IL10 Alters Peri-Collateral Macrophage Polarization and Hind-Limb Reperfusion in Mice after Femoral Artery Ligation

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Modulation of Blood Concentration Levels of IL10 after Pharmacological Stimulation with IL10 and Anti-IL10
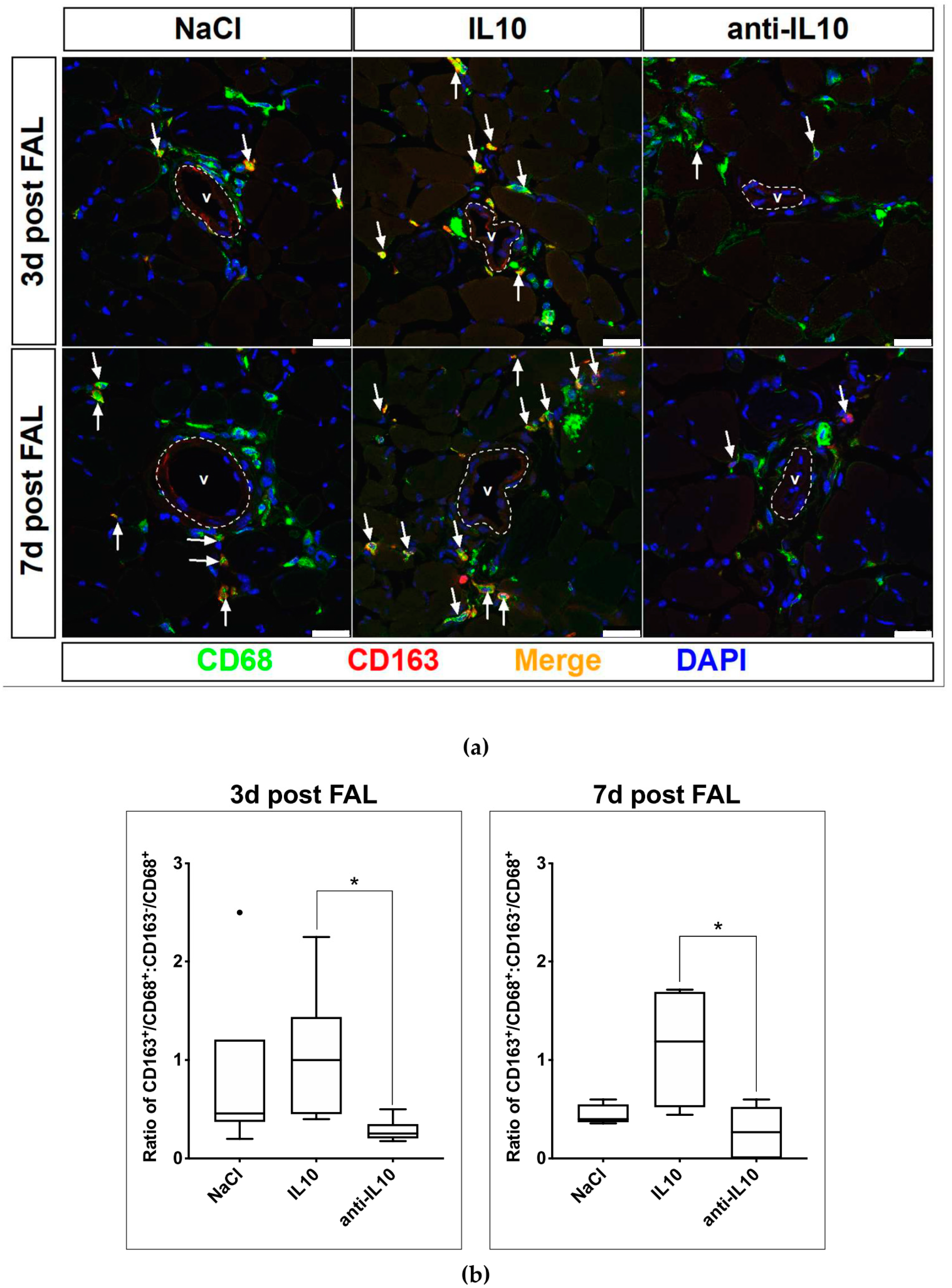
2.2. Alteration of Macrophage Polarization in the Perivascular Bed of Growing Collateral Vessels after Pharmacological Stimulation with IL10 and Anti-IL10
2.3. Evaluation of Hind-Limb Perfusion Recovery after FAL and Pharmacological Stimulation with IL10 and Anti-IL10
2.4. Macroscopic Observations on Ligated Hind-Limbs after FAL and Application of IL10 and Anti-IL10
3. Discussion
4. Materials and Methods
4.1. Animal Models
4.2. Mouse Model of Hind-Limb Ischemia
4.3. Pharmacological Stimulation
4.4. Measurement of Blood Concentration Levels of IL10
4.5. Hind-limb Perfusion Measurement after Pharmacological Stimulation and FAL
4.6. Immunohistochemistry
4.7. Data Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FAL | Femoral artery ligation |
| CIE | Critical ischemic events |
| VSMC | Vascular smooth muscle cells |
| SMC | Smooth muscle cells |
| ECM | Extracellular matrix |
References
- Heil, M.; Schaper, W. Influence of mechanical, cellular, and molecular factors on collateral artery growth (arteriogenesis). Circ. Res. 2004, 95, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, W.D.; Arras, M.; Winkler, B.; Scholz, D.; Schaper, J.; Schaper, W. Monocyte chemotactic protein-1 increases collateral and peripheral conductance after femoral artery occlusion. Circ. Res. 1997, 80, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, I.R.; Hoefer, I.E.; Van Royen, N.; Katzer, E.; Braun-Dulleaus, R.; Heil, M.; Kostin, S.; Bode, C.; Schaper, W. GM-CSF: A strong arteriogenic factor acting by amplification of monocyte function. Atherosclerosis 2001, 159, 343–356. [Google Scholar] [CrossRef]
- Ziegelhoeffer, T.; Fernandez, B.; Kostin, S.; Heil, M.; Voswinckel, R.; Helisch, A.; Schaper, W. Bone marrow-derived cells do not incorporate into the adult growing vasculature. Circ. Res. 2004, 94, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khmelewski, E.; Becker, A.; Meinertz, T.; Ito, W.D. Tissue resident cells play a dominant role in arteriogenesis and concomitant macrophage accumulation. Circ. Res. 2004, 95, e56–e64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jetten, N.; Donners, M.M.P.C.; Wagenaar, A.; Cleutjens, J.P.M.; Van Rooijen, N.; De Winther, M.P.J.; Post, M.J. Local delivery of polarized macrophages improves reperfusion recovery in a mouse hind limb ischemia model. PLoS ONE 2013, 8, e68811. [Google Scholar] [CrossRef]
