Antibodies, Nanobodies, or Aptamers—Which Is Best for Deciphering the Proteomes of Non-Model Species?


1
School of Medicine, Deakin University, 75 Pigdons Road, Waurn Ponds, Victoria 3216, Australia
2
Centre for Molecular and Medical Research, Deakin University, 75 Pigdons Road, Waurn Ponds, Victoria 3216, Australia
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(7), 2485; https://doi.org/10.3390/ijms21072485
Submission received: 29 February 2020
/
Revised: 30 March 2020
/
Accepted: 30 March 2020
/
Published: 3 April 2020
(This article belongs to the Special Issue Proteomics and Nucleotide Profiling as Tools for Biomarker and Drug Target Discovery)
Abstract
:This planet is home to countless species, some more well-known than the others. While we have developed many techniques to be able to interrogate some of the “omics”, proteomics is becoming recognized as a very important part of the puzzle, given how important the protein is as a functional part of the cell. Within human health, the proteome is fairly well-established, with numerous reagents being available to decipher cellular pathways. Recent research advancements have assisted in characterizing the proteomes of some model (non-human) species, however, in many other species, we are only just touching the surface. This review considers three main reagent classes—antibodies, aptamers, and nanobodies—as a means of continuing to investigate the proteomes of non-model species without the complications of understanding the full protein signature of a species. Considerations of ease of production, potential applications, and the necessity for producing a new reagent depending on homology are presented.
1. Introduction
In the era of personalized medicine and the ability to have your own sample sent for genetic testing, there is still a perceived lack of progression in treating human disease. While genetic mutations are valuable for determining the presence (or absence) of a number of diseases, it is the proteins that are transcribed from the genes that are responsible for the incidence of diseases, as these proteins become the functional part of the cell. While the human genome project was instigated in 1990, technological advancements have meant that analyzing the protein signature, or proteome, has lagged behind. Since then, researchers have managed to achieve a commendable feat by sequencing the genomes of around 15,000 species. However, unfortunately, the number of complete proteomes is far shorter. With the number of eukaryotic species on this planet being in the tens of millions and the diversity of prokaryotic species reaching the trillions, we are still a long way from truly understanding the species-richness and variety of biology (Figure 1). We are slowly grasping the importance of understanding these functional units of different species and how these can aid in our understanding of disease and pathological processes [1]. While most of the research has been conducted on mammalian species, we have gained a good deal of knowledge in development and disease from other vertebrate species, with certain species of fish being utilized more and more over the last few years [2].
2. The Zebrafish as a Model Organism for Human Disease
Though the zebrafish has only recently gained popularity as a model organism, we have already made vital discoveries using zebrafish to understand developmental pathways, disease progression, and therapeutic strategies for diseases and disorders [3]. While there are a plethora of reasons why zebrafish serve as an ideal model for investigating immune cell development and disease progression, such as their transparent embryonic and larval developmental stages (reviewed in [4]), the primary reason for their widespread use is that their genome has been sequenced, making it easier to make significant comparisons between the genome and the proteome. This makes it easier to decipher results of studies undertaken in zebrafish to human development and disease processes, significantly improving our understanding of human illnesses.
There have been numerous studies that have investigated the proteome of zebrafish [5,6,7,8]. Most of these studies have utilized mass spectrometry and liquid chromatography-tandem mass spectrophotometry approach. In situ hybridization using RNA probes, which were developed nearly 10 years ago [9], have been extensively used in this field to study developmental pathways and immunological pathways in health and disease. While these approaches have expanded our understanding of the role of immune-modulatory and regulatory proteins in Zebrafish, the field has been impeded due to a lack of reliable antibodies or probes to investigate the pathways involved in health and disease. There have been attempts to generate reliable antibodies and probes to overcome this limitation. One such progress has been in the field of neurobiology, wherein researchers have demonstrated a panel of antibodies that are effective in visualizing neural receptors in zebrafish [10]. A monoclonal antibody against a tumor suppressor protein has also been generated [11]; however, the commercial use is yet to be elucidated. Thus, despite the availability of a complete genomic sequence, one of the major limitations has been the lack of appropriate antibodies or probes to study disease progression in zebrafish models.
What of other piscine species and what we can learn from them? Fish are the most numerous and phylogenetically diverse group of vertebrates, encompassing over 20,000 species [12,13]. They are also the oldest extant vertebrates, having inhabited the earth for more than 500 million years [14,15,16,17]. Given this great diversity, along with a diverse pattern of ageing, a small size, and ease of cultivation, there is much that can be gathered from these species. This will open new avenues for us to understand the different biological processes, including disease modelling, response to external stimuli (such as infections, environmental changes), ageing, and response to drugs [18,19,20].
The molecular pathways involved in immunity are highly conserved, with the innate immune system dating back to the early metazoans, and the acquired immune system dating back to the origin of the jawed vertebrates about 450 million years ago [21,22]. When a pathogen invades a host, it stimulates a number of these conserved pathways [22,23], which leads to the altered expression of key genes that serve as sensitive markers of infection. The innate immune response is generally mounted early in response to invading organisms and includes a complex network of molecules and cells, both specific and non-specific, which operate to clear the pathogen from its host. Some of the main cells involved in these innate responses are leucocytes (including neutrophils, macrophages, and dendritic cells). Their primary role is to phagocytose and digest the pathogen to present it to adaptive immune cells (such as T and B cells), or kill/eliminate the pathogen by producing antimicrobial molecules and immune-modulatory proteins. Some of the molecules include antibacterial peptides (cathelicidins, human β defensins), lysozyme, transferrin, complement, acute-phase proteins, prostaglandins (PGE2), reactive oxygen intermediates (ROI), cyclooxygenase-2 (COX-2), cytokines, chemokines, and lectins [24,25]. While a vast amount of information regarding the immune system in mammals exists, transferring this knowledge to fish has led to some difficulties. This is due to a number of issues, not least of which is the fact that fish species live in their own unique environment. However, the newly acquired ability in molecular biology to sequence and clone genes has become a powerful system for the study of vertebrate immune development and disease [25]. Indeed, even though fish immunology at the molecular level is becoming well understood, functional biology of fish immune cells, both in vivo and in vitro, is still in its infancy [25]. Undoubtedly, this field has witnessed significant improvements over the last ten years, but there is still much that we do not know and the majority of research to date has focused on commercially important species and biosecurity [26].
Comparative immunologists have been interested in the immune system of fish and how it has similarities and differences with higher vertebrates [27,28,29]. However, the lagging step has been the production of suitable reagents to recognize piscine targets [30]. While the genetics and transcripts for a number of piscine species have been described, this does not provide sufficient functional knowledge of pathways activated during disease processes. To do that, knowledge of a particular protein sequence is required in order to generate reagents. With the availability of CRISPR-based technology, analyzing immunological pathways has become possible in the zebrafish,. Overall, while there is a large scope for generation of tools to study protein–protein interactions in fish species, the very slow process, even with technological advancements, to add knowledge regarding both piscine and broader species, and expanding the field of comparative immunology remains a key challenge.
