptor |
| p2rx4 | gene encoding P2 × 4R |
| TM | transmembrane |
| WT | wild-type |
| Imax | maximum inducible current |
| IVM | ivermectin |
Appendix A

References
- Sacks, J.J.; Gonzales, K.R.; Bouchery, E.E.; Tomedi, L.E.; Brewer, R.D. 2010 national and state costs of excessive alcohol consumption. Am. J. Prev. Med. 2015, 49, e73–e79. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Status Report on Alcohol and Health; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Franklin, K.M.; Asatryan, L.; Jakowec, M.W.; Trudell, J.R.; Bell, R.L.; Davies, D.L. P2X4 receptors (P2X4Rs) represent a novel target for the development of drugs to prevent and/or treat alcohol use disorders. Front. Neurosci. 2014, 8, 176. [Google Scholar] [CrossRef] [PubMed]
- Buell, G.; Collo, G.; Rassendren, F. P2X receptors: An emerging channel family. Eur. J. Neurosci. 1996, 8, 2221–2228. [Google Scholar] [CrossRef] [PubMed]
- Soto, F.; Garcia-Guzman, M.; Gomez-Hernandez, J.M.; Hollmann, M.; Karschin, C.; Stuhmer, W. P2X4: An ATP-activated ionotropic receptor clonned from rat brain. Proc. Natl. Acad. Sci. USA 1996, 93, 3684–3688. [Google Scholar] [CrossRef]
- Gonzales, R.A.; Job, M.O.; Doyon, W.M. The role of mesolimbic dopamine in the development and maintenance of ethanol reinforcement. Pharmacol. Ther. 2004, 103, 121–146. [Google Scholar] [CrossRef]
- Kimpel, M.W.; Strother, W.N.; McClintick, J.N.; Carr, L.G.; Liang, T.; Edenberg, H.J.; McBride, W.J. Functional gene expression differences between inbred alcohol-preferring and -non-preferring rats in five brain regions. Alcohol 2007, 41, 95–132. [Google Scholar] [CrossRef]
- Tabakoff, B.; Saba, L.; Printz, M.; Flodman, P.; Hodgkinson, C.; Goldman, D.; Koob, G.; Richardson, H.; Kechris, K.; Bell, R.L.; et al. Genetical genomic determinants of alcohol consumption in rats and humans. BMC Biol. 2009, 7, 70. [Google Scholar] [CrossRef]
- Wyatt, L.R.; Godar, S.C.; Khoja, S.; Jakowec, M.W.; Alkana, R.L.; Bortolato, M.; Davies, D.L. Sociocommunicative and sensorimotor impairments in male P2X4-deficient mice. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2013, 38, 1993–2002. [Google Scholar] [CrossRef]
- Xiong, K.; Li, C.; Weight, F.F. Inhibition by ethanol of rat P2X4 receptors expressed in Xenopus oocytes. Br. J. Pharmacol. 2000, 130, 1394–1398. [Google Scholar] [CrossRef]
- Davies, D.L.; Kochegarov, A.A.; Kuo, S.T.; Kulkarni, A.A.; Woodward, J.J.; King, B.F.; Alkana, R.L. Ethanol differentially affects ATP-gated P2X(3) and P2X(4) receptor subtypes expressed in Xenopus oocytes. Neuropharmacology 2005, 49, 243–253. [Google Scholar] [CrossRef]
- Popova, M.; Asatryan, L.; Ostrovskaya, O.; Wyatt, L.R.; Li, K.; Alkana, R.L.; Davies, D.L. A point mutation in the ectodomain-transmembrane 2 interface eliminates the inhibitory effects of ethanol in P2X4 receptors. J. Neurochem. 2010, 112, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Ostrovskaya, O.; Asatryan, L.; Wyatt, L.; Popova, M.; Li, K.; Peoples, R.; Alkana, R.; Davies, D. Ethanol is a fast channel inhibitor of purinergic P2X4 receptors. J. Pharm. Exp. Ther. 2011, 337, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Young, M.T.; Fisher, J.A.; Fountain, S.J.; Ford, R.C.; North, R.A.; Khakh, B.S. Molecular Shape, Architecture, and Size of P2X4 Receptors Determined Using Fluorescence Resonance Energy Transfer and Electron Microscopy. J. Biol. Chem. 2008, 283, 26241–26251. [Google Scholar] [CrossRef] [PubMed]
