
Molecular Iodine/Cyclophosphamide Synergism on Chemoresistant Neuroblastoma Models

 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. Results In Vitro
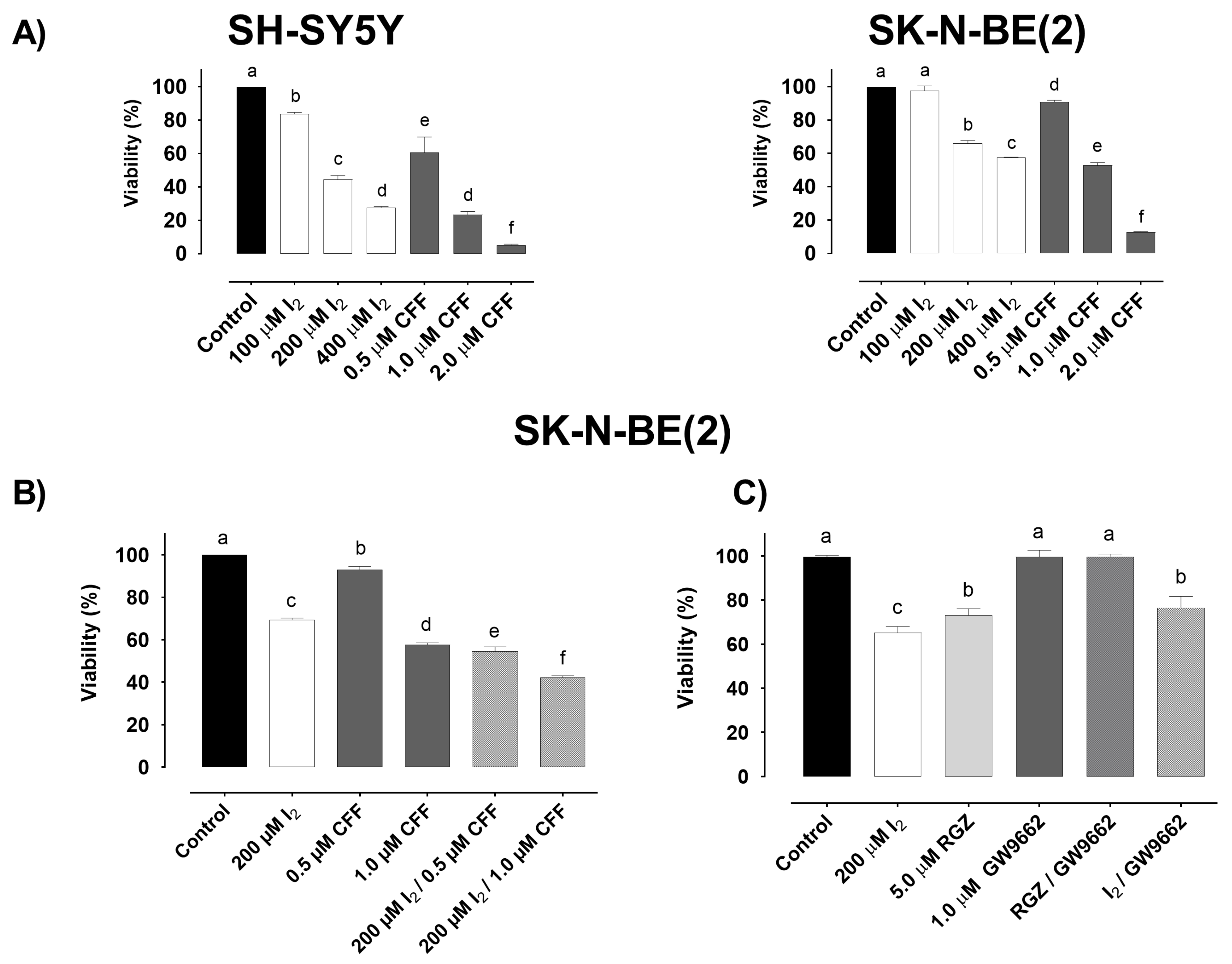
2.1.1. Viability
2.1.2. Apoptosis
2.1.3. Mitochondrial Activity
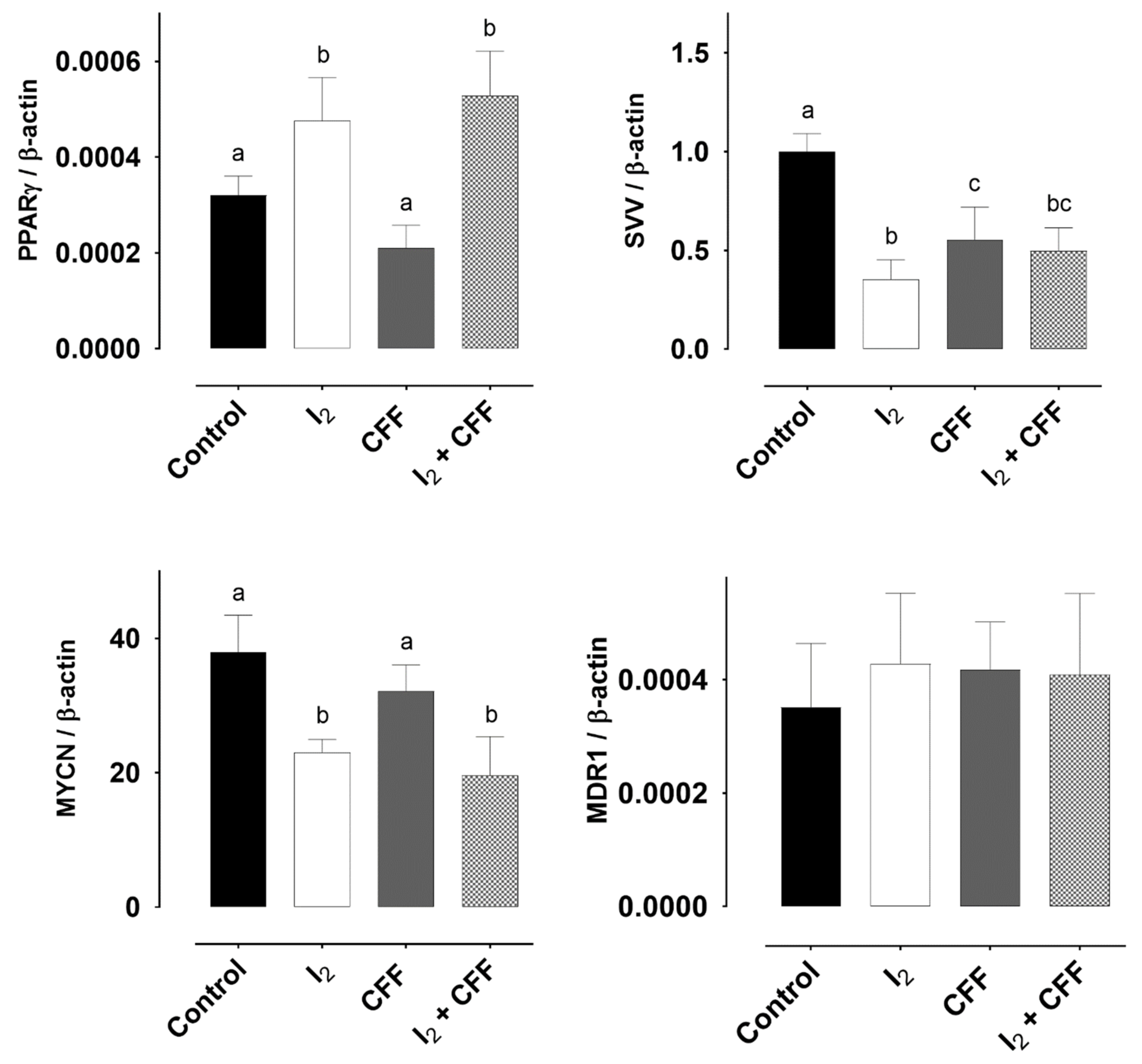
2.1.4. Molecular Response
2.2. Results In Vivo
2.2.1. Tumor Growth
2.2.2. Histopathology
2.2.3. Molecular Response
2.2.4. Preventive Effect of I2 in Bladder Damage
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Viability
4.4. Apoptosis
4.5. Gene Expression
4.6. Mitochondrial Membrane Potential
4.7. Tumoral Implantation and Progression
4.8. Immunohistochemistry
4.9. Western Blot
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Otte, J.; Dyberg, C.; Pepich, A.; Johnsen, J.I. MYCN Function in Neuroblastoma Development. Front. Oncol. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Montemurro, L.; Raieli, S.; Angelucci, S.; Bartolucci, D.; Amadesi, C.; Lampis, S.; Scardovi, A.L.; Venturelli, L.; Nieddu, G.; Cerisoli, L.; et al. A novel MYCN-specific antigene oligonucleotide deregulates mitochondria and inhibits tumor growth in MYCN-amplified neuroblastoma. Cancer Res. 2019, 79, 6166–6177. [Google Scholar] [CrossRef] [Green Version]
- Emadi, A.; Jones, R.J.; Brodsky, R.A. Cyclophosphamide and cancer: Golden anniversary. Nat. Rev. Clin. Oncol. 2009, 6, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Morscher, R.J.; Aminzadeh-Gohari, S.; Hauser-Kronberger, C.; Feichtinger, R.G.; Sperl, W.; Kofler, B. Combination of metronomic cyclophosphamide and dietary intervention inhibits neuroblastoma growth in a CD1-nu mouse model. Oncotarget 2016, 7, 17060–17073. [Google Scholar] [CrossRef] [Green Version]
- Fathpour, G.; Jafari, E.; Hashemi, A.; Dadgar, H.; Shahriari, M.; Zareifar, S.; Jenabzade, A.R.J.; Vali, R.; Ahmadzadehfar, H.; Assadi, M. Feasibility and Therapeutic Potential of Combined Peptide Receptor Radionuclide Therapy With Intensive Chemotherapy for Pediatric Patients With Relapsed or Refractory Metastatic Neuroblastoma. Clin. Nucl. Med. 2021, 46, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Tuyaerts, S.; van Nuffel, A.M.T.; Naert, E.; van Dam, P.A.; Vuylsteke, P.; de Caluwé, A.; Aspeslagh, S.; Dirix, P.; Lippens, L.; de Jaeghere, E.; et al. PRIMMO study protocol: A phase II study combining PD-1 blockade, radiation and immunomodulation to tackle cervical and uterine cancer. BMC Cancer 2019, 19, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aceves, C.; Mendieta, I.; Anguiano, B.; Delgado-González, E. Molecular iodine has extrathyroidal effects as an antioxidant, differentiator, and immunomodulator. Int. J. Mol. Sci. 2021, 22, 1228. [Google Scholar] [CrossRef]