3. Proteomes of Larger Species
The first step to understanding the complexities of the proteome across multiple animals, including humans, is to sequence the genomes. The Earth BioGenomes project was announced in 2018 to “sequence life for future life” [31]. The initial goal of this project was to generate annotated chromosome-scale reference assemblies for a minimum of one reference species across each of the approximately 9000 eukaryotic taxonomic families. Once this is completed, it will be possible to look for percentage homology amongst species and begin the process of assessing which species may have similar proteomes. This information, probably more so than with the use of smaller vertebrates or non-mammalian models, will provide more knowledge on predicting which animal model systems are more appropriate for studying specific disease states and lead to the development of a more refined list of species specific models rather than the reliance on mice and rats.
As discussed earlier, the molecular pathways involved in immunity are highly conserved across species, indicating that responses that are generated by hosts (human or non-human species) to infections are extremely likely to be similar. However, when attempting to model the response to an infection, or even how an infection spreads, the choice of animal model is complex. It is important to note that during an immune response to an infection or allergen, there is not only involvement of the immune system, but also the interplay with other pathways, including the neuroendocrine and physiological systems. The differences (and similarities) in these other physiological systems between humans and other species are bound to have an effect on the overall understanding of a particular disease or treatment. For new and emerging infections, an animal model which can predict the pathogenesis of the disease and respond in a similar manner to therapeutics or vaccines is required [32]. An example would be the use of ferrets for modelling influenza infections. Ferrets are one of the few animal models that exhibit most of the clinical symptoms observed in humans, and they can be easily infected with human influenza viruses and transmit the disease efficiently between each other [33]. While the number of studies using ferrets as an animal model for influenza has increased since 2008, it was not until 2011 that the proteome was deposited in UniProt.org. The ferret is also now being used as a model for cystic fibrosis research [34]. While there is a paucity of reagents [33], the National Ferret Resource and Research Center has compiled a list of commercially available antibodies that are applicable to ferret proteins.
Staying on the theme of animal models for human disease, it is not only knowledge of proteomes that can aid us in understanding which model to choose, but also which model not to use in pre-clinical studies. For example, mice and pigs have been used in cardiac research but with limited success. A recent study sought to elucidate the proteomes of cardiac development in the mouse, pig, and Xenopus to better understand the most appropriate model for modelling cardiac disease. Proteins known to be expressed in humans were also found to be enriched in frogs, but surprisingly not in mice or pigs [35]. It is unsurprising then that frogs may find their way into more translatable research in the future [36], especially given that half of human genes differ from their mouse orthologs in different developmental trajectories, including more than 200 disease genes associated with brain, heart, and liver diseases [37]. These differences can impact the proteomes, and therefore phenotypes, between humans and mice. This leads to the necessity of reliable reagents needed for interrogating the protein signatures of animal models for human disease.
When it comes to assessment of other animals, specifically in regard to the animals that are important to farming, proteome research has been very limited (Table 1). The majority of the research has largely focused on animal products rather than the animal itself. This can be attributed to two main reasons: high costs and a lack of genomic data. Where there has been a focus on animals, it has been driven by predictors of food quality or the detection of infectious diseases. In instances where traditional proteome research techniques, such as shotgun-based approaches, have been utilized in this field, these have missed proteins of relevance to research and health, such as cytokines and their receptors. This may be due to the low concentration of these immune-modulatory proteins in the biological samples when compared to other proteins, which increases the chances of them being undetected during analysis. When considering food security for the future, it is important that a different approach is considered to identify disease biomarkers and biomarkers relevant to health and immune function [38,39].
It is worth noting that while there is limited information available for animals important to humans as food sources, there is a lack of credible information available for native species. The Peptide Atlas has information on cows, horses, mice, pigs, rats, and zebrafish. UniProt contains more data for native species, though it mostly has not been reviewed. From an Australian perspective, proteomes are available for the platypus and the Tasmanian devil, but not the echidna, koala, kangaroo, or possum, leading to an extreme paucity of data for species of ecological significance.
4. Towards a Better System of Detecting Proteome Signatures
So if we are no closer to understanding the proteomes of all the species on this planet, or even those that are classed as important to humans, such as farm animals, pets, and wildlife, are there ways we can adapt current technology to interrogate proteins that appear to be similar or related to diseases across species? The main stay in human disease research over the past 40 years has been monoclonal antibodies [52]. These have proven to be very effective in diagnostic applications where the antibody has been fully validated and quality controls are in place to ensure each batch of antibody performs to the strict requirements [53]. However, in research applications there have been several limitations, notwithstanding the reproducibility crisis that has been placed at the feet of reagents, and antibodies in particular, in a high percentage of cases [54].
In order to generate an antibody, purified protein is required and this protein needs to be immunogenic in order to generate an immune response once the protein is injected into an animal host [30]. Following the time required to initiate an immune response and generate antibodies, the B-cells of the spleen are removed and fused with myeloma cells to form hybridomas which are then tested to identify the ones which produce the best monoclonal antibodies for their target [55]. This process has limitations, as you need to know what your target, or protein, is to begin with. Now, with the advent of in vitro combinatorial display libraries, it may be possible to identify antibody fragments that bind to a particular protein. However, the peptide fragments may not necessarily show reactivity to other species, as the peptides are selected on the Fc region of typically human IgG [56]. There are commercial phage display libraries derived from mouse, rabbit, chicken, camel, and llama (creative-biolabs.com), and there may be some cross-reactivity to other species, though the ability of these combinatorial libraries to be used for native species is limited given the probable differences in genome and proteome homology (Figure 2). Additionally, if an antibody is discovered from one of these libraries, its applications may be limited to diagnostic applications, rather than theranostic, due to their likely immunogeneic nature.
The discovery that camels have IgG-like material in their serum that showed similarity to antibodies but were devoid of light chains suggested new opportunities for the engineering of antibodies [57]. These camelid single chain antibodies show a high sequence homology to the variable domains of heavy chains in humans (80 %) [58]. The highly stable single antigen binding domain is much smaller than a conventional antibody, a key benefit, as they are only about 15 kDa, which has led to them being known as nanobodies [59,60]. This may allow them to bind to antigenic sites not recognized by antibodies or to “hidden” antigenic sites [58]. Due to their similarity to human immunoglobulins, they have a low immunogenic profile in humans [61], although the same probably could not be said for other species, which presents a limitation for wider therapeutic applications. In terms of selecting nanobody phage libraries, the process is similar to the phage display libraries used to select for antibodies. Interestingly, these nanobody libraries have recently been used to select for cell surface receptors in situ as well as intracellular targets [62,63], which does open up possibilities for selecting nanobodies against unknown targets on particular sub-populations of cells. Additionally, nanobodies have been generated that are cross-reactive to human and murine proteins [64] or all Trypanosoma species [65].
