
Rac1/Wave2/Arp3 Pathway Mediates Rat Blood-Brain Barrier Dysfunction under Simulated Microgravity Based on Proteomics Strategy



Abstract
:
1. Introduction
2. Results
2.1. 21-Day SMG (21d-SMG) Damaged Brain Histomorphology and BBB Ultrastructure
2.2. 21d-SMG Induced BBB Hyperpermeability in Rats
2.3. 21d-SMG Increased Oxidative Stress Levels and Proinflammatory Cytokine Levels in the Rat Brain
2.4. SMG Downregulated TJs and AJs Proteins in the Rat Brain
2.5. Proteomics Analysis
2.5.1. GO Analysis
2.5.2. KEGG Pathway Analysis
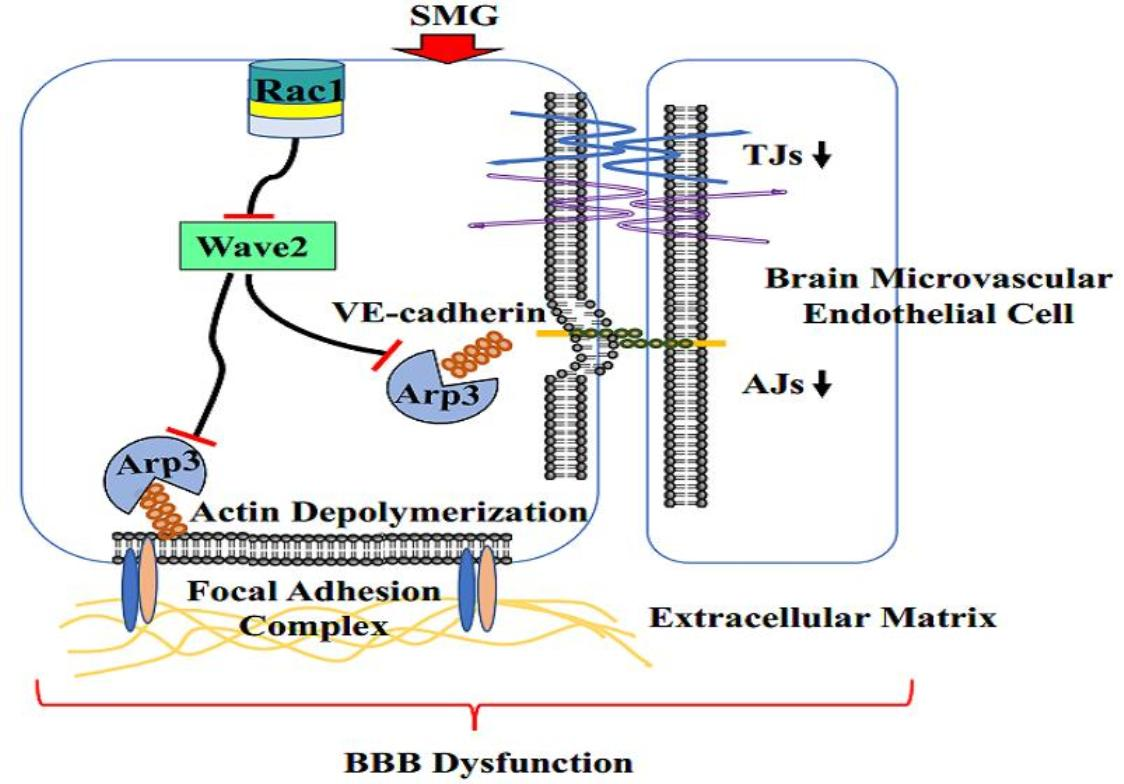
2.6. SMG Inhibited the Rac1/Wave2/Arp3 Pathway
2.7. SMG Induced Apoptosis and Oxidative Stress Injury and F-Actin Distribution in HBMECs
2.8. SMG Downregulated AJs and TJs Protein Expression and the Rac1/Wave2/Arp3 Pathway in HBMECs
3. Discussion
4. Materials and Methods
4.1. Animal Treatment and Sample Collection
4.2. Measurement of BBB Permeability
4.3. Histomorphology and Ultrastructure Observation of Rat Brain
4.3.1. The H&E Staining
4.3.2. Transmission Electron Microscopy (TEM) Observation
4.4. Cell Culture and MG Simulation
Cell Apoptosis Assays and Cell Cycle Analysis
4.5. Detection of Oxidative Stress Levels and Proinflammatory Cytokine Levels in HBMECs and Rat Brain
4.6. Western-Blot Analysis
4.7. Immunohistochemistry (IHC) and Immunofluorescence Staining Analysis
4.8. Proteomics
4.8.1. Protein Extraction and In-Gel Digestion
4.8.2. High-Performance Liquid Chromatography (HPLC)-Tandem Mass Spectrometry (MS/MS) Analysis
4.8.3. Protein Identification and Bioinformatics Analysis
4.9. Rac1, Wave2, and Arp3 Expression and the Ratio of F-Actin to G-Actin
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MG | Microgravity |
| BBB | Blood-brain barrier |
| TS | tail-suspension |
| VE-cadherin | vascular endothelial cadherin |
| FA | focal adhesion complex |
| Dex | texas red-Dextran |
| KEGG | kyoto encyclopedia of genes and genomes |
| Arp3 | actin-related protein-3 |
| HPLC | high performance liquid chromatography |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| HBMECs | human brain microvascular endothelial cell |
| Wave2 | the Wiskott–Aldrich syndrome protein family verprolin-homologous protein-2 |
| SMG | simulated microgravity |
| CNS | central nervous system |
| AJs | adherens junctions |
| TJs | tight junctions |
| ZO | zonula occludens |
| DEPs | differentially expressed proteins |
| ECM | extracellular matrix |
| EB | Evans blue |
| GO | gene-ontology |
| F-actin | filamentous-actin |
| G-actin | globular-actin |
References
- Grimm, D.; Grosse, J.; Wehland, M.; Mann, V.; Reseland, J.E.; Sundaresan, A.; Corydon, T.J. The impact of microgravity on bone in humans. Bone 2016, 87, 44–56. [Google Scholar] [CrossRef]
- Jadvar, H. Medical imaging in microgravity. Aviat. Space. Environ. Med. 2000, 71, 640–646. [Google Scholar]
- Demontis, G.C.; Germani, M.M.; Caiani, E.G.; Barravecchia, I.; Passino, C.; Angeloni, D. Human pathophysiological adaptations to the space environment. Front. Physiol. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- Borchers, A.T.; Keen, C.L.; Gershwin, M.E. Microgravity and immune responsiveness: Implications for space travel. Nutrition 2002, 18, 889–898. [Google Scholar] [CrossRef]
- De la Torre, G.G. Cognitive neuroscience in space. Life 2014, 4, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.M.; Chen, H.L.; Lv, K.; Ji, G.H.; Zhang, Y.L.; Wang, Y.L.; Li, Y.H.; Qu, L. iTRAQ-based proteomics analysis of hippocampus in spatial memory deficiency rats induced by simulated microgravity. J. Proteom. 2017, 160, 64–73. [Google Scholar] [CrossRef]