- Mani, S.; Swargiary, G.; Singh, K.K. Natural agents targeting mitochondria in cancer. Int. J. Mol. Sci. 2020, 2, 6992. [Google Scholar] [CrossRef]
- Mendieta, I.; Rodríguez-Gómez, G.; Rueda-Zarazúa, B.; Rodríguez-Castelán, J.; Álvarez-León, W.; Delgado-González, E.; Anguiano, B.; Vázquez-Martínez, O.; Díaz-Muñoz, M.; Aceves, C. Molecular iodine synergized and sensitized neuroblastoma cells to the antineoplastic effect of ATRA. Endocr. Relat. Cancer 2020, 27, 699–710. [Google Scholar] [CrossRef]
- Coulter, D.W.; McGuire, T.R.; Sharp, J.G.; McIntyre, E.M.; Dong, Y.; Wang, X.; Gray, S.; Alexander, G.R.; Chatuverdi, N.K.; Joshi, S.S.; et al. Treatment of a chemoresistant neuroblastoma cell line with the antimalarial ozonide OZ513. BMC Cancer 2016, 16, 867. [Google Scholar] [CrossRef] [Green Version]
- Chesler, L.; Goldenberg, D.D.; Collins, R.; Grimmer, M.; Kim, G.E.; Tihan, T.; Nguyen, K.; Yakovenko, S.; Matthay, K.K.; Weiss, W.A. Chemotherapy-Induced Apoptosis in a Transgenic Model of Neuroblastoma Proceeds Through p53 Induction. Neoplasia 2008, 10, 1268–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, A.; Tiwari, M.; Sinha, R.A.; Kumar, A.; Balapure, A.K.; Bajpai, V.K.; Sharma, R.; Mitra, K.; Tandon, A.; Godbole, M.M. Molecular iodine induces caspase-independent apoptosis in human breast carcinoma cells involving the mitochondria-mediated pathway. J. Biol. Chem. 2006, 281, 19762–19771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousefnia, S.; Momenzadeh, S.; Forootan, F.S.; Ghaedi, K.; Esfahani, M.H.N. The influence of peroxisome proliferator-activated receptor γ (PPARγ) ligands on cancer cell tumorigenicity. Gene 2018, 649, 14–22. [Google Scholar] [CrossRef]
- Arroyo-Helguera, O.; Rojas, E.; Delgado, G.; Aceves, C. Signaling pathways involved in the antiproliferative effect of molecular iodine in normal and tumoral breast cells: Evidence that 6-iodolactone mediates apoptotic effects. Endocr. Relat. Cancer 2008, 15, 1003–1011. [Google Scholar] [CrossRef]
- Anguiano, B.; Aceves, C. Iodine in Mammary and Prostate Pathologies. Curr. Chem. Biol. 2011, 5, 177–182. [Google Scholar] [CrossRef]