Given the challenges associated with the use of antibodies and nanobodies in therapeutic applications, and lack of cross-species reactivity, other classes of molecules are required to advance our knowledge in other species. A comparatively recent advancement in the field has been the generation of aptamers, which are single stranded DNA or RNA sequences that bind to their targets in a similar manner to antibodies and nanobodies—through shape recognition to their target binding sites. In much the same way as combinatorial libraries of antibodies and nanobodies are used, combinatorial libraries of nucleic acid sequences are used to select aptamers that bind to a target. This process, known as the systematic evolution of ligands by exponential enrichment (SELEX), was first described in 1990 [66,67,68] and involves a library of approximately 1014 individual sequences that are incubated with a target over iterative rounds to “evolve” a number of sequences that bind with high specificity and sensitivity. These binding sequences are typically 6–12 kDa in size and therefore have similar properties to nanobodies in that they can bind to antigenic sites that are not recognized by antibodies (Figure 2). For pathways that are highly conserved [69], it is possible that a “binding site” might be large enough and homologous enough that antibodies may show cross-reactivity amongst species. The smaller the binding molecule though, the more likely the recognition. However, where there is divergence and homology is decreased, the binding site may only be amenable to very small binders that have been generated to other species’ proteins. In a similar manner to nanobodies being smaller versions of antibodies, an aptamer is fairly easy to truncate to its smallest functional form, which may increase its possibility of binding to multiple species [70,71,72]
Of note for the use of combinatorial libraries, it is possible to take the sequence of the targeting moiety and mutate this sequence with multiple mutations to find one that will bind to a different species’ protein, though depending on the format of the assay and the knowledge of the other protein, this may not be possible [73]. However, there are several other protocols available that might be amenable for developing targeting agents. For example, a protocol amendment for aptamer development that reacts to human and porcine thrombin was first described in 2001 [74]. This process was coined “toggle SELEX” and involved interchanging the protein in selection rounds between the two. There have since been several adaptations to this protocol to generate cross-species binding aptamers [75,76,77].
5. Fishing for Homology: Potential Protocols for Developing Targeting Agents
It is still possible to determine cross-species affinity without complete knowledge of the genomes and proteomes. If sufficient protein can be purified, this can be the starting point for screening against commercially available antibodies. A study by Villarreal and colleagues purified protein from zebrafish to clone into the recombinant SUMO solubility tag expression system to test an antibody from Abcam that had been generated for a human protein [78]. Cross-species reactivity of antibodies has also been shown between fish species [79]. Studies using aptamers and nanobodies are limited in the literature, though this is an ever-expanding space. However, most of the research has been limited to human and murine cross-reactivity for translational studies, with very few studies looking at other species. Interestingly, and highlighting the point earlier made about the size of the binding region, there have been a few studies that have truncated aptamers to enhance their cross-species reactivity. For example, Dhiman et al. truncated an aptamer from 40 nucleotides to 26 nucleotides to recognize toxins from two different snake species [70]. One study has investigated the ability of three different aptamers to bind to thrombin in six different species (human, bovine, porcine, rabbit, rat, and mouse). The shortest aptamer sequence, comprising 15 nucleotides, had the least variation in activity compared to the other two longer sequences [80]. These studies, taken together, suggest that truncating aptamers to their smallest possible functional units would enhance researchers’ abilities to interrogate cellular pathways in non-model animal models.
Purified protein or a known protein sequence is not necessarily a limiting factor for the generation of a targeting ligand. Selection can be attempted without knowing the precise target if sufficient materials, such as specific cells, are available. For example, nanobody libraries have been used to select for cell surface receptors [63], though in this case the proteins were expressed in cells prior to selection. This can also be performed using aptamer libraries [81] though it is also possible to “fish” for targets on whole cells or organisms [82,83]. It is even possible to develop antibodies against targets on whole cells using the combinatorial libraries [84]. A schematic is provided in Figure 3 to demonstrate the selection process. The protocol used for generating nanobodies to cell surface receptors used a process similar to SELEX whereby the nanobody library was incubated with target cells for iterative rounds and bound species were re-amplified using Escherichia coli rather than using PCR to re-amplify the bound nucleic acid species [63,85]. This process can be expanded on through the use of tissue sections, and both antibodies and aptamers have been generated to rare cells using both frozen [86,87] and paraffin embedded tissue as the antigen substrate [88]. Where homology is suggested, rather than undergoing a complete selection of a full combinatorial library, it is possible to use a directed library. For example, mutations can be introduced into the known binding sequence to generate a much smaller library that may require only one incubation step to detect a targeting ligand. This is an adaptation of the process underpinning panning for better affinity or novel therapeutic mechanisms [89,90]. This process may be simpler with aptamers, due to the smaller sequences and nucleic acid nature which allows base pair switching or error-prone PCR to introduce mutations [73].
Where no knowledge of the homology of the protein, or even the protein target, is available, it may still be possible to develop targeting regents. Traditionally, antibodies and nanobodies have been generated following an immunization event, with antibodies/nanobodies being collected from the B-cells of the spleen. An improvement on this has been the injection of combinatorial libraries into target animals to collect those that have bound to a target organ or cell type [91], a method that has also been achieved with aptamer libraries [92]. This method has the benefit of providing aptamers or antibodies that are relevant for in vivo applications, though it does present some technical challenges.
6. In Vitro Applications: Which is Best?
Given that antibody, nanobody, and aptamer selection can all now be based on combinatorial libraries, are there benefits to using one over the other? This comes down to what the future application(s) may be (Table 2). Antibodies and nanobodies are proteins, and so have requirements for limited changes to pH, salt composition, and temperature, as extreme conditions can denature them. As well, selection conditions need to be close to physiological conditions due to the risk of affecting the protein structure. This means that the possibility of developing antibodies or nanobodies to targets in complex media or non-physiological conditions is limited. Aptamers are not limited by these parameters, and selection conditions can be modified to ensure the aptamers will work in any application [93]. Indeed, with the use of modified nucleotides, it is possible to select aptamers in very acidic or alkaline conditions [94,95]. One other major drawback of antibodies and nanobodies over aptamers is the cost involved in generating them [96].
While traditional antibody tests, such as enzyme linked immunosorbent assays (ELISAs) and immunophenotyping by flow cytometry are completed in a laboratory environment, there is a requirement for point of care testing (POCT) and rapid diagnostic tests that span all species [97,98,99]. Going back to the protein nature of antibodies and nanobodies, these require special conditions to be used in the field for POCT. For example, transport needs to be refrigerated and antibodies/nanobodies need to be kept cool to ensure they remain functional, whereas aptamers are stable at higher temperatures [53,100]. Additionally, while aptamers have been used to replace antibodies in typical laboratory tests, they have also come into their own in a number of in vitro diagnostic assays that would be easily adapted to in field or POCT [101,102,103,104].
An avenue where aptamers are being utilized due to the diverse advantages they offer is the rapidly emerging field of handheld electrochemical sensors. Aptamers are smaller in size, have a much more flexible nature, and have better conductivity than antibodies [97,105]. Add in the issues of stability and protein biofouling, and it not strange that aptamers have gained popularity in this field in recent years [106]. Indeed, aptamer based electrochemical sensors have been used in many fields, such as clinical diagnostics, food analysis, and environmental science [107]. While these devices have been used to detect small molecules, proteins, and even cells in clinically relevant environments, they have not yet been used in non-model organisms [97,105,107]. As researchers start to investigate these non-model organisms to understand development or disease pathology, and to understand the health and well-being of species, the field of aptamer research is likely to change and expand.
7. Conclusions
This review has been kept deliberately broad due to the nature of this area of research being almost all encompassing. With so many species on the planet still waiting for complete genomes, let alone proteomes, to be completed so we have a better idea of the Tree of Life, there are some reagents that can be directly used to interrogate the proteins of other species. However, for the vast majority of species, new reagents need to be generated to advance knowledge. These reagents can be antibodies, nanobodies, or aptamers, with the choice being ease of development, ease of use, and future applications. Some may be more likely to cross-react across species than others, depending on the size of target binding site. However, with antibodies dominating the diagnostic and research field for 40 years, it may now be necessary to investigate the benefits offered by nanobodies and aptamers as alternative affinity reagents to decipher similarities and differences in evolutionary pathways and the implications of these developmental changes between species.