- Silberberg, S.D.; Chang, T.H.; Swartz, K.J. Secondary structure and gating rearrangements of transmembrane segments in rat P2X4 receptor channels. J. Gen. Physiol. 2005, 125, 347–359. [Google Scholar] [CrossRef]
- Jelinkova, I.; Vavra, V.; Jindrichova, M.; Obsil, T.; Zemkova, H.W.; Zemkova, H.; Stojilkovic, S.S. Identification of P2X(4) receptor transmembrane residues contributing to channel gating and interaction with ivermectin. Pflugers Arch. 2008, 456, 939–950. [Google Scholar] [CrossRef]
- Stojilkovic, S.S.; Yan, Z.; Obsil, T.; Zemkova, H. Structural insights into the function of P2X4: An ATP-gated cation channel of neuroendocrine cells. Cell. Mol. Neurobiol. 2010, 30, 1251–1258. [Google Scholar] [CrossRef]
- Heymann, G.; Dai, J.; Li, M.; Silberberg, S.D.; Zhou, H.X.; Swartz, K.J. Inter- and intrasubunit interactions between transmembrane helices in the open state of P2X receptor channels. Proc. Natl. Acad. Sci. USA 2013, 110, E4045–E4054. [Google Scholar] [CrossRef]
- Asatryan, L.; Popova, M.; Perkins, D.I.; Trudell, J.R.; Alkana, R.L.; Davies, D.L. Ivermectin antagonizes ethanol inhibition in P2X4 receptors. J. Pharmacol. Exp. Ther. 2010, 334, 720–728. [Google Scholar] [CrossRef]
- Popova, M.; Trudell, J.; Li, K.; Alkana, R.; Davies, D.; Asatryan, L. Tryptophan 46 is a site for ethanol and ivermectin action in P2X4 receptors. Purinergic Signal. 2013. [Google Scholar] [CrossRef]
- Kawate, T.; Michel, J.C.; Birdsong, W.T.; Gouaux, E. Crystal structure of the ATP-gated P2X4 ion channel in the closed state. Nature 2009, 460, 592–598. [Google Scholar] [CrossRef]
- Hattori, M.; Gouaux, E. Molecular mechanism of ATP binding and ion channel activation in P2X receptors. Nature 2012, 485, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Silberberg, S.D.; Li, M.; Swartz, K.J. Ivermectin interaction with transmembrane helices reveals widespread rearrangements during opening of P2X receptor channels. Neuron 2007, 54, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Deber, C.M. Glycine and beta-branched residues support and modulate peptide helicity in membrane environments. FEBS Lett. 1992, 311, 217–220. [Google Scholar] [CrossRef]
- Polinsky, A.; Goodman, M.; Williams, K.A.; Deber, C.M. Minimum energy conformations of proline-containing helices. Biopolymers 1992, 32, 399–406. [Google Scholar] [CrossRef]
- Deber, C.M.; Khan, A.R.; Li, Z.; Joensson, C.; Glibowicka, M.; Wang, J. Val-->Ala mutations selectively alter helix-helix packing in the transmembrane segment of phage M13 coat protein. Proc. Natl. Acad. Sci. USA 1993, 90, 11648–11652. [Google Scholar] [CrossRef]
- Jindrichova, M.; Vavra, V.; Obsil, T.; Stojilkovic, S.S.; Zemkova, H. Functional relevance of aromatic residues in the first transmembrane domain of P2X receptors. J. Neurochem. 2009, 109, 923–934. [Google Scholar] [CrossRef]