- Takeda, Y.; Costa, S.; Delamarre, E.; Roncal, C.; Leite De Oliveira, R.; Squadrito, M.L.; Finisguerra, V.; Deschoemaeker, S.; Bruyere, F.; Wenes, M.; et al. Macrophage skewing by Phd2 haplodeficiency prevents ischaemia by inducing arteriogenesis. Nature 2011, 479, 122–126. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Mills, C.D.; Ley, K. M1 and M2 macrophages: The chicken and the egg of immunity. J. Innate Immun. 2014, 6, 716–726. [Google Scholar] [CrossRef]
- Troidl, C.; Jung, G.; Troidl, K.; Hoffmann, J.; Mollmann, H.; Nef, H.; Schaper, W.; Hamm, C.W.; Schmitz-Rixen, T. The temporal and spatial distribution of macrophage subpopulations during arteriogenesis. Curr. Vasc. Pharmacol. 2013, 11, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Italiani, P.; Boraschi, D. From monocytes to M1/M2 macrophages: Phenotypical vs. functional differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komohara, Y.; Jinushi, M.; Takeya, M. Clinical significance of macrophage heterogeneity in human malignant tumors. Cancer Sci. 2014, 105, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaper, W.; Ito Wulf, D. Molecular mechanisms of coronary collateral vessel growth. Circ. Res. 1996, 79, 911–919. [Google Scholar] [CrossRef]
- Berk, B.C. Vascular smooth muscle growth: Autocrine growth mechanisms. Physiol. Rev. 2001, 81, 999–1030. [Google Scholar] [CrossRef] [PubMed]
- Schaper, W. Collateral circulation: Past and present. Basic Res. Cardiol. 2009, 104, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Micklem, K.; Rigney, E.; Cordell, J.; Simmons, D.; Stross, P.; Turley, H.; Seed, B.; Mason, D. A human macrophage-associated antigen (CD68) detected by six different monoclonal antibodies. Br. J. Haematol. 1989, 73, 6–11. [Google Scholar] [CrossRef]
- Buechler, C.; Ritter, M.; Orsó, E.; Langmann, T.; Klucken, J.; Schmitz, G. Regulation of scavenger receptor CD163 expression in human monocytes and macrophages by pro-and antiinflammatory stimuli. J. Leukoc. Biol. 2000, 67, 97–103. [Google Scholar] [CrossRef]
- O’rourke, S.A.; Dunne, A.; Monaghan, M.G. The Role of macrophages in the infarcted myocardium: Orchestrators of ECM remodeling. Front. Cardiovasc. Med. 2019, 6, 101. [Google Scholar] [CrossRef] [Green Version]