- Graebe, A.; Schuck, E.L.; Lensing, P.; Putcha, L.; Derendorf, H. Physiological, pharmacokinetic, and pharmacodynamic changes in space. J. Clin. Pharmacol. 2004, 44, 837–853. [Google Scholar] [CrossRef] [PubMed]
- Nation, D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A.R.; Buennagel, D.P.; Harrington, M.G.; et al. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 2019, 25, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Muresanu, D.F.; Lafuente, J.V.; Patnaik, R.; Tian, Z.R.; Buzoianu, A.D.; Sharma, H.S. Sleep deprivation-induced blood-brain barrier breakdown and brain dysfunction are exacerbated by size-related exposure to Ag and Cu nanoparticles. neuroprotective effects of a 5-HT3 receptor antagonist ondansetron. Mol. Neurobiol. 2015, 52, 867–881. [Google Scholar] [CrossRef] [PubMed]
- Costea, L.; Mészáros, Á.; Bauer, H.; Bauer, H.C.; Traweger, A.; Wilhelm, I.; Farkas, A.E.; Krizbai, I.A. The blood-brain barrier and its intercellular junctions in age-related brain disorders. Int. J. Mol. Sci. 2019, 20, 5472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pun, P.B.; Lu, J.; Moochhala, S. Involvement of ROS in BBB dysfunction. Free Radic. Res. 2009, 43, 348–364. [Google Scholar] [CrossRef] [PubMed]
- Lajkó, E.; Tuka, B.; Fülöp, F.; Krizbai, I.; Toldi, J.; Magyar, K.; Vécsei, L.; Kőhidai, L. Kynurenic acid and its derivatives are able to modulate the adhesion and locomotion of brain endothelial cells. J. Neural. Transm. 2018, 125, 899–912. [Google Scholar] [CrossRef] [PubMed]
- Persidsky, Y.; Ramirez, S.H.; Haorah, J.; Kanmogne, G.D. Blood-brain barrier: Structural components and function under physiologic and pathologic conditions. J. Neuroimmune. Pharmacol. 2006, 1, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Hartsock, A.; Nelson, W.J. Adherens and tight junctions: Structure, function and connections to the actin cytoskeleton. Biochim. Biophys. Acta. 2008, 1778, 660–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.J.; Wang, H.; Liang, W.Y.; Xu, W.; Li, Y.P.; Song, L.X.; Zhang, D.X.; Hu, Y.M.; Han, B.; Wang, W.X.; et al. Bilobalide reversibly modulates blood-brain barrier permeability through promoting adenosine A1 receptor mediated phosphorylation of actin-binding proteins. Biochem. Biophys. Res. Commun. 2020, 526, 1077–1084. [Google Scholar] [CrossRef]
- Lakin, W.D.; Stevens, S.A.; Penar, P.L. Modeling intracranial pressures in microgravity: The influence of the blood-brain barrier. Aviat. Space. Environ. Med. 2007, 78, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Nishiyama, N.C.; Byrum, S.D.; Stanbouly, S.; Jones, T.; Holley, J.; Sridharan, V.; Boerma, M.; Tackett, A.J.; Willey, J.S.; et al. Spaceflight induces oxidative damage to blood-brain barrier integrity in a mouse model. FASEB J. 2020, 34, 15516–15530. [Google Scholar] [CrossRef] [PubMed]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Khoutorova, L.; Bazan, N.G.; Belayev, L. Docosahexaenoic acid improves behavior and attenuates blood-brain barrier injury induced by focal cerebral ischemia in rats. Exp. Transl. Stroke. Med. 2015, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.Y.; Xu, W.; Zhu, J.; Gu, Q.L.; Li, Y.P.; Guo, C.J.; Huang, Y.J.; Yu, J.F.; Wang, W.X.; Hu, Y.M.; et al. Ginkgo biloba extract improves brain uptake of ginsenosides by increasing blood-brain barrier permeability via activating A1 adenosine receptor signaling pathway. J. Ethnopharmacol. 2020, 246, 112243. [Google Scholar] [CrossRef] [PubMed]
