
PI3K/PTEN/AKT Signaling Pathways in Germ Cell Development and Their Involvement in Germ Cell Tumors and Ovarian Dysfunctions



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
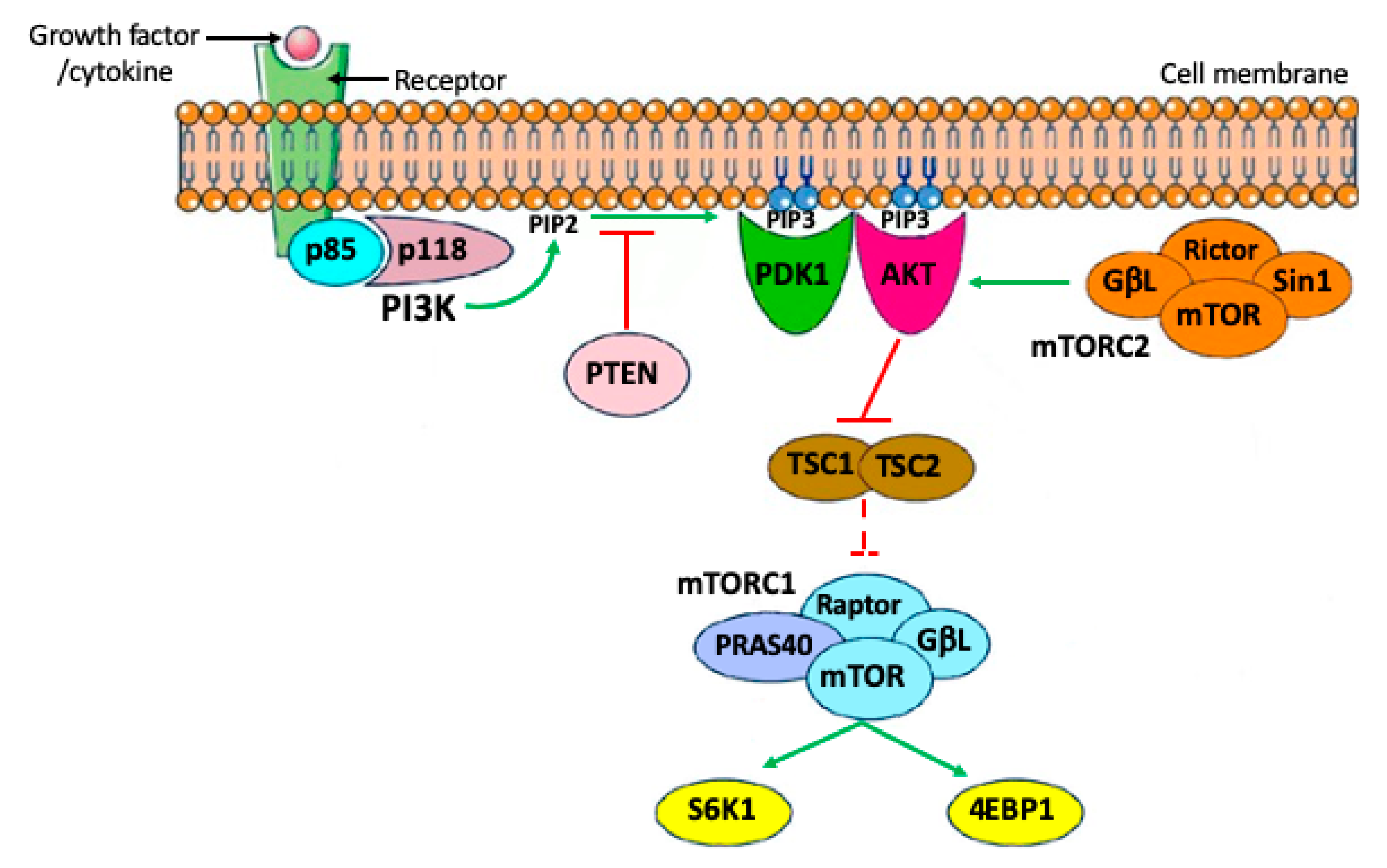
2. Brief Outline of PI3K/PTEN/AKT Signaling Pathways
3. The PI3K/PTEN/AKT Signaling Pathways in PGC Development
3.1. KL/KIT in PGCs as a Major Effector of PI3K/AKT Signaling in PGCs
3.2. The PI3K/PTEN/AKT Signaling Pathways in the Generation of Germ Cell Tumors from PGCs
4. The PI3K/PTEN/AKT Signaling Pathways in Oogenesis and Folliculogenesis
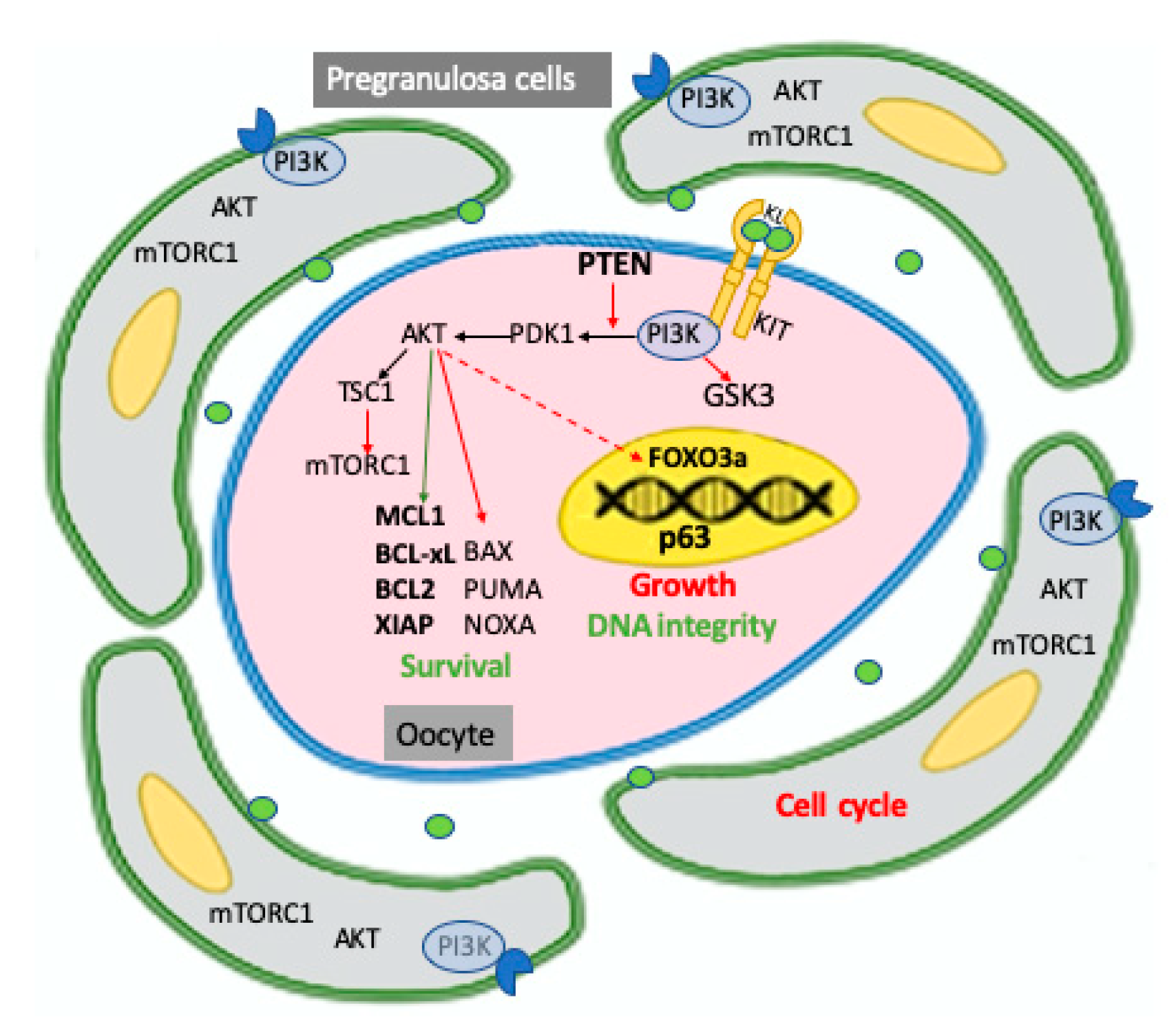
4.1. The PI3K/PTEN/AKT Signaling Pathways in Oocyte Survival/Apoptosis
4.2. The PI3K/PTEN/AKT Signaling Pathways in Primordial Follicle Activation and Development
4.3. The PI3K/PTEN/AKT Signaling Pathways in the Oocyte DNA Damage Response
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Makker, A.; Goel, M.M.; Mahdi, A.A. PI3K/PTEN/Akt and TSC/MTOR Signaling Pathways, Ovarian Dysfunction, and Infertility: An Update. J. Mol. Endocrinol. 2014, 53, R103–R118. [Google Scholar] [CrossRef] [Green Version]
- Maidarti, M.; Anderson, R.A.; Telfer, E.E. Crosstalk between PTEN/PI3K/Akt Signalling and DNA Damage in the Oocyte: Implications for Primordial Follicle Activation, Oocyte Quality and Ageing. Cells 2020, 9, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blume-Jensen, P.; Hunter, T. Oncogenic Kinase Signalling. Nature 2001, 411, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, S.; Hayashi, K.; Kurimoto, K.; Ohta, H.; Yabuta, Y.; Iwanari, H.; Mochizuki, Y.; Hamakubo, T.; Kato, Y.; Shirahige, K.; et al. A Mesodermal Factor, T, Specifies Mouse Germ Cell Fate by Directly Activating Germline Determinants. Dev. Cell 2013, 27, 516–529. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Liu, W.; Lukianchikov, A.; Hancock, G.V.; Zimmerman, J.; Lowe, M.G.; Kim, R.; Galic, Z.; Irie, N.; Surani, M.A.; et al. Germline Competency of Human Embryonic Stem Cells Depends on Eomesodermin. Biol. Reprod. 2017, 97, 850–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakaki, F.; Hayashi, K.; Ohta, H.; Kurimoto, K.; Yabuta, Y.; Saitou, M. Induction of Mouse Germ-Cell Fate by Transcription Factors in Vitro. Nature 2013, 501, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Liu, W.; Zimmerman, J.; Pastor, W.A.; Kim, R.; Hosohama, L.; Ho, J.; Aslanyan, M.; Gell, J.J.; Jacobsen, S.E.; et al. The TFAP2C-Regulated OCT4 Naive Enhancer Is Involved in Human Germline Formation. Cell Rep. 2018, 25, 3591–3602.e5. [Google Scholar] [CrossRef] [Green Version]