- Ye, J.H.; Ren, J.; Liu, P.L.; McArdle, J.J. Glycine-activated chloride currents of neurons freshly isolated from the ventral tegmental area of rats. Brain Res. 1998, 796, 53–62. [Google Scholar] [CrossRef]
- Crawford, D.K.; Trudell, J.R.; Bertaccini, E.; Li, K.X.; Davies, D.L.; Alkana, R.L. Evidence that ethanol acts on a target in Loop 2 of the extracellular domain of alpha1 glycine receptors. J. Neurochem. 2007, 102, 2097–2109. [Google Scholar] [CrossRef]
- Perkins, D.I.; Trudell, J.R.; Crawford, D.K.; Alkana, R.L.; Davies, D.L. Targets for ethanol action and antagonism in loop 2 of the extracellular domain of glycine receptors. J. Neurochem. 2008, 106, 1337–1349. [Google Scholar] [CrossRef]
- Naito, A.; Muchhala, K.H.; Asatryan, L.; Trudell, J.R.; Homanics, G.E.; Perkins, D.I.; Davies, D.L.; Alkana, R.L. Glycine and GABA(A) ultra-sensitive ethanol receptors as novel tools for alcohol and brain research. Mol. Pharmacol. 2014, 86, 635–646. [Google Scholar] [CrossRef]
- Haines, W.R.; Migita, K.; Cox, J.A.; Terrance, M.E.; Voigt, M.M. The first transmembrane domain of the P2X receptor subunit participates in the agonist-induced gating of the channel. J. Biol. Chem. 2001, 276, 32793–32798. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Rassendren, F.; Spelta, V.; Suprenant, A.; North, R.A. Amino acid residues involved in gating identified in the first membrane-spanning domain of the rat P2X(2) receptor. J. Biol. Chem. 2001, 276, 14902–14908. [Google Scholar] [CrossRef]
- Samways, D.S.; Khakh, B.S.; Egan, T.M. Allosteric modulation of Ca2+ flux in ligand-gated cation channel (P2X4) by actions on lateral portals. J. Biol. Chem. 2012, 287, 7594–7602. [Google Scholar] [CrossRef]
- Egan, T.M.; Haines, W.R.; Voigt, M.M. A Domain Contributing to the Ion Channel of ATP-Gated P2X2 Receptors Identified by the Substituted Cysteine Accessibility Method. J. Neurosci. 1998, 18, 2350–2359. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Broomhead, H.E.; Young, M.T.; North, R.A. Polar residues in the second transmembrane domain of the rat P2X2 receptor that affect spontaneous gating, unitary conductance, and rectification. J. Neurosci. 2009, 29, 14257–14264. [Google Scholar] [CrossRef] [PubMed]
- Kracun, S.; Chaptal, V.; Abramson, J.; Khakh, B.S. Gated access to the pore of a P2X receptor: Structural implications for closed-open transitions. J. Biol. Chem. 2010, 285, 10110–10121. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptor | ATP | |||
|---|---|---|---|---|
| Imax, nA | EC50, μM | Hill Slope | N | |
| WT P2X4 | 4215 ± 678 | 5.74 ± 0.45 | 1.48 ± 0.10 | 9 |
| V49A | n.f. | n.d. | n.d. | 7 |
| F48A | 8716 ± 1533 # | 3.53 ± 0.73 * | 1.04 ± 0.09 * | 8 |
| V47A | 2049 ± 319 # | 5.0 ± 0.76 | 1.27 ± 0.06 | 7 |
| W46A | 3230 ± 498 | 3.58 ± 0.57 * | 1.02 ± 0.08 * | 10 |
| G45A | 5994 ± 1100 | 4.65 ± 0.54 | 1.16 ± 0.05 * | 10 |
| I44A | 4025 ± 687 | 5.68 ± 0.42 | 1.21 ± 0.05 | 11 |
| V43A | 6987 ± 1602 | 4.83 ± 0.59 | 1.34 ± 0.13 | 7 |
| Y42A | 1372 ± 119 ## | 4.33 ± 0.7 | 1.11 ± 0.25 | 5 |
| A41W | 1667 ± 233 ## | 4.62 ± 0.47 | 1.34 ± 0.07 | 12 |