- Legerstee, K.; Geverts, B.; Slotman, J.A.; Houtsmuller, A.B. Dynamics and distribution of paxillin, vinculin, zyxin and VASP depend on focal adhesion location and orientation. Sci. Rep. 2019, 9, 10460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldmann, W.H. Vinculin-p130Cas interaction is critical for focal adhesion dynamics and mechano-transduction. Cell. Biol. Int. 2014, 38, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, M.H. Integrin activation. BMB Rep. 2014, 47, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Bian, C.; Wang, H.; Huang, S.; Wu, D. MiR-203 down-regulates Rap1A and suppresses cell proliferation, adhesion and invasion in prostate cancer. J. Exp. Clin. Cancer. Res. 2015, 34, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadri, S.K. Cross talk between focal adhesion kinase and cadherins: Role in regulating endothelial barrier function. Microvasc. Res. 2012, 83, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu Taha, A.; Taha, M.; Seebach, J.; Schnittler, H.J. ARP2/3-mediated junction-associated lamellipodia control VE-cadherin-based cell junction dynamics and maintain monolayer integrity. Mol. Biol. Cell. 2014, 25, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Abu Taha, A.; Schnittler, H.J. Dynamics between actin and the VE-cadherin/catenin complex: Novel aspects of the ARP2/3 complex in regulation of endothelial junctions. Cell. Adh. Migr. 2014, 8, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shawki, A.; McCole, D.F. Mechanisms of intestinal epithelial barrier dysfunction by adherent-invasive escherichia coli. Cell. Mol. Gastroenterol Hepatol. 2016, 3, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Graziano, B.R.; Town, J.P.; Sitarska, E.; Nagy, T.L.; Fošnarič, M.; Penič, S.; Iglič, A.; Kralj-Iglič, V.; Gov, N.S.; Diz-Muñoz, A.; et al. Cell confinement reveals a branched-actin independent circuit for neutrophil polarity. PLoS Biol. 2019, 17, e3000457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infanger, M.; Ulbrich, C.; Baatout, S.; Wehland, M.; Kreutz, R.; Bauer, J.; Grosse, J.; Vadrucci, S.; Cogoli, A.; Derradji, H.; et al. Inhibition of the Rac1-WAVE2-Arp2/3 signaling pathway promotes radiosensitivity via downregulation of cofilin-1 in U251 human glioma cells. Mol. Med. Rep. 2016, 13, 4414–4420. [Google Scholar]
- Ko, H.S.; Kim, J.S.; Cho, S.M.; Lee, H.J.; Ahn, K.S.; Kim, S.H.; Lee, E.O. Urokinase-type plasminogen activator expression and Rac1/WAVE-2/Arp2/3 pathway are blocked by pterostilbene to suppress cell migration and invasion in MDA-MB-231 cells. Bioorg. Med. Chem. Lett. 2014, 24, 1176–1179. [Google Scholar] [CrossRef] [Green Version]
- Benavides, D.T.; Richard, S.; Tanner, S.; Wyss, F.; Egli, M.; Franco-Obregón, A. Calcium-dependent deceleration of the cell cycle in muscle cells by simulated microgravity. FASEB J. 2013, 27, 2045–2054. [Google Scholar]
- Rahman, M.T.; Ghosh, C.; Hossain, M.; Linfield, D.; Rezaee, F.; Janigro, D.; Marchi, N.; van Boxel-Dezaire, A.H.H. IFN-γ, IL-17A, or zonulin rapidly increase the permeability of the blood-brain and small intestinal epithelial barriers: Relevance for neuro-inflammatory diseases. Biochem. Biophys. Res. Commun. 2018, 507, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.Y.; Zhang, Z.; Liu, Y.; Tang, X.L.; Shu, S.; Bao, X.Y.; Zhang, Y.; Gu, Y.; Xu, Y.; Cao, X. TMEM16A inhibition preserves blood-Brain barrier integrity after ischemic stroke. Front. Cell. Neurosci. 2019, 13, 360. [Google Scholar] [CrossRef] [Green Version]