- Royen, N.V.; Hoefer, I.; Buschmann, I.; Heil, M.; Kostin, S.; Deindl, E.; Vogel, S.; Korff, T.; Augustin, H.; Bode, C.; et al. Exogenous application of transforming growth factor beta 1 stimulates arteriogenesis in the peripheral circulation. FASEB J. 2002, 16, 432–434. [Google Scholar] [CrossRef]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The role of macrophages in acute and chronic wound healing and interventions to promote pro-wound healing phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.-J.; Koltai, S.; Kocsis, E.; Scholz, D.; Kostin, S.; Luo, X.; Schaper, W.; Schaper, J. Remodeling of the adventitia during coronary arteriogenesis. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H31–H40. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Akahori, H.; Harari, E.; Smith, S.L.; Polavarapu, R.; Karmali, V.; Otsuka, F.; Gannon, R.L.; Braumann, R.E.; Dickinson, M.H.; et al. CD163+ macrophages promote angiogenesis and vascular permeability accompanied by inflammation in atherosclerosis. J. Clin. Investig. 2018, 128, 1106–1124. [Google Scholar] [CrossRef] [PubMed]
- Limbourg, A.; Korff, T.; Napp, L.C.; Schaper, W.; Drexler, H.; Limbourg, F.P. Evaluation of postnatal arteriogenesis and angiogenesis in a mouse model of hind-limb ischemia. Nat. Protoc. 2009, 4, 1737–1748. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Ziegelhoeffer, T.; Pipp, F.; Kostin, S.; Martin, S.; Clauss, M.; Schaper, W. Blood monocyte concentration is critical for enhancement of collateral artery growth. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2411–H2419. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Götze, A.M.; Schubert, C.; Jung, G.; Dörr, O.; Liebetrau, C.; Hamm, C.W.; Schmitz-Rixen, T.; Troidl, C.; Troidl, K. IL10 Alters Peri-Collateral Macrophage Polarization and Hind-Limb Reperfusion in Mice after Femoral Artery Ligation. Int. J. Mol. Sci. 2020, 21, 2821. https://doi.org/10.3390/ijms21082821
Götze AM, Schubert C, Jung G, Dörr O, Liebetrau C, Hamm CW, Schmitz-Rixen T, Troidl C, Troidl K. IL10 Alters Peri-Collateral Macrophage Polarization and Hind-Limb Reperfusion in Mice after Femoral Artery Ligation. International Journal of Molecular Sciences. 2020; 21(8):2821. https://doi.org/10.3390/ijms21082821
Chicago/Turabian StyleGötze, Alexander M., Christian Schubert, Georg Jung, Oliver Dörr, Christoph Liebetrau, Christian W. Hamm, Thomas Schmitz-Rixen, Christian Troidl, and Kerstin Troidl. 2020. "IL10 Alters Peri-Collateral Macrophage Polarization and Hind-Limb Reperfusion in Mice after Femoral Artery Ligation" International Journal of Molecular Sciences 21, no. 8: 2821. https://doi.org/10.3390/ijms21082821