- Gkountela, S.; Li, Z.; Vincent, J.J.; Zhang, K.X.; Chen, A.; Pellegrini, M.; Clark, A.T. The Ontogeny of CKIT+ Human Primordial Germ Cells Proves to Be a Resource for Human Germ Line Reprogramming, Imprint Erasure and in Vitro Differentiation. Nat. Cell Biol. 2013, 15, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.W.C.; Dietmann, S.; Irie, N.; Leitch, H.G.; Floros, V.I.; Bradshaw, C.R.; Hackett, J.A.; Chinnery, P.F.; Surani, M.A. A Unique Gene Regulatory Network Resets the Human Germline Epigenome for Development. Cell 2015, 161, 1453–1467. [Google Scholar] [CrossRef] [Green Version]
- Vértesy, Á.; Arindrarto, W.; Roost, M.S.; Reinius, B.; Torrens-Juaneda, V.; Bialecka, M.; Moustakas, I.; Ariyurek, Y.; Kuijk, E.; Mei, H.; et al. Parental Haplotype-Specific Single-Cell Transcriptomics Reveal Incomplete Epigenetic Reprogramming in Human Female Germ Cells. Nat. Commun. 2018, 9, 1873. [Google Scholar] [CrossRef]
- Solter, D.; Knowles, B.B. Monoclonal Antibody Defining a Stage-Specific Mouse Embryonic Antigen (SSEA-1). Proc. Natl. Acad. Sci. USA 1978, 75, 5565–5569. [Google Scholar] [CrossRef] [Green Version]
- Yabuta, Y.; Kurimoto, K.; Ohinata, Y.; Seki, Y.; Saitou, M. Gene Expression Dynamics During Germline Specification in Mice Identified by Quantitative Single-Cell Gene Expression Profiling1. Biol. Reprod. 2006, 75, 705–716. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Molyneaux, K.; Runyan, C.; Schaible, K.; Wylie, C. The Roles of FGF Signaling in Germ Cell Migration in the Mouse. Development 2005, 132, 5399–5409. [Google Scholar] [CrossRef] [Green Version]
- Todaro, F.; Campolo, F.; Barrios, F.; Pellegrini, M.; Di Cesare, S.; Tessarollo, L.; Rossi, P.; Jannini, E.A.; Dolci, S. Regulation of Kit Expression in Early Mouse Embryos and ES Cells: Kit in Embryo and ESC Development. Stem Cells 2019, 37, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Runyan, C.; Shoemaker, A.; Surani, A.; Wylie, C. Steel Factor Controls Primordial Germ Cell Survival and Motility from the Time of Their Specification in the Allantois, and Provides a Continuous Niche throughout Their Migration. Development 2009, 136, 1295–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolci, S.; Williams, D.E.; Ernst, M.K.; Resnick, J.L.; Brannan, C.I.; Lock, L.F.; Lyman, S.D.; Boswell, H.S.; Donovan, P.J. Requirement for Mast Cell Growth Factor for Primordial Germ Cell Survival in Culture. Nature 1991, 352, 809–811. [Google Scholar] [CrossRef] [PubMed]
- Farini, D.; Scaldaferri, M.L.; Iona, S.; La Sala, G.; De Felici, M. Growth Factors Sustain Primordial Germ Cell Survival, Proliferation and Entering into Meiosis in the Absence of Somatic Cells. Dev. Biol. 2005, 285, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Durcova-Hills, G.; Tang, F.; Doody, G.; Tooze, R.; Surani, M.A. Reprogramming Primordial Germ Cells into Pluripotent Stem Cells. PLoS ONE 2008, 3, e3531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sette, C.; Dolci, S.; Geremia, R.; Rossi, P. The Role of Stem Cell Factor and of Alternative C-Kit Gene Products in the Establishment, Maintenance and Function of Germ Cells. Int. J. Dev. Biol. 2000, 44, 599–608. [Google Scholar] [PubMed]