- Nuñez-Anita, R.E.; Arroyo-Helguera, O.; Cajero-Juárez, M.; López-Bojorquez, L.; Aceves, C. A complex between 6-iodolactone and the peroxisome proliferator-activated receptor type gamma may mediate the antineoplastic effect of iodine in mammary cancer. Prostaglandins Other Lipid Mediat. 2009, 89, 34–42. [Google Scholar] [CrossRef]
- Nava-Villalba, M.; Nuñez-Anita, R.E.; Bontempo, A.; Aceves, C. Activation of peroxisome proliferator-activated receptor gamma is crucial for antitumoral effects of 6-iodolactone. Mol. Cancer. 2015, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vella, S.; Conaldi, P.G.; Florio, T.; Pagano, A. PPAR Gamma in Neuroblastoma: The Translational Perspectives of Hypoglycemic Drugs. PPAR Research. 2016, 2016, 3038164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushner, B.H.; Cheung, N.K.V.; Modak, S.; Becher, O.J.; Basu, E.M.; Roberts, S.S.; Kramer, K.; Dunkel, I.J. A phase I/Ib trial targeting the Pi3k/Akt pathway using perifosine: Long-term progression-free survival of patients with resistant neuroblastoma. Int. J. Cancer. 2017, 140, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.; Kageyama, H.; Takada, N.; Kawamato, T.; Takayasu, H.; Isogai, E.; Ohira, M.; Hashizume, K.; Kobayashi, H.; Kaneko, Y.; et al. High expression of Survivin, mapped to 17q25, is significantly associated with poor prognostic factors and promotes cell survival in human neuroblastoma. Oncogene 2000, 19, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Nakagawara, A.; Ohira, M. Comprehensive genomics linking between neural development and cancer: Neuroblastoma as a model. Cancer Lett. 2004, 204, 213–224. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Cross-Talk between NADPH Oxidase and Mitochondria: Role in ROS Signaling and Angiogenesis. Cells 2020, 9, 1849. [Google Scholar] [CrossRef] [PubMed]
- Rösner, H.; Torremante, P.; Möller, W.; Gärtner, R. Antiproliferative/cytotoxic activity of molecular iodine and iodolactones in various human carcinoma cell lines. No interfering with EGF-signaling, but evidence for apoptosis. Exp. Clin. Endocrinol. Diabetes 2010, 118, 410–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Duan, N.; Zhang, C.; Zhang, W. Survivin and tumorigenesis: Molecular mechanisms and therapeutic strategies. J. Cancer 2016, 7, 314–323. [Google Scholar] [CrossRef]