Author Contributions
P.D., R.M.S. and S.S. contributed to the conception and design of the review, as well as development of the figures, manuscript text and revision. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| COX-2 | Cyclooxygenase |
| MYA | Million years ago |
| PGE2 | prostaglandins |
| POCT | Point of care test |
| ROI | Reactive oxygen species |
| SELEX | Systematic evolution of ligands by exponential enrichment |
References
- Gotelli, N.J.; Ellison, A.M.; Ballif, B.A. Environmental proteomics, biodiversity statistics and food-web structure. Trends Ecol. Evol. 2012, 27, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teame, T.; Zhang, Z.; Ran, C.; Zhang, H.; Yang, Y.; Ding, Q.; Xie, M.; Gao, C.; Ye, Y.; Duan, M.; et al. The use of zebrafish (Danio rerio) as biomedical models. Anim. Front. 2019, 9, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, Y.M.; Toro, S.; Ramachandran, S.; Ruzicka, L.; Howe, D.G.; Eagle, A.; Kalita, P.; Martin, R.; Taylor Moxon, S.A.; Schaper, K.; et al. Zebrafish Models of Human Disease: Gaining Insight into Human Disease at ZFIN. ILAR J. 2017, 58, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patton, E.E.; Tobin, D.M. Spotlight on Zebrafish: The Next Wave of Translational Research. Dis. Model Mech. 2019. [Google Scholar] [CrossRef] [Green Version]
- Kessels, M.Y.; Huitema, L.F.A.; Boeren, S.; Kranenbarg, S.; Schulte-Merker, S.; van Leeuwen, J.L.; de Vries, S.C. Proteomics Analysis of the Zebrafish Skeletal Extracellular Matrix. PLoS ONE 2014, 9, e90568. [Google Scholar] [CrossRef] [Green Version]
- Abramsson, A.; Westman-Brinkmalm, A.; Pannee, J.; Gustavsson, M.; von Otter, M.; Blennow, K.; Brinkmalm, G.; Kettunen, P.; Zetterberg, H. Proteomics Profiling of Single Organs from Individual Adult Zebrafish. Zebrafish 2010, 7, 161–168. [Google Scholar] [CrossRef]
- Li, C.; Tan, X.F.; Lim, T.K.; Lin, Q.; Gong, Z. Comprehensive and quantitative proteomic analyses of zebrafish plasma reveals conserved protein profiles between genders and between zebrafish and human. Sci. Rep. 2016, 6, 24329. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Sundaram, C.S.; Shanbhag, S.; Idris, M.M. Proteomic Profile of Zebrafish Brain Based on Two-Dimensional Gel Electrophoresis Matrix-Assisted Laser Desorption/Ionization MS/MS Analysis. Zebrafish 2010, 7, 169–177. [Google Scholar] [CrossRef]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef]
- Staudt, N.; Müller-Sienerth, N.; Fane-Dremucheva, A.; Yusaf, S.P.; Millrine, D.; Wright, G.J. A panel of recombinant monoclonal antibodies against zebrafish neural receptors and secreted proteins suitable for wholemount immunostaining. Biochem. Biophys. Res. Commun. 2015, 456, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.C.; Goh, W.L.P.; Xu, M.; Kua, N.; Lunny, D.; Wong, J.S.; Coomber, D.; Vojtesek, B.; Lane, E.B.; Lane, D.P. Detection of the p53 response in zebrafish embryos using new monoclonal antibodies. Oncogene 2008, 27, 629–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, K.; Fishelson, L.; Amselgruber, W.M. Cytological ontogenesis and involution of the thymus and head-kidney in juvenile and old domestic carp: Is ageing in fish a chronological or growth-related phenomenon? J. Appl. Ichthyol. 2001, 17, 1–13. [Google Scholar] [CrossRef]
- Spitsbergen, J.M.; Kent, M.L. The State of the Art of the Zebrafish Model for Toxicology and Toxicologic Pathology Research—Advantages and Current Limitations. Toxicol. Pathol. 2003, 31, 62–87. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.M.; Maclean, N. Teleost microarrays: Development in a broad phylogenetic range reflecting diverse applications. J. Fish Biol. 2008, 72, 2039–2050. [Google Scholar] [CrossRef]
- Hine, P.M. The granulocytes of fish. Fish Shellfish Immunol. 1992, 2, 79–98. [Google Scholar] [CrossRef]
- Jarvik, E. Basic Structure and Evolution of Vertebrates; Academic Press: New York, NY, USA, 1980; Volume 1. [Google Scholar]
- Nelson, J.S. Fishes of the World, 3rd ed.; John Wiley & Sons: New York, NY, USA, 1994. [Google Scholar]
- Gerhard, G.S. Small laboratory fish as models for aging research. Ageing Res. Rev. 2007, 6, 64–72. [Google Scholar] [CrossRef]
- Brown, A.R.; Gunnarsson, L.; Kristiansson, E.; Tyler, C.R. Assessing variation in the potential susceptibility of fish to pharmaceuticals, considering evolutionary differences in their physiology and ecology. Philos. Trans R. Soc. Lond. B Biol. Sci. 2014, 369, 20130576. [Google Scholar] [CrossRef] [Green Version]
- Fazio, M.; Zon, L.I. Fishing for answers in precision cancer medicine. Proc. Natl. Acad. Sci. USA 2017, 114, 10306. [Google Scholar] [CrossRef] [Green Version]
- Hartenstein, V. Blood Cells and Blood Cell Development in the Animal Kingdom. Ann. Rev. Cell Dev. Biol. 2006, 22, 677–712. [Google Scholar] [CrossRef] [Green Version]
- Magnadóttir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Bryant, P.A.; Venter, D.; Robins-Browne, R.; Curtis, N. Chips with everything: DNA microarrays in infectious diseases. Lancet Infect. Dis. 2004, 4, 100–111. [Google Scholar] [CrossRef]
- Ellis, A.E. Immunity to bacteria in fish. Fish Shellfish Immunol. 1999, 9, 291–308. [Google Scholar] [CrossRef]
- Randelli, E.; Buonocore, F.; Scapigliati, G. Cell markers and determinants in fish immunology. Fish Shellfish Immunol. 2008, 25, 326–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, C.; Low, C. Synthetic antibody: Prospects in aquaculture biosecurity. Fish Shellfish Immunol. 2019, 86, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, H.W.; Findly, R.C. Vertebrate Adaptive Immunity—Comparative Insights from a Teleost Model. Front. Immunol. 2017, 8, 1379. [Google Scholar] [CrossRef] [Green Version]
- Scapigliati, G.; Fausto, A.M.; Picchietti, S. Fish Lymphocytes: An Evolutionary Equivalent of Mammalian Innate-Like Lymphocytes? Front. Immunol. 2018, 9, 971. [Google Scholar] [CrossRef] [Green Version]
- Johnston, H.J.; Verdon, R.; Gillies, S.; Brown, D.M.; Fernandes, T.F.; Henry, T.B.; Rossi, A.G.; Tran, L.; Tucker, C.; Tyler, C.R.; et al. Adoption of in vitro systems and zebrafish embryos as alternative models for reducing rodent use in assessments of immunological and oxidative stress responses to nanomaterials. Crit. Rev. Toxicol. 2018, 48, 252–271. [Google Scholar] [CrossRef]