- Tajima, Y.; Moore, M.A.; Soares, V.; Ono, M.; Kissel, H.; Besmer, P. Consequences of Exclusive Expression in Vivo of Kit-Ligand Lacking the Major Proteolytic Cleavage Site. Proc. Natl. Acad. Sci. USA 1998, 95, 11903–11908. [Google Scholar] [CrossRef] [Green Version]
- Godin, I.; Deed, R.; Cooke, J.; Zsebo, K.; Dexter, M.; Wylie, C.C. Effects of the Steel Gene Product on Mouse Primordial Germ Cells in Culture. Nature 1991, 352, 807–809. [Google Scholar] [CrossRef]
- Farini, D.; La Sala, G.; Tedesco, M.; De Felici, M. Chemoattractant Action and Molecular Signaling Pathways of Kit Ligand on Mouse Primordial Germ Cells. Dev. Biol. 2007, 306, 572–583. [Google Scholar] [CrossRef]
- Miguel, M.P.D.; Cheng, L.; Holland, E.C.; Federspiel, M.J.; Donovan, P.J. Dissection of the C-Kit Signaling Pathway in Mouse Primordial Germ Cells by Retroviral-Mediated Gene Transfer. Proc. Natl. Acad. Sci. USA 2002, 99, 10458–10463. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-M.; Yokoyama, T.; Ng, D.; Ulu, F.; Yamazaki, Y. Retinoic Acid-Stimulated ERK1/2 Pathway Regulates Meiotic Initiation in Cultured Fetal Germ Cells. PLoS ONE 2019, 14, e0224628. [Google Scholar] [CrossRef]
- Moe-Behrens, G.H.G.; Klinger, F.G.; Eskild, W.; Grotmol, T.; Haugen, T.B.; De Felici, M. Akt/PTEN Signaling Mediates Estrogen-Dependent Proliferation of Primordial Germ Cells in Vitro. Mol. Endocrinol. 2003, 17, 2630–2638. [Google Scholar] [CrossRef] [Green Version]
- Di Zazzo, E.; Feola, A.; Zuchegna, C.; Romano, A.; Donini, C.F.; Bartollino, S.; Costagliola, C.; Frunzio, R.; Laccetti, P.; Di Domenico, M.; et al. The P85 Regulatory Subunit of PI3K Mediates CAMP-PKA and Insulin Biological Effects on MCF-7 Cell Growth and Motility. Sci. World J. 2014, 2014, 565839. [Google Scholar] [CrossRef] [Green Version]
- Ueda, S.; Mizuki, M.; Ikeda, H.; Tsujimura, T.; Matsumura, I.; Nakano, K.; Daino, H.; Honda, Z.; Sonoyama, J.; Shibayama, H.; et al. Critical Roles of C-Kit Tyrosine Residues 567 and 719 in Stem Cell Factor–Induced Chemotaxis: Contribution of Src Family Kinase and PI3-Kinase on Calcium Mobilization and Cell Migration. Blood 2002, 99, 3342–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felici, M.; Klinger, F.G.; Campolo, F.; Balistreri, C.R.; Barchi, M.; Dolci, S. To Be or Not to Be a Germ Cell: The Extragonadal Germ Cell Tumor Paradigm. Int. J. Mol. Sci. 2021, 22, 5982. [Google Scholar] [CrossRef] [PubMed]