- Vandenhaute, E.; Culot, M.; Gosselet, F.; Dehouck, L.; Godfraind, C.; Franck, M.; Plouët, J.; Cecchelli, R.; Dehouck, M.P.; Ruchoux, M.M. Brain pericytes from stress-susceptible pigs increase blood-brain barrier permeability in vitro. Fluids. Barriers. CNS. 2012, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbuzova-Davis, S.; Rodrigues, M.C.; Hernandez-Ontiveros, D.G.; Tajiri, N.; Frisina-Deyo, A.; Boffeli, S.M.; Abraham, J.V.; Pabon, M.; Wagner, A.; Ishikawa, H.; et al. Blood-brain barrier alterations provide evidence of subacute diaschisis in an ischemic stroke rat model. PLoS ONE 2013, 8, e63553. [Google Scholar] [CrossRef] [PubMed]
- El-Sokkary, G.H.; Nafady, A.A.; Shabash, E.H. Melatonin administration ameliorates cadmium-induced oxidative stress and morphological changes in the liver of rat. Ecotoxicol. Environ. Saf. 2010, 73, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Małkiewicz, M.A.; Szarmach, A.; Sabisz, A.; Cubała, W.J.; Szurowska, E.; Winklewski, P.J. Blood-brain barrier permeability and physical exercise. J. Neuroinflamm. 2019, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Steller, J.G.; Alberts, J.R.; Ronca, A.E. Oxidative stress as cause, consequence, or biomarker of altered female reproduction and development in the space environment. Int. J. Mol. Sci. 2018, 19, 3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.W.; Sandberg, L.B.; Gridley, D.S.; Herrmann, E.C.; Zhang, G.; Raghavan, R.; Zubarev, R.A.; Zhang, B.; Stodieck, L.S.; Ferguson, V.L.; et al. Proteomic analysis of mouse brain subjected to spaceflight. Int. J. Mol. Sci. 2018, 20, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Javed, I.; Liu, Y.H.; Lu, S.; Peng, G.; Zhang, Y.Q.; Qing, H.; Deng, Y.L. Effect of prolonged simulated microgravity on metabolic proteins in rat hippocampus: Steps toward safe space travel. J. Proteome. Res. 2016, 15, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Nishiyama, N.C.; Campbell-Beachler, M.; Gifford, P.; Haynes, K.E.; Gridley, D.S.; Pecaut, M.J. Role of NADPH oxidase as a mediator of oxidative damage in low-dose irradiated and hindlimb-unloaded mice. Radiat. Res. 2017, 188, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Katsu, M.; Niizuma, K.; Yoshioka, H.; Okami, N.; Sakata, H.; Chan, P.H. Hemoglobin-induced oxidative stress contributes to matrix metalloproteinase activation and blood-brain barrier dysfunction in vivo. J. Cereb. Blood Flow Metab. 2010, 30, 1939–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Jin, Y.P. Toxic effects of combined treatment of 1,2-dichloroethane and ethanol on mouse brain and the related mechanisms. J. Biochem. Mol. Toxicol. 2019, 33, e22294. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.Y.; Wang, S.J.; Yao, G.T.; Liu, Z.G.; Xiao, N. The protective effect and its mechanism of 3-n-butylphthalide pretreatment on cerebral ischemia reperfusion injury in rats. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5275–5282. [Google Scholar] [PubMed]
- Jing, L.; Wu, N.; He, L.; Shao, J.; Ma, H. Establishment of an experimental rat model of high altitude cerebral edema by hypobaric hypoxia combined with temperature fluctuation. Brain. Res. Bull. 2020, 165, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.J.; Hu, W.; Yang, Z.; Li, T.; Jiang, S.; Ma, Z.Q.; Chen, F.L.; Yang, Y. Focusing on claudin-5: A promising candidate in the regulation of BBB to treat ischemic stroke. Prog. Neurobiol. 2018, 161, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Keaney, J.; Walsh, D.M.; O’Malley, T.; Hudson, N.; Crosbie, D.E.; Loftus, T.; Sheehan, F.; McDaid, J.; Humphries, M.M.; Callanan, J.J.; et al. Autoregulated paracellular clearance of amyloid-β across the blood-brain barrier. Sci. Adv. 2015, 1, e1500472. [Google Scholar] [CrossRef] [Green Version]