- Dixon, B.; Barreda, D.R.; Sunyer, J.O. Perspective on the Development and Validation of Ab Reagents to Fish Immune Proteins for the Correct Assessment of Immune Function. Front. Immunol. 2018, 9, 2957. [Google Scholar] [CrossRef]
- Lewin, H.A.; Robinson, G.E.; Kress, W.J.; Baker, W.J.; Coddington, J.; Crandall, K.A.; Durbin, R.; Edwards, S.V.; Forest, F.; Gilbert, M.T.P.; et al. Earth BioGenome Project: Sequencing life for the future of life. Proc. Natl. Acad. Sci. USA 2018, 115, 4325. [Google Scholar] [CrossRef] [Green Version]
- Swearengen, J.R. Choosing the right animal model for infectious disease research. Anim. Models. Exp. Med. 2018, 1, 100–108. [Google Scholar] [CrossRef]
- Oh, D.Y.; Hurt, A.C. Using the Ferret as an Animal Model for Investigating Influenza Antiviral Effectiveness. Front. Microbiol. 2016, 7, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keiser, N.W.; Birket, S.E.; Evans, I.A.; Tyler, S.R.; Crooke, A.K.; Sun, X.; Zhou, W.; Nellis, J.R.; Stroebele, E.K.; Chu, K.K.; et al. Defective innate immunity and hyperinflammation in newborn cystic fibrosis transmembrane conductance regulator-knockout ferret lungs. Am. J. Respir. Cell Mol. Biol. 2015, 52, 683–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federspiel, J.D.; Tandon, P.; Wilczewski, C.M.; Wasson, L.; Herring, L.E.; Venkatesh, S.S.; Cristea, I.M.; Conlon, F.L. Conservation and divergence of protein pathways in the vertebrate heart. PLoS Biol. 2019, 17, e3000437. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Ott, T. Xenopus: An Undervalued Model Organism to Study and Model Human Genetic Disease. Cells Tissues Organs 2018, 205, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Cardoso Moreira, M.; Velten, B.; Mort, M.; Cooper, D.; Huber, W.; Kaessmann, H. Developmental Gene Expression Differences between Humans and Mammalian Models. bioRxiv 2019. [Google Scholar] [CrossRef]
- Almeida, A.M.; Bassols, A.; Bendixen, E.; Bhide, M.; Ceciliani, F.; Cristobal, S.; Eckersall, P.D.; Hollung, K.; Lisacek, F.; Mazzucchelli, G.; et al. Animal board invited review: Advances in proteomics for animal and food sciences. Animal 2015, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Campos, M.O.A.; De Almeida, M.A. Top-Down Proteomics and Farm Animal and Aquatic Sciences. Proteomes 2016, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Basit, A.; Radi, Z.; Vaidya, V.S.; Karasu, M.; Prasad, B. Kidney Cortical Transporter Expression across Species Using Quantitative Proteomics. Drug Metab. Dispos. 2019, 47, 802. [Google Scholar] [CrossRef]
- Fallon, J.K.; Smith, P.C.; Xia, C.Q.; Kim, M.-S. Quantification of Four Efflux Drug Transporters in Liver and Kidney Across Species Using Targeted Quantitative Proteomics by Isotope Dilution NanoLC-MS/MS. Pharm. Res. 2016, 33, 2280–2288. [Google Scholar] [CrossRef]
- Golizeh, M.; Schneider, C.; Ohlund, L.B.; Sleno, L. Multidimensional LC–MS/MS analysis of liver proteins in rat, mouse and human microsomal and S9 fractions. EuPA Open Proteom. 2015, 6, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhao, X.; Huang, D.; Pan, X.; Qi, Y.; Yang, Y.; Zhao, H.; Cheng, G. Proteomic analysis and cross species comparison of casein fractions from the milk of dairy animals. Sci. Rep. 2017, 7, 43020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Wang, X.; Zhang, W.; Liu, L.; Pang, X.; Zhang, S.; Lv, J. Comparative proteomics of milk fat globule membrane in different species reveals variations in lactation and nutrition. Food Chem. 2016, 196, 665–672. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, N.; Wang, W.; Zhao, X.; Zhang, Y.; Han, R.; Ma, L.; Zhao, S.; Li, S.; Guo, T.; et al. N-glycosylation proteomic characterization and cross-species comparison of milk fat globule membrane proteins from mammals. Proteomics 2016, 16, 2792–2800. [Google Scholar] [CrossRef] [PubMed]
- Martens, G.A. Species-Related Differences in the Proteome of Rat and Human Pancreatic Beta Cells. J. Diabetes Res. 2015, 2015, 549818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulo, J.A.; Urrutia, R.; Kadiyala, V.; Banks, P.; Conwell, D.L.; Steen, H. Cross-species analysis of nicotine-induced proteomic alterations in pancreatic cells. Proteomics 2013, 13, 1499–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Leng, T.; Yun, D.; Liu, N.; Yao, J.; Dai, Y.; Yang, P.; Chen, X. Global analysis of the rat and human platelet proteome—The molecular blueprint for illustrating multi-functional platelets and cross-species function evolution. Proteomics 2010, 10, 2444–2457. [Google Scholar] [CrossRef] [Green Version]
- de Sousa-Pereira, P.; Cova, M.; Abrantes, J.; Ferreira, R.; Trindade, F.; Barros, A.; Gomes, P.; Colaço, B.; Amado, F.; Esteves, P.J.; et al. Cross-species comparison of mammalian saliva using an LC–MALDI based proteomic approach. Proteomics 2015, 15, 1598–1607. [Google Scholar] [CrossRef]
- Druart, X.; Rickard, J.P.; Mactier, S.; Kohnke, P.L.; Kershaw-Young, C.M.; Bathgate, R.; Gibb, Z.; Crossett, B.; Tsikis, G.; Labas, V.; et al. Proteomic characterization and cross species comparison of mammalian seminal plasma. J. Proteom. 2013, 91, 13–22. [Google Scholar] [CrossRef]
- Bayram, H.L.; Claydon, A.J.; Brownridge, P.J.; Hurst, J.L.; Mileham, A.; Stockley, P.; Beynon, R.J.; Hammond, D.E. Cross-species proteomics in analysis of mammalian sperm proteins. J. Proteom. 2016, 135, 38–50. [Google Scholar] [CrossRef]
- KÖHler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- Bauer, M.; Strom, M.; Hammond, S.D.; Shigdar, S. Anything You Can Do, I Can Do Better: Can Aptamers Replace Antibodies in Clinical Diagnostic Applications? Molecules 2019, 24, 4377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M. 1,500 scientists lift the lid on reproducibility. Nature 2016, 533, 452–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaroff, S.; Tan, G. Hybridoma technology: The preferred method for monoclonal antibody generation for in vivo applications. Biotechniques 2019, 67, 90–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vodnik, M.; Zager, U.; Strukelj, B.; Lunder, M. Phage display: Selecting straws instead of a needle from a haystack. Molecules 2011, 16, 790–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hammers, C.; Songa, E.B.; Bendahman, N.; Hammers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.M.; De Haard, H.J. Properties, production, and applications of camelid single-domain antibody fragments. Appl. Microbiol. Biotechnol. 2007, 77, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Wagner, H.J.; Wehrle, S.; Weiss, E.; Cavallari, M.; Weber, W. A Two-Step Approach for the Design and Generation of Nanobodies. Int. J. Mol. Sci. 2018, 19, 3444. [Google Scholar] [CrossRef] [Green Version]