- Dolci, S.; Campolo, F.; De Felici, M. Gonadal Development and Germ Cell Tumors in Mouse and Humans. Semin. Cell Dev. Biol. 2015, 45, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Marcozzi, S.; Rossi, V.; Salustri, A.; De Felici, M.; Klinger, F.G. Programmed Cell Death in the Human Ovary. Minerva Ginecol. 2018, 70, 549–560. [Google Scholar] [CrossRef]
- Clarkson, Y.L.; McLaughlin, M.; Waterfall, M.; Dunlop, C.E.; Skehel, P.A.; Anderson, R.A.; Telfer, E.E. Initial Characterisation of Adult Human Ovarian Cell Populations Isolated by DDX4 Expression and Aldehyde Dehydrogenase Activity. Sci. Rep. 2018, 8, 6953. [Google Scholar] [CrossRef] [Green Version]
- White, Y.A.R.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte Formation by Mitotically Active Germ Cells Purified from Ovaries of Reproductive-Age Women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujihara, M.; Comizzoli, P.; Keefer, C.L.; Wildt, D.E.; Songsasen, N. Epidermal Growth Factor (EGF) Sustains In Vitro Primordial Follicle Viability by Enhancing Stromal Cell Proliferation via MAPK and PI3K Pathways in the Prepubertal, but Not Adult, Cat Ovary1. Biol. Reprod. 2014, 90, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, M.É.S.; Barberino, R.S.; Menezes, V.G.; Gouveia, B.B.; Macedo, T.J.S.; Santos, J.M.S.; Monte, A.P.O.; Barros, V.R.P.; Matos, M.H.T. Insulin-like Growth Factor-1 (IGF-1) Promotes Primordial Follicle Growth and Reduces DNA Fragmentation through the Phosphatidylinositol 3-Kinase/Protein Kinase B (PI3K/AKT) Signalling Pathway. Reprod. Fertil. Dev. 2018, 30, 1503–1513. [Google Scholar] [CrossRef]
- Liu, K.; Rajareddy, S.; Liu, L.; Jagarlamudi, K.; Boman, K.; Selstam, G.; Reddy, P. Control of Mammalian Oocyte Growth and Early Follicular Development by the Oocyte PI3 Kinase Pathway: New Roles for an Old Timer. Dev. Biol. 2006, 299, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hutt, K.J.; McLaughlin, E.A.; Holland, M.K. Kit Ligand and C-Kit Have Diverse Roles during Mammalian Oogenesis and Folliculogenesis. Mol. Hum. Reprod. 2006, 12, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Orr-Urtreger, A.; Avivi, A.; Zimmer, Y.; Givol, D.; Yarden, Y.; Lonai, P. Developmental Expression of C-Kit, a Proto-Oncogene Encoded by the W Locus. Development 1990, 109, 911–923. [Google Scholar] [CrossRef]
- Manova, K.; Huang, E.J.; Angeles, M.; De Leon, V.; Sanchez, S.; Pronovost, S.M.; Besmer, P.; Bachvarova, R.F. The Expression Pattern of the C-Kit Ligand in Gonads of Mice Supports a Role for the c-Kit Receptor in Oocyte Growth and in Proliferation of Spermatogonia. Dev. Biol. 1993, 157, 85–99. [Google Scholar] [CrossRef]
- Horie, K.; Takakura, K.; Taii, S.; Narimoto, K.; Noda, Y.; Nishikawa, S.; Nakayama, H.; Fujita, J.; Mori, T. The Expression of C-Kit Protein during Oogenesis and Early Embryonic Development1. Biol. Reprod. 1991, 45, 547–552. [Google Scholar] [CrossRef]