- Ausserlechner, M.J.; Hagenbuchner, J. Mitochondrial survivin–an Achilles’ heel in cancer chemoresistance. Mol. Cell. Oncol. 2016, 3, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Brodeur, G.M.; Minturn, J.E.; Ho, R.; Simpson, A.M.; Iyer, R.; Varela, C.R.; Light, J.E.; Kolla, V.; Evans, A.E. Trk receptor expression and inhibition in neuroblastomas. Clin. Cancer Res. 2009, 15, 3244–3250. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Y.; Tong, Y.; Tong, J.; Thiele, C.J. Trk inhibitor attenuates the BDNF/TrkB-induced protection of neuroblastoma cells from etoposide in vitro and in vivo. Cancer Biol. Ther. 2015, 16, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Pin, F.; Barreto, R.; Kitase, Y.; Mitra, S.; Erne, C.E.; Novinger, L.J.; Zimmers, T.A.; Couch, M.E.; Bonewald, L.F.; Bonetto, A. Growth of ovarian cancer xenografts causes loss of muscle and bone mass: A new model for the study of cancer cachexia. J. Cachexia Sarcopenia Muscle 2018, 9, 685–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambaut-Guérin, A.-M.; Martinez, M.-C.; Hamimi, C.; Gauthereau, X.; Nunez, J. Tumor Necrosis Factor Receptors in Neuroblastoma SKNBE Cells and Their Regulation by Retinoic Acid. J. Neurochem. 1995, 65, 537–544. [Google Scholar] [CrossRef]
- Winkler, R. Iodine—A Potential Antioxidant and the Role of Iodine/Iodide in Health and Disease. Nat. Sci. 2015, 7, 548–557. [Google Scholar] [CrossRef] [Green Version]
- Alfaro, Y.; Delgado, G.; Cárabez, A.; Anguiano, B.; Aceves, C. Iodine and doxorubicin, a good combination for mammary cancer treatment: Antineoplastic adjuvancy, chemoresistance inhibition, and cardioprotection. Mol. Cancer 2013, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Liu, L.; Li, L.; Liu, J.; Song, E.; Song, Y. Protective effects of lipoic acid and mesna on cyclophosphamide-induced haemorrhagic cystitis in mice. Cell Biochem. Funct. 2014, 32, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Keles, I.; Bozkurt, M.F.; Cemek, M.; Karalar, M.; Hazini, A.; Alpdagtas, S.; Keles, H.; Yildiz, T.; Ceylan, C.; Buyukokuroglu, M.E. Prevention of cyclophosphamide-induced hemorrhagic cystitis by resveratrol: A comparative experimental study with mesna. Int. Urol. Nephrol. 2014, 46, 833. [Google Scholar] [CrossRef] [PubMed]
- Monach, P.A. Incidence and prevention of bladder toxicity from cyclophosphamide in the treatment of rheumatic diseases: A data-driven review. Arthritis Rheum. 2010, 62, 165. [Google Scholar] [CrossRef]