- Bannas, P.; Hambach, J.; Koch-Nolte, F. Nanobodies and Nanobody-Based Human Heavy Chain Antibodies as Antitumor Therapeutics. Front. Immunol. 2017, 8, 1603. [Google Scholar] [CrossRef]
- Jovčevska, I.; Muyldermans, S. The Therapeutic Potential of Nanobodies. BioDrugs 2020, 34, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Woods, J. Selection of Functional Intracellular Nanobodies. SLAS Discov. 2019, 24, 703–713. [Google Scholar] [CrossRef]
- Crepin, R.; Veggiani, G.; Djender, S.; Beugnet, A.; Planeix, F.; Pichon, C.; Moutel, S.; Amigorena, S.; Perez, F.; Ghinea, N.; et al. Whole-cell biopanning with a synthetic phage display library of nanobodies enabled the recovery of follicle-stimulating hormone receptor inhibitors. Biochem. Biophys. Res. Commun. 2017, 493, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Kazemi-Lomedasht, F.; Pooshang-Bagheri, K.; Habibi-Anbouhi, M.; Hajizadeh-Safar, E.; Shahbazzadeh, D.; Mirzahosseini, H.; Behdani, M. In vivo immunotherapy of lung cancer using cross-species reactive vascular endothelial growth factor nanobodies. Iran. J. Basic Med. Sci. 2017, 20, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Obishakin, E.; Stijlemans, B.; Santi-Rocca, J.; Vandenberghe, I.; Devreese, B.; Muldermans, S.; Bastin, P.; Magez, S. Generation of a Nanobody Targeting the Paraflagellar Rod Protein of Trypanosomes. PLoS ONE 2015, 9, e115893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.; Szostak, J. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Joyce, G.F. Selection in vitro of an RNA enzyme that specifically cleaves single-stranded DNA. Nature 1990, 344, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Riera Romo, M.; Pérez-Martínez, D.; Castillo Ferrer, C. Innate immunity in vertebrates: An overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef]
- Dhiman, A.; Anand, A.; Malhotra, A.; Khan, E.; Santra, V.; Kumar, A.; Sharma, T.K. Rational truncation of aptamer for cross-species application to detect krait envenomation. Sci. Rep. 2018, 8, 17795. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, J.; Denoyer, D.; Henri, J.; Jamieson, A.; Burvenich, I.J.G.; Pouliot, N.; Shigdar, S. Bifunctional Aptamer-Doxorubicin Conjugate Crosses the Blood-Brain Barrier and Selectively Delivers Its Payload to Epithelial Cell Adhesion Molecule-Positive Tumor Cells. Nucleic Acid Ther. 2020. [Google Scholar] [CrossRef]
- Macdonald, J.; Houghton, P.; Xiang, D.; Duan, W.; Shigdar, S. Truncation and mutation of a transferrin receptor aptamer enhances binding affinity. Nucleic Acid Ther. 2016, 26, 348–354. [Google Scholar] [CrossRef]
- McCullum, E.; Williams, B.; Zhang, J.; Chaput, J. Random Mutagenesis by Error-Prone PCR. Methods Mol. Biol. 2010, 634, 103–109. [Google Scholar] [CrossRef] [PubMed]
- White, R.; Rusconi, C.; Scardino, E.; Wolberg, A.; Lawson, J.; Hoffman, M.; Sullenger, B. Generation of species cross-reactive aptamers using toggle SELEX. Mol. Ther. 2001, 4, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Levay, A.; Brenneman, R.; Hoinka, J.; Sant, D.; Cardone, M.; Trinchieri, G.; Przytycka, T.M.; Berezhnoy, A. Identifying high-affinity aptamer ligands with defined cross-reactivity using high-throughput guided systematic evolution of ligands by exponential enrichment. Nucleic Acids Res. 2015, 43, e82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Yu, H.; Alkhamis, O.; Liu, Y.; Canoura, J.; Fu, F.; Xiao, Y. In vitro isolation of class-specific oligonucleotide-based small-molecule receptors. Nucleic Acids Res. 2019, 47, e71. [Google Scholar] [CrossRef]
- Dua, P.; Kang, S.; Shin, H.-S.; Kim, S.; Lee, D.-K. Cell-SELEX-Based Identification of a Human and Mouse Cross-Reactive Endothelial Cell-Internalizing Aptamer. Nucleic Acid Ther. 2018, 28, 262–271. [Google Scholar] [CrossRef]
- Villarreal, M.A.; Biediger, N.M.; Bonner, N.A.; Miller, J.N.; Zepeda, S.K.; Ricard, B.J.; García, D.M.; Lewis, K.A. Determining Zebrafish Epitope Reactivity to Commercially Available Antibodies. Zebrafish 2017, 14, 387–389. [Google Scholar] [CrossRef]
- Miyazawa, R.; Matsuura, Y.; Shibasaki, Y.; Imamura, S.; Nakanishi, T. Cross-reactivity of monoclonal antibodies against CD4-1 and CD8α of ginbuna crucian carp with lymphocytes of zebrafish and other cyprinid species. Dev. Comp. Immunol. 2018, 80, 15–23. [Google Scholar] [CrossRef]
- Zavyalova, E.; Golovin, A.; Timoshenko, T.; Babiy, A.; Pavlova, G.; Kopylov, A. DNA aptamers for human thrombin with high anticoagulant activity demonstrate target- and species-specificity. Curr. Med. Chem. 2012, 19, 5232–5237. [Google Scholar] [CrossRef]
- Shigdar, S.; Qiao, L.; Zhou, S.F.; Xiang, D.X.; Wang, T.; Li, Y.; Lim, L.Y.; Kong, L.X.; Li, L.H.; Duan, W. RNA aptamers targeting cancer stem cell marker CD133. Cancer Lett. 2013, 330, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Sefah, K.; Shangguan, D.; Xiong, X.; O’Donoghue, M.B.; Tan, W. Development of DNA aptamers using cell-SELEX. Nat. Protoc. 2010, 5, 1169. [Google Scholar] [CrossRef]
- Nagarkatti, R.; Bist, V.; Sun, S.; Fortes de Araujo, F.; Nakhasi, H.L.; Debrabant, A. Development of an Aptamer-Based Concentration Method for the Detection of Trypanosoma cruzi in Blood. PLoS ONE 2012, 7, e43533. [Google Scholar] [CrossRef] [PubMed]
- Stark, Y.; Venet, S.; Schmid, A. Whole Cell Panning with Phage Display. In Synthetic Antibodies: Methods and Protocols, Tiller, T., Ed.; Springer: New York, NY, USA, 2017; pp. 67–91. [Google Scholar]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. SELEX—A (r)evolutionary method to generate high-affinity nucleic acid ligands. Biomol. Eng. 2007, 24, 381–403. [Google Scholar] [CrossRef] [PubMed]