- Horie, K.; Fujita, J.; Takakura, K.; Kanzaki, H.; Suginami, H.; Iwai, M.; Nakayama, H.; Mori, T. Pregnancy: The Expression of c-Kit Protein in Human Adult and Fetal Tissues. Hum. Reprod. 1993, 8, 1955–1962. [Google Scholar] [CrossRef]
- Motro, B.; Bernstein, A. Dynamic Changes in Ovarian C-Kit and Steel Expression during the Estrous Reproductive Cycle. Dev. Dyn. 1993, 197, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Laitinen, M.; Rutanen, E.M.; Ritvos, O. Expression of C-Kit Ligand Messenger Ribonucleic Acids in Human Ovaries and Regulation of Their Steady State Levels by Gonadotropins in Cultured Granulosa-Luteal Cells. Endocrinology 1995, 136, 4407–4414. [Google Scholar] [CrossRef] [PubMed]
- Ismail, R.S.; Okawara, Y.; Fryer, J.N.; Vanderhyden, B.C. Hormonal Regulation of the Ligand for C-Kit in the Rat Ovary and Its Effects on Spontaneous Oocyte Meiotic Maturation. Mol. Reprod. Dev. 1996, 43, 458–469. [Google Scholar] [CrossRef]
- Silva, J.R.V.; Figueiredo, J.R.; van den Hurk, R. Involvement of Growth Hormone (GH) and Insulin-like Growth Factor (IGF) System in Ovarian Folliculogenesis. Theriogenology 2009, 71, 1193–1208. [Google Scholar] [CrossRef] [PubMed]
- Poljicanin, A.; Filipovic, N.; Vukusic Pusic, T.; Soljic, V.; Caric, A.; Saraga-Babic, M.; Vukojevic, K. Expression Pattern of RAGE and IGF-1 in the Human Fetal Ovary and Ovarian Serous Carcinoma. Acta Histochem. 2015, 117, 468–476. [Google Scholar] [CrossRef]
- Louhio, H.; Hovatta, O.; Sjöberg, J.; Tuuri, T. The Effects of Insulin, and Insulin-like Growth Factors I and II on Human Ovarian Follicles in Long-Term Culture. Mol. Hum. Reprod. 2000, 6, 694–698. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo Signaling Disruption and Akt Stimulation of Ovarian Follicles for Infertility Treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Feng, Y.; Jansson, L.; Sato, Y.; Deguchi, M.; Kawamura, K.; Hsueh, A.J. Actin Polymerization-Enhancing Drugs Promote Ovarian Follicle Growth Mediated by the Hippo Signaling Effector YAP. FASEB J. 2015, 29, 2423–2430. [Google Scholar] [CrossRef] [Green Version]
- Richani, D.; Gilchrist, R.B. The Epidermal Growth Factor Network: Role in Oocyte Growth, Maturation and Developmental Competence. Hum. Reprod. Update 2018, 24, 1–14. [Google Scholar] [CrossRef] [Green Version]
- De Felici, M.; Klinger, F.G. Programmed Cell Death in Mouse Primordial Germ Cells. Int. J. Dev. Biol. 2015, 59, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Ene, A.C.; Park, S.; Edelmann, W.; Taketo, T. Caspase 9 Is Constitutively Activated in Mouse Oocytes and Plays a Key Role in Oocyte Elimination during Meiotic Prophase Progression. Dev. Biol. 2013, 377, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Klinger, F.G.; Rossi, V.; De Felici, M. Multifaceted Programmed Cell Death in the Mammalian Fetal Ovary. Int. J. Dev. Biol. 2015, 59, 51–54. [Google Scholar] [CrossRef] [Green Version]