- Menard, C.; Pfau, M.L.; Hodes, G.E.; Kana, V.; Wang, V.X.; Bouchard, S.; Takahashi, A.; Flanigan, M.E.; Aleyasin, H.; LeClair, K.B.; et al. Social stress induces neurovascular pathology promoting depression. Nat. Neurosci. 2017, 20, 1752–1760. [Google Scholar] [CrossRef] [Green Version]
- Sonar, S.A.; Shaikh, S.; Joshi, N.; Atre, A.N.; Lal, G. IFN-γ promotes transendothelial migration of CD4+ T cells across the blood-brain barrier. Immunol. Cell. Biol. 2017, 95, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.M.; Gu, C.P.; Mandeville, E.T.; Dong, Y.L.; Esposito, E.; Zhang, Y.Y.; Yang, G.; Shen, Y.; Fu, X.; Lo, E.H.; et al. Anesthesia and surgery impair blood-brain barrier and cognitive function in mice. Front. Immunol. 2017, 8, 902. [Google Scholar] [CrossRef] [PubMed]
- Austen, K.; Ringer, P.; Mehlich, A.; Chrostek-Grashoff, A.; Kluger, C.; Klingner, C.; Sabass, B.; Zent, R.; Rief, M.; Grashoff, C. Extracellular rigidity sensing by talin isoform-specific mechanical linkages. Nat. Cell Biol. 2015, 17, 1597–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izard, T.; Brown, D.T. Mechanisms and functions of vinculin interactions with phospholipids at cell adhesion sites. J. Biol. Chem. 2016, 291, 2548–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldmann, W.H. Role of vinculin in cellular mechanotransduction. Cell Biol. Int. 2016, 40, 241–256. [Google Scholar] [CrossRef] [PubMed]
- DeMali, K.A.; Barlow, C.A.; Burridge, K. Recruitment of the Arp2/3 complex to vinculin: Coupling membrane protrusion to matrix adhesion. J. Cell Biol. 2002, 159, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Zaidel-Bar, R.; Ballestrem, C.; Kam, Z.; Geiger, B. Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. J. Cell Sci. 2003, 116, 4605–4613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamir, E.; Katz, B.Z.; Aota, S.; Yamada, K.M.; Geiger, B.; Kam, Z. Molecular diversity of cell-matrix adhesions. J. Cell Sci. 1999, 112, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Zimerman, B.; Volberg, T.; Geiger, B. Early molecular events in the assembly of the focal adhesion-stress fiber complex during fibroblast spreading. Cell Motil. Cytoskelet. 2004, 58, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ramirez, M.A.; Wu, D.; Pryce, G.; Simpson, J.E.; Reijerkerk, A.; King-Robson, J.; Kay, O.; de Vries, H.E.; Hirst, M.C.; Sharrack, B.; et al. MicroRNA-155 negatively affects blood-brain barrier function during neuroinflammation. FASEB J. 2014, 28, 2551–2565. [Google Scholar] [CrossRef] [PubMed]
- Nassef, M.Z.; Kopp, S.; Wehland, M.; Melnik, D.; Sahana, J.; Krüger, M.; Corydon, T.J.; Oltmann, H.; Schmitz, B.; Schütte, A.; et al. Real microgravity influences the cytoskeleton and focal adhesions in human breast cancer cells. Int. J. Mol. Sci. 2019, 20, 3156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayo, A.; Parsons, M. Fascin: A key regulator of cytoskeletal dynamics. Int. J. Biochem. Cell Biol. 2010, 42, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Dawson, J.C.; Forero-Vargas, M.; Spence, H.J.; Yu, X.; König, I.; Anderson, K.; Machesky, L.M. The actin-bundling protein fascin stabilizes actin in invadopodia and potentiates protrusive invasion. Curr. Biol. 2010, 20, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, Y.; Parsons, M.; Adams, J.C. Dual actin-bundling and protein kinase C-binding activities of fascin regulate carcinoma cell migration downstream of Rac and contribute to metastasis. Mol. Biol. Cell 2007, 18, 4591–4602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncharuk, V.N.; Ross, J.S.; Carlson, J.A. Actin-binding protein fascin expression in skin neoplasia. J. Cutan. Pathol. 