- Lykkemark, S.; Mandrup, O.A.; Jensen, M.B.; Just, J.; Kristensen, P. A novel excision selection method for isolation of antibodies binding antigens expressed specifically by rare cells in tissue sections. Nucleic Acids Res. 2017, 45, e107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Li, X.; Volk, D.E.; Lokesh, G.L.R.; Elizondo-Riojas, M.-A.; Li, L.; Nick, A.M.; Sood, A.K.; Rosenblatt, K.P.; Gorenstein, D.G. Morph-X-Select: Morphology-based tissue aptamer selection for ovarian cancer biomarker discovery. Biotechniques 2016, 61, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Xu, H.; Ding, H.; Huang, Y.; Cao, X.; Yang, G.; Li, J.; Xie, Z.; Meng, Y.; Li, X.; et al. Identification of an aptamer targeting hnRNP A1 by tissue slide-based SELEX. J. Pathol. 2009, 218, 327–336. [Google Scholar] [CrossRef]
- Könning, D.; Kolmar, H. Beyond antibody engineering: Directed evolution of alternative binding scaffolds and enzymes using yeast surface display. Microb. Cell Fact. 2018, 17, 32. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Q.; Sun, Z.; Wang, Y.; Su, B.; Zhang, C.; Cao, H.; Liu, X. Nanobody affinity improvement: Directed evolution of the anti-ochratoxin A single domain antibody. Int. J. Biol. Macromol. 2020, 151, 312–321. [Google Scholar] [CrossRef]
- Sánchez-Martín, D.; Sanz, L.; Ruoslahti, E.; Alvarez-Vallina, L. In vivo selection of tumor-specific antibodies. Oncotarget 2013, 4, 1547. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Chen, Y.H.; Lennox, K.A.; Behlke, M.A.; Davidson, B.L. In vivo SELEX for Identification of Brain-penetrating Aptamers. Mol. Ther. Nucleic Acids 2013, 2, e67. [Google Scholar] [CrossRef]
- Gordon, C.K.L.; Eisenstein, M.; Soh, H.T. Direct Selection Strategy for Isolating Aptamers with pH-Sensitive Binding Activity. ACS Sens. 2018, 3, 2574–2580. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, V.B.; Taylor, A.I.; Cozens, C.; Abramov, M.; Renders, M.; Zhang, S.; Chaput, J.C.; Wengel, J.; Peak-Chew, S.-Y.; McLaughlin, S.H.; et al. Synthetic genetic polymers capable of heredity and evolution. Science 2012, 336, 341–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eremeeva, E.; Fikatas, A.; Margamuljana, L.; Abramov, M.; Schols, D.; Groaz, E.; Herdewijn, P. Highly stable hexitol based XNA aptamers targeting the vascular endothelial growth factor. Nucleic Acids Res. 2019, 47, 4927–4939. [Google Scholar] [CrossRef] [PubMed]
- Beghein, E.; Gettemans, J. Nanobody Technology: A Versatile Toolkit for Microscopic Imaging, Protein–Protein Interaction Analysis, and Protein Function Exploration. Front. Immunol. 2017, 8, 771. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, A.; Kalra, P.; Bansal, V.; Bruno, J.G.; Sharma, T.K. Aptamer-based point-of-care diagnostic platforms. Sens. Actuators B Chem. 2017, 246, 535–553. [Google Scholar] [CrossRef]
- Vidic, J.; Manzano, M.; Chang, C.-M.; Jaffrezic-Renault, N. Advanced biosensors for detection of pathogens related to livestock and poultry. Vet. Res. 2017, 48, 11. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Wu, J.; Gu, J.; Shen, L.; Mao, L. Application of Aptamers in Virus Detection and Antiviral Therapy. Front. Microbiol. 2019, 10, 1462. [Google Scholar] [CrossRef] [Green Version]
- Henri, J.L.; Bayat, N.; Macdonald, J.; Shigdar, S. A guide to using aptamers in cell based assays. Aptamers 2019, 3, 4–9. [Google Scholar]
- Zhang, W.; Liu, Q.X.; Guo, Z.H.; Lin, J.S. Practical Application of Aptamer-Based Biosensors in Detection of Low Molecular Weight Pollutants in Water Sources. Molecules 2018, 23, 344. [Google Scholar] [CrossRef] [Green Version]
- Mehlhorn, A.; Rahimi, P.; Joseph, Y. Aptamer-Based Biosensors for Antibiotic Detection: A Review. Biosensors 2018, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, I.; Sharma, N.; Vasilescu, A.; Iancu, M.; Badea, G.; Boukherroub, R.; Ogale, S.; Szunerits, S. Electrochemical Aptamer-Based Biosensors for the Detection of Cardiac Biomarkers. ACS Omega 2018, 3, 12010–12018. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Zhang, F.; Goldys, E.M.; Gao, F.; Liu, G. Advances in structure-switching aptasensing towards real time detection of cytokines. TrAC Trends Anal. Chem. 2018, 102, 379–396. [Google Scholar] [CrossRef]
- Citartan, M.; Tang, T.-H. Recent developments of aptasensors expedient for point-of-care (POC) diagnostics. Talanta 2019, 199, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.N.; Song, E. Lab-on-a-Chip Systems for Aptamer-Based Biosensing. Micromachines 2020, 11, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Wang, R.; Luo, F.; Wang, P.; Lin, Z. Miniaturized electrochemical sensors and their point-of-care applications. Chin. Chem. Lett. 2020, 31, 589–600. [Google Scholar] [CrossRef]
Figure 1.
Current representation of the Tree of Life from Open Tree of Life (opentreeoflife.org). Of the more than two million known species on this planet, only ≈70,000 are vertebrates. Only a limited number of these have genomes sequenced and fewer have information regarding their proteomes. A high resolution image of this figure can be found at http://www.onezoom.org/life.html/@biota=93302#x1033,y1463,w1.9286.
Figure 1.
Current representation of the Tree of Life from Open Tree of Life (opentreeoflife.org). Of the more than two million known species on this planet, only ≈70,000 are vertebrates. Only a limited number of these have genomes sequenced and fewer have information regarding their proteomes. A high resolution image of this figure can be found at http://www.onezoom.org/life.html/@biota=93302#x1033,y1463,w1.9286.

Figure 2.
Binding of antibody, nanobody, and aptamer to a protein, highlighting the size differences between the three potential binding reagents. In proteins where the homologous region (highlighted in red) between species is small, only part of the antibody may fit, leading to poor species cross-reactivity. The nanobody and aptamer are 10 and 20 times smaller, respectively, and are more likely to cross-react between species.
Figure 2.
Binding of antibody, nanobody, and aptamer to a protein, highlighting the size differences between the three potential binding reagents. In proteins where the homologous region (highlighted in red) between species is small, only part of the antibody may fit, leading to poor species cross-reactivity. The nanobody and aptamer are 10 and 20 times smaller, respectively, and are more likely to cross-react between species.