- De Felici, M.; Lobascio, A.M.; Klinger, F.G. Cell Death in Fetal Oocytes: Many Players for Multiple Pathways. Autophagy 2008, 4, 240–242. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Klinger, F.G.; Liu, J.; De Felici, M.; Shen, W.; Sun, X. MiR-378-3p Maintains the Size of Mouse Primordial Follicle Pool by Regulating Cell Autophagy and Apoptosis. Cell Death Dis. 2020, 11, 737. [Google Scholar] [CrossRef] [PubMed]
- Markholt, S.; Grøndahl, M.L.; Ernst, E.H.; Andersen, C.Y.; Ernst, E.; Lykke-Hartmann, K. Global Gene Analysis of Oocytes from Early Stages in Human Folliculogenesis Shows High Expression of Novel Genes in Reproduction. MHR Basic Sci. Reprod. Med. 2012, 18, 96–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsueh, A.J.W.; Kawamura, K.; Cheng, Y.; Fauser, B.C.J.M. Intraovarian Control of Early Folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.; Adhikari, D.; Zheng, W.; Liang, S.; Hämäläinen, T.; Tohonen, V.; Ogawa, W.; Noda, T.; Volarevic, S.; Huhtaniemi, I.; et al. PDK1 Signaling in Oocytes Controls Reproductive Aging and Lifespan by Manipulating the Survival of Primordial Follicles. Hum. Mol. Genet. 2009, 18, 2813–2824. [Google Scholar] [CrossRef] [Green Version]
- Jagarlamudi, K.; Liu, L.; Adhikari, D.; Reddy, P.; Idahl, A.; Ottander, U.; Lundin, E.; Liu, K. Oocyte-Specific Deletion of Pten in Mice Reveals a Stage-Specific Function of PTEN/PI3K Signaling in Oocytes in Controlling Follicular Activation. PLoS ONE 2009, 4, e6186. [Google Scholar] [CrossRef]
- Zhang, M.; Su, Y.-Q.; Sugiura, K.; Xia, G.; Eppig, J.J. Granulosa Cell Ligand NPPC and Its Receptor NPR2 Maintain Meiotic Arrest in Mouse Oocytes. Science 2010, 330, 366–369. [Google Scholar] [CrossRef] [Green Version]
- Andrade, G.M.; da Silveira, J.C.; Perrini, C.; Del Collado, M.; Gebremedhn, S.; Tesfaye, D.; Meirelles, F.V.; Perecin, F. The Role of the PI3K-Akt Signaling Pathway in the Developmental Competence of Bovine Oocytes. PLoS ONE 2017, 12, e0185045. [Google Scholar] [CrossRef]
- Choi, J.; Jo, M.; Lee, E.; Choi, D. AKT Is Involved in Granulosa Cell Autophagy Regulation via MTOR Signaling during Rat Follicular Development and Atresia. Reproduction 2014, 147, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, D.; Liu, K. MTOR Signaling in the Control of Activation of Primordial Follicles. Cell Cycle 2010, 9, 1673–1674. [Google Scholar] [CrossRef] [Green Version]
- Gorre, N.; Adhikari, D.; Lindkvist, R.; Brännström, M.; Liu, K.; Shen, Y. MTORC1 Signaling in Oocytes Is Dispensable for the Survival of Primordial Follicles and for Female Fertility. PLoS ONE 2014, 9, e110491. [Google Scholar] [CrossRef]