2002, 29, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.L. Linking Rap to cell adhesion. Curr. Opin. Cell Biol. 2005, 17, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Belvitch, P.; Htwe, Y.M.; Brown, M.E. Dudek S cortical actin dynamics in endothelial permeability. Curr. Top. Membr. 2018, 82, 141–195. [Google Scholar] [PubMed]
- Song, H.; Zhang, J.; He, W.; Wang, P.; Wang, F. Activation of cofilin increases intestinal permeability via depolymerization of F-Actin during hypoxia in vitro. Front. Physiol. 2019, 10, 1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infanger, M.; Ulbrich, C.; Baatout, S.; Wehland, M.; Kreutz, R.; Bauer, J.; Grosse, J.; Vadrucci, S.; Cogoli, A.; Derradji, H.; et al. Modeled gravitational unloading induced downregulation of endothelin-1 in human endothelial cells. J. Cell. Biochem. 2007, 101, 1439–1455. [Google Scholar] [CrossRef]
- Rizzo, A.M.; Montorfano, G.; Negroni, M.; Corsetto, P.; Berselli, P.; Marciani, P.; Zava, S.; Berra, B. Simulated microgravity induce glutathione antioxidant pathway in Xenopus laevis embryos. Cell Biol. Int. 2009, 33, 893–898. [Google Scholar] [CrossRef]
- Tan, X.; Xu, A.; Zhao, T.; Zhao, Q.; Zhang, J.; Fan, C.; Deng, Y.; Freywald, A.; Genth, H.; Xiang, J. Simulated microgravity inhibits cell focal adhesions leading to reduced melanoma cell proliferation and metastasis via FAK/RhoA-regulated mTORC1 and AMPK pathways. Sci. Rep. 2018, 8, 3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatan, M.; Shinder, V.; Israeli, D.; Schnorrer, F.; Volk, T. The Drosophila blood brain barrier is maintained by GPCR-dependent dynamic actin structures. J. Cell Biol. 2011, 192, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Huang, C.J.; Khan, F.A.; Jiao, X.F.; Liu, X.M.; Pandupuspitasari, N.S.; Brohi, R.D.; Huo, L.J. SENP3 grants tight junction integrity and cytoskeleton architecture in mouse Sertoli cells. Oncotarget 2017, 8, 58430–58442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.X.; Chen, Y.H.; Liu, J.F.; Zhang, L.; Zhang, J.; Yang, Y.L.; Lv, Q.; Xie, M.X. Blood-brain barrier disruption induced by diagnostic ultrasound combined with microbubbles in mice. Oncotarget 2017, 9, 4897–4914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.F.; Theofanis, T.N.; Lang, M.J.; Stricsek, G.P.; Lin, R.; Lebrun, A.; Hooper, D.C.; Rosenwasser, R.H.; Sharan, A.D.; Iacovitti, L. Sphenopalatine ganglion stimulation is a reversible and frequency-dependent modulator of the blood-brain barrier. Brain Res. 2019, 1718, 231–241. [Google Scholar] [CrossRef]
- Quynh, C.H.N.; Nghia Son, H.; Chinh Chung, D.; Huan, L.D.; Hong Diem, T.; Long, L.T. Simulated microgravity reduces proliferation and reorganizes the cytoskeleton of human umbilical cord mesenchymal stem cells. Physiol. Res. 2020, 69, 897–906. [Google Scholar]
- Sokolovskaya, A.; Korneeva, E.; Zaichenko, D.; Virus, E.; Kolesov, D.; Moskovtsev, A.; Kubatiev, A. Changes in the surface expression of Intercellular adhesion molecule 3, the induction of apoptosis, and the inhibition of cell-cycle progression of human multidrug-resistant jurkat/A4 cells exposed to a random positioning machine. Int. J. Mol. Sci. 2020, 21, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.B.; Zhang, Y.S.; Guo, J.J.; Kang, L.; Deng, Y.; Li, Y. Investigation on rat intestinal homeostasis alterations induced by 7-day simulated microgravity effect based on a proteomic approach. Acta. Astronaut. 2018, 166, 560–566. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Term | Gene Counts | FE |