| L40A | 1633 ± 282 ## | 6.41 ± 0.73 | 1.50 ± 0.10 | 8 |
| I39A | 5544 ± 1510 | 5.87 ± 0.70 | 1.39 ± 0.07 | 7 |
| L38A | 5719 ± 1211 | 7.33 ± 1.05 | 1.32 ± 0.06 | 8 |
| L37A | 5688 ± 749 | 8.91 ± 0.84 ** | 1.20 ± 0.08 | 7 |
| Q36A | 1568 ± 129 # | 6.35 ± 1.81 | 1.33 ± 0.14 | 4 |
| V35A | 7079 ± 1491 | 6.35 ± 0.55 | 1.36 ± 0.10 | 6 |
| A34W | 4386 ± 608 | 6.50 ± 1.02 | 1.21 ± 0.06 | 11 |
| R33A | 3545 ± 748 | 7.04 ± 0.51 | 1.03 ± 0.02 * | 5 |
| N32A | 2986 ± 397 | 5.40 ± 1.00 | 1.44 ± 0.08 | 7 |
| M31A | 3456 ± 375 | 7.07 ± 1.64 | 1.49 ± 0.23 | 7 |
| L30A | 7161 ± 1188 | 6.76 ± 0.84 | 1.37 ± 0.19 | 6 |
| G29A | 1175 ± 44 ## | 6.43 ± 0.34 | 1.33 ± 0.10 | 8 |
| Receptor | ATP | |||
|---|---|---|---|---|
| Imax, nA | EC50, μM | Hill Slope | N | |
| WT P2X4 | 5425 ± 1235 | 5.45 ± 0.4 | 1.47 ± 0.13 | 8 |
| V49P | n.f. | n.d. | n.d. | 5 |
| V49G | 2131 ± 292 * | 9.60 ± 0.9 * | 1.23 ± 0.10 | 6 |
| V49L | 2365 ± 277 | 10.60 ± 1.3* | 1.54 ± 0.18 | 4 |
| R33K | 5428 ± 986 | 6.33 ± 0.93 | 1.35 ± 0.09 | 9 |
| R33S | 2492 ± 912 | 4.94 ± 1.09 | 1.35 ± 0.22 | 6 |
| R33V | 5947 ± 1396 | 3.67 ± 1.28 | 0.86 ± 0.08 * | 6 |
| R33C | 5214 ± 560 | 3.96 ± 2.03 | 1.24 ± 0.36 | 7 |
| R33F | 4270 ± 791 | 4.68 ± 1.14 | 1.03 ± 0.03 | 7 |
| R33L | 2539 ± 418 * | 5.06 ± 0.70 | 0.85 ± 0.12 * | 9 |
| R33Y | n.f. | n.d. | n.d. | 11 |
| R33Q | n.f. | n.d. | n.d. | 3 |
| R33E | n.f. | n.d. | n.d. | 3 |
| Receptor | ATP | |||
|---|---|---|---|---|
| Imax, nA | EC50, μM | Hill Slope | N | |
| WT P2X4 | 2886 ± 614 | 5.34 ± 0.11 | 1.11 ± 0.08 | 7 |
| R33D | n.f. | n.d. | n.d. | 5 |
| D354R | n.f. | n.d. | n.d. | 6 |
| R33D-354R | n.f. | n.d. | n.d. | 6 |
| R33D:WT,1:1 | 763.3 ± 519 | 5.84 ± 0.11 | 1.45 ± 0.11 | 7 |
| D354R:WT, 1:1 | 560 ± 195 | 7.24 ± 1.07 | 1.2 ± 0.95 | 5 |
| R33D-D354R:WT, 1:1 | 540 ± 143 | 4.94 ± 1.06 | 1.2 ± 0.09 | 6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popova, M.; Rodriguez, L.; Trudell, J.R.; Nguyen, S.; Bloomfield, M.; Davies, D.L.; Asatryan, L. Residues in Transmembrane Segments of the P2X4 Receptor Contribute to Channel Function and Ethanol Sensitivity. Int. J. Mol. Sci. 2020, 21, 2471. https://doi.org/10.3390/ijms21072471
Popova M, Rodriguez L, Trudell JR, Nguyen S, Bloomfield M, Davies DL, Asatryan L. Residues in Transmembrane Segments of the P2X4 Receptor Contribute to Channel Function and Ethanol Sensitivity. International Journal of Molecular Sciences. 2020; 21(7):2471. https://doi.org/10.3390/ijms21072471
Chicago/Turabian StylePopova, Maya, Larry Rodriguez, James R. Trudell, Sylvia Nguyen, Michael Bloomfield, Daryl L. Davies, and Liana Asatryan. 2020. "Residues in Transmembrane Segments of the P2X4 Receptor Contribute to Channel Function and Ethanol Sensitivity" International Journal of Molecular Sciences 21, no. 7: 2471. https://doi.org/10.3390/ijms21072471
APA StylePopova, M., Rodriguez, L., Trudell, J. R., Nguyen, S., Bloomfield, M., Davies, D. L., & Asatryan, L. (2020). Residues in Transmembrane Segments of the P2X4 Receptor Contribute to Channel Function and Ethanol Sensitivity. International Journal of Molecular Sciences, 21(7), 2471. https://doi.org/10.3390/ijms21072471