- Castrillon, D.H.; Miao, L.; Kollipara, R.; Horner, J.W.; DePinho, R.A. Suppression of Ovarian Follicle Activation in Mice by the Transcription Factor Foxo3a. Science 2003, 301, 215–218. [Google Scholar] [CrossRef]
- Rajareddy, S.; Reddy, P.; Du, C.; Liu, L.; Jagarlamudi, K.; Tang, W.; Shen, Y.; Berthet, C.; Peng, S.L.; Kaldis, P.; et al. P27kip1 (Cyclin-Dependent Kinase Inhibitor 1B) Controls Ovarian Development by Suppressing Follicle Endowment and Activation and Promoting Follicle Atresia in Mice. Mol. Endocrinol. 2007, 21, 2189–2202. [Google Scholar] [CrossRef]
- Pelosi, E.; Omari, S.; Michel, M.; Ding, J.; Amano, T.; Forabosco, A.; Schlessinger, D.; Ottolenghi, C. Constitutively Active Foxo3 in Oocytes Preserves Ovarian Reserve in Mice. Nat. Commun. 2013, 4, 1843. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. (Lausanne) 2019, 10, 811. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, Q.-N.; Zerafa, N.; Liew, S.H.; Morgan, F.H.; Strasser, A.; Scott, C.L.; Findlay, J.K.; Hickey, M.; Hutt, K.J. Loss of PUMA Protects the Ovarian Reserve during DNA-Damaging Chemotherapy and Preserves Fertility. Cell Death Dis. 2018, 9, 618. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Luo, L.-L.; Qian, Y.-S.; Fu, Y.-C.; Sui, X.-X.; Geng, Y.-J.; Huang, D.-N.; Gao, S.-T.; Zhang, R.-L. FOXO3a Is Involved in the Apoptosis of Naked Oocytes and Oocytes of Primordial Follicles from Neonatal Rat Ovaries. Biochem. Biophys. Res. Commun. 2009, 381, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Di Emidio, G.; Falone, S.; Vitti, M.; D’Alessandro, A.M.; Vento, M.; Di Pietro, C.; Amicarelli, F.; Tatone, C. SIRT1 Signalling Protects Mouse Oocytes against Oxidative Stress and Is Deregulated during Aging. Hum. Reprod. 2014, 29, 2006–2017. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Rajareddy, S.; Reddy, P.; Jagarlamudi, K.; Du, C.; Shen, Y.; Guo, Y.; Boman, K.; Lundin, E.; Ottander, U.; et al. Phosphorylation and Inactivation of Glycogen Synthase Kinase-3 by Soluble Kit Ligand in Mouse Oocytes during Early Follicular Development. J. Mol. Endocrinol. 2007, 38, 137–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, G.B.; Shidler, M.J.; Besmer, P.; Castrillon, D.H. Kit Signaling via PI3K Promotes Ovarian Follicle Maturation but Is Dispensable for Primordial Follicle Activation. Dev. Biol. 2009, 331, 292–299. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Kawamura, N.; Hsueh, A.J.W. Activation of Dormant Follicles: A New Treatment for Premature Ovarian Failure? Curr. Opin. Obstet. Gynecol. 2016, 28, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, G.B.; Zielonka, E.M.; Coutandin, D.; Weber, T.A.; Schäfer, B.; Hannewald, J.; Luh, L.M.; Durst, F.G.; Ibrahim, M.; Hoffmann, J.; et al. DNA Damage in Oocytes Induces a Switch of the Quality Control Factor TAp63α from Dimer to Tetramer. Cell 2011, 144, 566–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lena, A.M.; Rossi, V.; Osterburg, S.; Smirnov, A.; Osterburg, C.; Tuppi, M.; Cappello, A.; Amelio, I.; Dötsch, V.; De Felici, M.; et al. The P63 C-Terminus Is Essential for Murine Oocyte Integrity. Nat. Commun. 2021, 12, 383. [Google Scholar] [CrossRef]