|---|---|---|---|
| Annotation Cluster 1 | Enrichment Score: 13.61 | ||
| GOTERM_CC | cell–cell adherent junction | 36 | 4.7 |
| GOTERM_MF | cadherin binding involved in cell–cell adhesion | 35 | 4.9 |
| GOTERM_BP | cell–cell adhesion | 34 | 5.2 |
| Annotation Cluster 2 | Enrichment Score: 11.66 | ||
| GOTERM_MF | GTP binding | 51 | 4.2 |
| GOTERM_MF | GTPase activity | 32 | 5.0 |
| GOTERM_MF | GDP binding | 18 | 9.3 |
| GOTERM_BP | small GTPase mediated signal transduction | 23 | 3.2 |
| Annotation Cluster 3 | Enrichment Score: 7.6 | ||
| GOTERM_CC | proteasome complex | 18 | 10.3 |
| GOTERM_CC | proteasome core complex | 11 | 17.6 |
| GOTERM_MF | threonine-type endopeptidase activity | 11 | 14.3 |
| GOTERM_CC | proteasome core complex, alpha-subunit complex | 5 | 18.7 |
| GOTERM_BP | proteolysis involved in cellular protein catabolic process | 8 | 5.1 |
| Annotation Cluster 4 | Enrichment Score: 5.51 | ||
| GOTERM_BP | translation | 33 | 2.8 |
| GOTERM_CC | cytosolic small ribosomal subunit | 15 | 4.7 |
| GOTERM_MF | structural constituent of ribosome | 35 | 2.6 |
| GOTERM_CC | ribosome | 17 | 3.8 |
| GOTERM_BP | cytosolic large ribosomal subunit | 17 | 3.3 |
| UniProt IDs | Protein Names | Fold Change | p-Value |
|---|---|---|---|
| Q7TP36 | shroom family member 2 (Shroom2) | 12.369 | 0.0017 |
| Q9Z1Z3 | epsin 2 (Epn2) | 9.2441 | 0.0005 |
| O35964 | SH3 domain-containing GRB2-like 1 (Sh3gl1) | 3.4588 | 0.0072 |
| Q5U2U2 | v-crk avian sarcoma virus CT10 oncogene homolog-like (Crkl) | 2.6584 | 0.0490 |
| Q9JHL4 | drebrin-like (Dbnl) | 1.9196 | 0.0306 |
| Q6AXS5 | Serpine1 mRNA binding protein 1 (Serbp1) | 1.9004 | 0.0325 |
| A0JPJ7 | Obg-like ATPase 1 (Ola1) | 0.6494 | 0.0063 |
| Q9JK11 | reticulon 4(Rtn4) | 0.6290 | 0.0282 |
| P41562 | isocitrate dehydrogenase (NADP(+)) 1, cytosolic (Idh1) | 0.5536 | 0.0186 |
| Q99N27 | sorting nexin 1 (Snx1) | 0.5484 | 0.0382 |
| P61314 | ribosomal protein L15 (Rpl15) | 0.5042 | 0.0020 |
| Q9QYL8 | lysophospholipase II (Lypla2) | 0.4824 | 0.0402 |
| P04642 | lactate dehydrogenase A (Ldha) | 0.4518 | 0.0060 |
| O88767 | Parkinsonism associated deglycase (Park7) | 0.4374 | 0.0002 |
| P85972 | vinculin (Vcl) | 0.4353 | 0.0145 |
| Q5PPJ9 | SH3 domain-containing GRB2-like endophilin B2 (Sh3glb2) | 0.4270 | 0.0066 |
| P85845 | fascin actin-bundling protein 1 (Fscn1) | 0.4080 | 0.0048 |
| Q5XFX0 | transgelin 2 (Tagln2) | 0.3826 | 0.0005 |
| P63102 | tyrosine3/5-monooxygenase activation protein, zeta (Ywhaz) | 0.3470 | 0.00008 |
| Q6JE36 | N-myc downstream regulated 1 (Ndrg1) | 0.3191 | 0.0094 |
| Q6GMN2 | Brain-specific angiogenesis inhibitor 1-associated protein 2 (Baiap2) | 0.3057 | 0.0048 |
| Q9Z269 | VAMP associated protein B and C(Vapb) | 0.2950 | 0.0112 |
| Q5EGY4 | YKT6 v-SNARE homolog (S. cerevisiae) (Ykt6) | 0.2830 | 0.00008 |
| P35281 | RAB10, member RAS oncogene family (Rab10) | 0.2583 | 0.0008 |
| P35213 | tyrosine 3/5-monooxygenase activation protein, beta (Ywhab) | 0.1981 | 0.0057 |
| Q9WTT7 | basic leucine zipper and W2 domains 2(Bzw2) | 0.1896 | 0.01543 |
| Q4FZT2 | protein phosphatase methylesterase 1 (Ppme1) | 0.1874 | 0.0014 |
| Q91Y81 | septin 2 (Sept2) | 0.1820 | 0.0022 |