- Chou, T.C. Drug combination studies and their synergy quantification using the chou-talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendieta, I.; Nuñez-Anita, R.E.; Nava-Villalba, M.; Zambrano-Estrada, X.; Delgado-González, E.; Anguiano, B.; Aceves, C. Molecular iodine exerts antineoplastic effects by diminishing proliferation and invasive potential and activating the immune response in mammary cancer xenografts. BMC Cancer 2019, 19, 261. [Google Scholar] [CrossRef] [PubMed]
- Sander, H.; Wallace, S.; Plouse, R.; Tiwari, S.; Gomes, A.V. Ponceau S waste: Ponceau S staining for total protein normalization. Anal. Biochem. 2019, 575, 44–53. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gen | Reference | Sense | Antisense | bp | Ta (°C) |
|---|---|---|---|---|---|
| TrkA | NM_002529.3 | CATCGTGAAGAGTGGTCTCCG | GAGAGAGACTCCAGAGCGTTGAA | 102 | 60 |
| TrkB | NM_001007097.3 | TCGTGGCATTTCCGAGATTGG | TCGTCAGTTTGTTTCGGGTAAA | 231 | 60 |
| PPARγ | NM_001354666.3 | TCTCTCCGTAATGGAAGACC | GCATTATGAGACATCCCCAC | 474 | 62 |
| SVV | NM_001168.3 | TTCTCAAGGACCACCGCATC | CCAAGTCTGGCTCGTTCTCA | 126 | 60 |
| MDR1 | NM_001348945.2 | GAGAGATCCTCACCAAGCGG | ATCATTGGCGAGCCTGGTAC | 122 | 60 |
| MYCN | NM_001293228.2 | ACCCTGAGCGATTCAGATGAT | GTGGTGACAGCCTTGGTGTT | 113 | 62 |
| P53 | NM_001126118.2 | CCATGAGCGCTGCTCAGATA | GGGCACCACCACACTATGTC | 124 | 60 |
| Bax | NM_138764.5 | AAGCTGAGCGAGTGTCTCAAGCGC | TCCCGCCACAAAGATGGTCACG | 327 | 60 |
| Bcl-2 | NM_000633.3 | CTCGTCGCTACCGTCGTGACTTCG | CAGATGCCGGTTCAGGTACTCAGTC | 242 | 60 |
| VEGF | NM_001025366.3 | CTCGATTGGATGGCAGTAGCT | AGGAGGAGGGCAGAATCATCA | 76 | 60 |
| HIF1 | NM_001530.4 | TTGATGGGATATGAGCCAGA | TGTCCTGTGGTGACTTGTCC | 128 | 60 |
| β-actin | NM_001101.5 | CCATCATGAAGTGTGACGTTG | ACAGAGTACTTGCGCTCAGGA | 175 | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-León, W.; Mendieta, I.; Delgado-González, E.; Anguiano, B.; Aceves, C. Molecular Iodine/Cyclophosphamide Synergism on Chemoresistant Neuroblastoma Models. Int. J. Mol. Sci. 2021, 22, 8936. https://doi.org/10.3390/ijms22168936
Álvarez-León W, Mendieta I, Delgado-González E, Anguiano B, Aceves C. Molecular Iodine/Cyclophosphamide Synergism on Chemoresistant Neuroblastoma Models. International Journal of Molecular Sciences. 2021; 22(16):8936. https://doi.org/10.3390/ijms22168936
Chicago/Turabian StyleÁlvarez-León, Winniberg, Irasema Mendieta, Evangelina Delgado-González, Brenda Anguiano, and Carmen Aceves. 2021. "Molecular Iodine/Cyclophosphamide Synergism on Chemoresistant Neuroblastoma Models" International Journal of Molecular Sciences 22, no. 16: 8936. https://doi.org/10.3390/ijms22168936