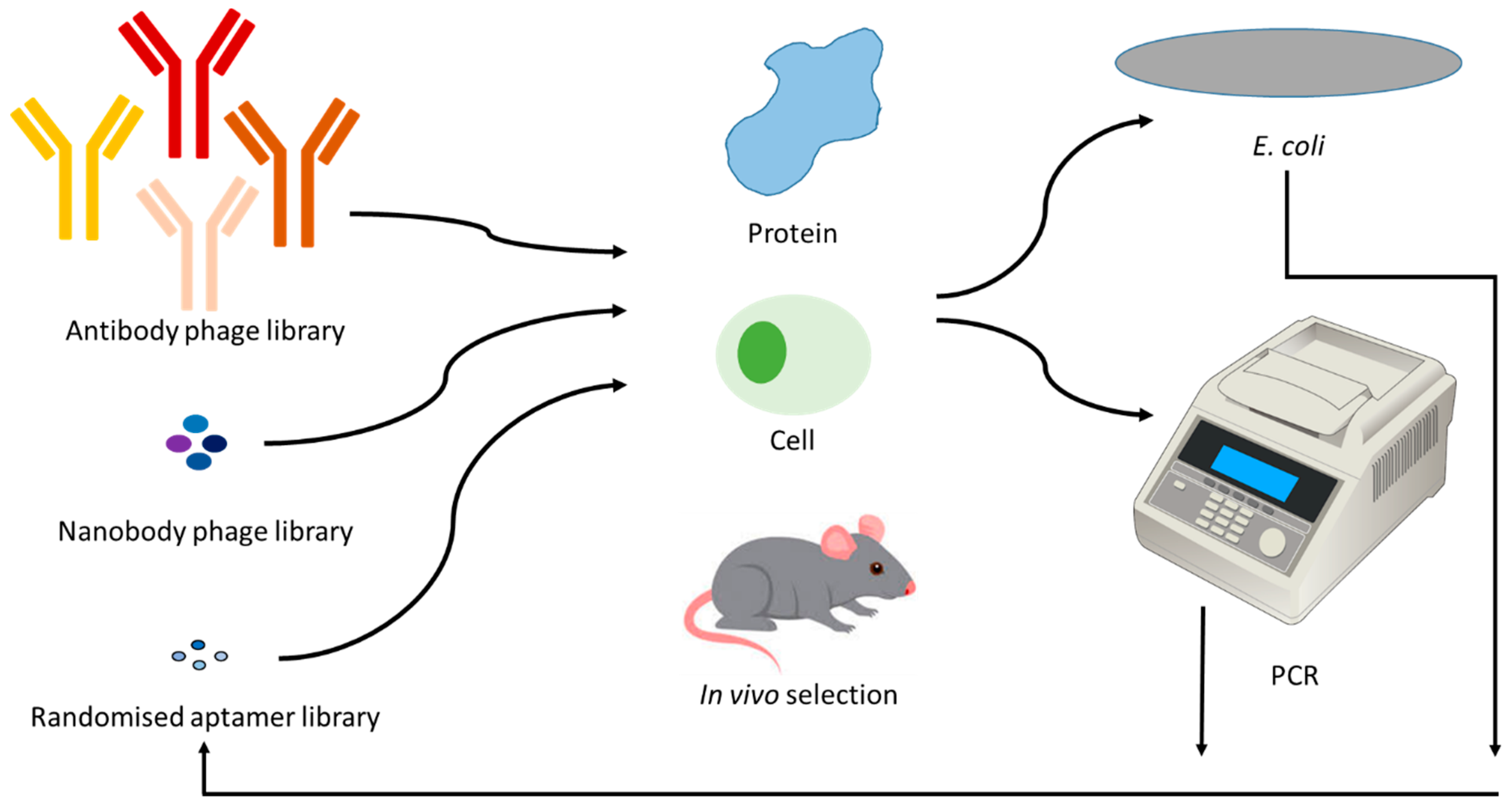
Figure 3.
Diagrammatic representation of the process involved in the generation of specific binding reagents. Combinatorial phage libraries of antibodies or nanobodies, or combinatorial randomized sequences of aptamers are incubated with a target, followed by removal of unbound species via washing. The bound sequences are propagated in Escherichia coli or via PCR. Re-incubation of these enriched species is continued for 3–7 iterative rounds.
Figure 3.
Diagrammatic representation of the process involved in the generation of specific binding reagents. Combinatorial phage libraries of antibodies or nanobodies, or combinatorial randomized sequences of aptamers are incubated with a target, followed by removal of unbound species via washing. The bound sequences are propagated in Escherichia coli or via PCR. Re-incubation of these enriched species is continued for 3–7 iterative rounds.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Studies that have compared species’ proteomes.
| Tested Tissue/Cells/ Protein | Species | Technique | Total No of Similar Proteins between Species | Reference |
|---|---|---|---|---|
| Kidney cortical transporters | Human, monkey, dog, rat, and mouse | LC–MS/MS | 19 | [40] |
| Liver and kidney efflux drug transporters | Human, monkey, rat, dog | Isotope dilution nano LC-MS/MS | 4 specific transporters: MDR1/P-gp, BCRP, MRP2 and MRP3 | [41] |
| Liver microsomes | Human, rat, mouse | 2D-(SCX-RP)-LC–MS/MS | 704 | [42] |
| Milk casein micelles | Holstein cows, buffaloes, Jersey cows, yaks, goats, camels, and horses | LC-MS/MS | 25 | [43] |
| Milk fat globule membrane | Human, cow, goat and yak | LC–MS/MS | 50 | [44] |
| Milk fat globule membrane | Humans, Holstein and Jersey cows, buffaloes, yaks, goats, camels, horses | LC–MS/MS | 399 | [45] |
| Pancreatic beta cells | Human and rat | label-free LC-MS/MS | 185 | [46] |
| Pancreatic cells | Mouse, rat and human | SDS-PAGE (gel) coupled with LC-MS/MS | [47] | |
| Platelets | Human, Rat | SDS-PAGE–LC MS/MS | 837 | [48] |
| Saliva | Human, Dog, Glires, Sheep, cattle, horses | LC–MALDI, SDS-PAGE–LC MS/MS | 13 | [49] |
| Seminal plasma | Alpaca, cattle, horse, sheep, pig, goat and camel | SDS-PAGE LC–MS/MS and 2D–LC–MS/MS | 302 | [50] |
| Sperm | Rodents and ungulates | LC-MS/MS | 623 | [51] |
BCRP: breast cancer resistance protein; LC-MS/MS: liquid chromatography with tandem mass spectrometry; MALDI: matrix-assisted laser desorption/ionization; MDR: multidrug resistance protein; MRP: multidrug resistance-associated protein; P-gp: P-glycoprotein; SCX-RP: strong cation exchange reverse phase; SDS-PAGE: sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
Table 2.
Pros and cons of using antibodies, nanobodies, or aptamers in research.
| Conditions | Antibodies | Nanobodies | Aptamers |
|---|---|---|---|
| Use in physiological conditions (pH, temp, etc) | ✓ | ✓ | ✓ |
| Use in non-physiological conditions | X | X | ✓ |
| Complex target selection | ✓ | ✓ | ✓ |
| Stability in wide temperature range | X | X | ✓ |
| Immunogenicity | ✓ | ✓ | X/limited |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dhar, P.; Samarasinghe, R.M.; Shigdar, S. Antibodies, Nanobodies, or Aptamers—Which Is Best for Deciphering the Proteomes of Non-Model Species? Int. J. Mol. Sci. 2020, 21, 2485. https://doi.org/10.3390/ijms21072485
AMA Style
Dhar P, Samarasinghe RM, Shigdar S. Antibodies, Nanobodies, or Aptamers—Which Is Best for Deciphering the Proteomes of Non-Model Species? International Journal of Molecular Sciences. 2020; 21(7):2485. https://doi.org/10.3390/ijms21072485
Chicago/Turabian StyleDhar, Poshmaal, Rasika M. Samarasinghe, and Sarah Shigdar. 2020. "Antibodies, Nanobodies, or Aptamers—Which Is Best for Deciphering the Proteomes of Non-Model Species?" International Journal of Molecular Sciences 21, no. 7: 2485. https://doi.org/10.3390/ijms21072485
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.