- Tuppi, M.; Kehrloesser, S.; Coutandin, D.W.; Rossi, V.; Luh, L.M.; Strubel, A.; Hötte, K.; Hoffmeister, M.; Schäfer, B.; De Oliveira, T.; et al. Oocyte DNA Damage Quality Control Requires Consecutive Interplay of CHK2 and CK1 to Activate P63. Nat. Struct. Mol. Biol. 2018, 25, 261–269. [Google Scholar] [CrossRef]
- Kerr, J.B.; Hutt, K.J.; Michalak, E.M.; Cook, M.; Vandenberg, C.J.; Liew, S.H.; Bouillet, P.; Mills, A.; Scott, C.L.; Findlay, J.K.; et al. DNA Damage-Induced Primordial Follicle Oocyte Apoptosis and Loss of Fertility Require TAp63-Mediated Induction of Puma and Noxa. Mol. Cell 2012, 48, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-L.; Sengupta, S.; Gurdziel, K.; Bell, G.W.; Jacks, T.; Flores, E.R. P63 and P73 Transcriptionally Regulate Genes Involved in DNA Repair. PLoS Genet. 2009, 5, e1000680. [Google Scholar] [CrossRef]
- Abraham, A.G.; O’Neill, E. PI3K/Akt-Mediated Regulation of P53 in Cancer. Biochem. Soc. Trans. 2014, 42, 798–803. [Google Scholar] [CrossRef]
- Ye, S.; Lee, K.B.; Park, M.H.; Lee, J.-S.; Kim, S.M. P63 Regulates Growth of Esophageal Squamous Carcinoma Cells via the Akt Signaling Pathway. Int. J. Oncol. 2014, 44, 2153–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Lao, Y.; Zhang, Y.; Gillespie, D.A. Akt: A Double-Edged Sword in Cell Proliferation and Genome Stability. J. Oncol. 2012, 2012, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rossi, V.; Lispi, M.; Longobardi, S.; Mattei, M.; Rella, F.D.; Salustri, A.; De Felici, M.; Klinger, F.G. LH Prevents Cisplatin-Induced Apoptosis in Oocytes and Preserves Female Fertility in Mouse. Cell Death Differ. 2017, 24, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felici, M.; Klinger, F.G. PI3K/PTEN/AKT Signaling Pathways in Germ Cell Development and Their Involvement in Germ Cell Tumors and Ovarian Dysfunctions. Int. J. Mol. Sci. 2021, 22, 9838. https://doi.org/10.3390/ijms22189838
De Felici M, Klinger FG. PI3K/PTEN/AKT Signaling Pathways in Germ Cell Development and Their Involvement in Germ Cell Tumors and Ovarian Dysfunctions. International Journal of Molecular Sciences. 2021; 22(18):9838. https://doi.org/10.3390/ijms22189838
Chicago/Turabian StyleDe Felici, Massimo, and Francesca Gioia Klinger. 2021. "PI3K/PTEN/AKT Signaling Pathways in Germ Cell Development and Their Involvement in Germ Cell Tumors and Ovarian Dysfunctions" International Journal of Molecular Sciences 22, no. 18: 9838. https://doi.org/10.3390/ijms22189838