| O35509 | RAB11B, member RAS oncogene family (Rab11b) | 0.1397 | 0.0050 |
| P62260 | tyrosine3/5-monooxygenase activation protein, epsilon (Ywhae) | 0.1304 | 0.0001 |
| Q5I0D1 | glyoxalase domain containing 4 (Glod4) | 0.1077 | 0.0016 |
| Q6NYB7 | RAB1A, member RAS oncogene family (Rab1a) | 0.0591 | 0.0002 |
| Q568Z6 | IST1, ESCRT-III associated factor (Ist1) | 0.0560 | 0.0010 |
| B1H267 | sorting nexin 5 (Snx5) | 0.0494 | 0.0038 |
| Q66HA5 | coiled-coil and C2 domain containing 1A (Cc2d1a) | 0.0348 | 0.0132 |
| Q07205 | eukaryotic translation initiation factor 5 (Eif5) | 0.0286 | 0.0138 |
| UniProt IDs | Protein Names | Fold Change | p-Value |
|---|---|---|---|
| Involved inregulation of actin cytoskeleton | |||
| O88377 | phosphatidylinositol-5-phosphate 4-kinase type 2 beta (Pip4k2b) | 4.4439 | 0.0170 |
| P20171 | GTPase HRas | 0.6631 | 0.0122 |
| P62138 | protein phosphatase 1 catalytic subunit alpha (Ppp1ca) | 0.5881 | 0.0253 |
| P35465 | p21 (RAC1) activated kinase 1 (Pak1) | 0.5060 | 0.0462 |
| P18666 | myosin light chain 12B (Myl12b) | 0.4906 | 0.0014 |
| P45592 | cofilin 1 (Cfl1) | 0.4314 | 0.0001 |
| Q9R0I8 | phosphatidylinositol-5-phosphate 4-kinase type 2 alpha (Pip4k2a) | 0.3561 | 0.0248 |
| P21708 | Mitogen-activated protein kinase 3 (Mapk3) | 0.3446 | 0.0127 |
| Q6GMN2 | Brain-specific angiogenesis inhibitor 1-associated protein 2 (Baiap2) | 0.3057 | 0.0048 |
| P04937 | fibronectin 1 (Fn1) | 0.1998 | 0.0023 |
| Q01986 | mitogen activated protein kinase kinase 1 (Map2k1) | 0.1986 | 0.0032 |
| P63088 | protein phosphatase 1 catalytic subunit gamma (Ppp1cc) | 0.0429 | 0.0010 |
| P85970 | actin related protein 2/3 complex, subunit 2 (Arpc2) | 0.0130 | 0.0004 |
| Involved in both regulation of actin cytoskeleton and bacterial invasion of epithelial cell | |||
| Q5U2U2 | v-crk avian sarcoma virus CT10 oncogene homolog-like (Crkl) | 2.6583 | 0.0489 |
| Q6RUV5 | ras-related C3 botulinum toxin substrate 1(Rac1) | 0.6302 | 0.0478 |
| P85972 | vinculin (Vcl) | 0.4353 | 0.0145 |
| O35346 | protein tyrosine kinase 2 (Ptk2) | 0.4182 | 0.0176 |
| A1L108 | actin related protein 2/3 complex, subunit 5-like (Arpc5l) | 0.4027 | 0.0045 |
| Q99PD4 | actin-related protein 2/3 complex subunit 1A (Arpc1a) | 0.0981 | 0.0001 |
| Involved in bacterial invasion of epithelial cell | |||
| Q66HL2 | Cortactin (Cttn) | 2.2129 | 0.0003 |
| B0BNF1 | septin 8 (Sept8) | 1.7935 | 0.0485 |
| Q08877 | dynamin 3 (Dnm3) | 0.5656 | 0.0287 |
| B3GNI6 | septin 11 (Sept11) | 0.5343 | 0.0069 |
| P08082 | clathrin, light chain B (Cltb) | 0.3047 | 0.0009 |
| Q9WU34 | septin 3 (Sept3) | 0.2932 | 0.0002 |
| P04937 | fibronectin 1 (Fn1) | 0.1998 | 0.0023 |
| Q91Y81 | septin 2 (Sept2) | 0.1820 | 0.0022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, R.; Liu, H.; Lv, F.; Deng, Y.; Li, Y. Rac1/Wave2/Arp3 Pathway Mediates Rat Blood-Brain Barrier Dysfunction under Simulated Microgravity Based on Proteomics Strategy. Int. J. Mol. Sci. 2021, 22, 5165. https://doi.org/10.3390/ijms22105165
Yan R, Liu H, Lv F, Deng Y, Li Y. Rac1/Wave2/Arp3 Pathway Mediates Rat Blood-Brain Barrier Dysfunction under Simulated Microgravity Based on Proteomics Strategy. International Journal of Molecular Sciences. 2021; 22(10):5165. https://doi.org/10.3390/ijms22105165
Chicago/Turabian StyleYan, Ranran, Huayan Liu, Fang Lv, Yulin Deng, and Yujuan Li. 2021. "Rac1/Wave2/Arp3 Pathway Mediates Rat Blood-Brain Barrier Dysfunction under Simulated Microgravity Based on Proteomics Strategy" International Journal of Molecular Sciences 22, no. 10: 5165. https://doi.org/10.3390/ijms22105165