
Pseudomonas donghuensis HYS gtrA/B/II Gene Cluster Contributes to Its Pathogenicity toward Caenorhabditis elegans

Abstract
:1. Introduction
2. Results
2.1. Screening of Specific Virulence Genes Based on Virulence Factor Prediction and Comparative Genomic Analysis
2.2. The Particularity of GtrA, GtrB and GtrII in Pseudomonas
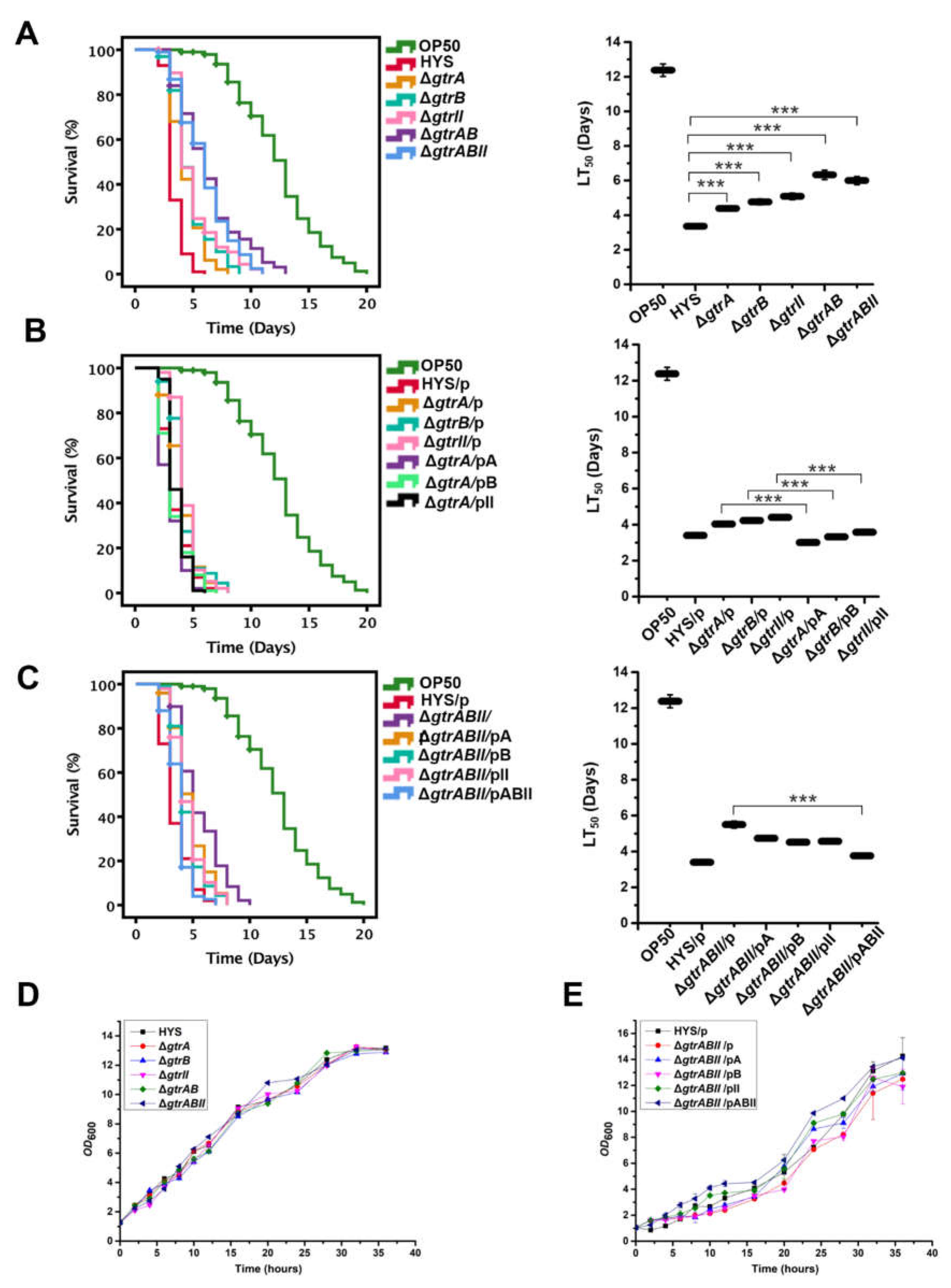
2.3. The Virulence of P. donghuensis HYS Is Optimized by the gtrA/B/II Virulence Gene Cluster in a C. elegans Slow-Killing Assay
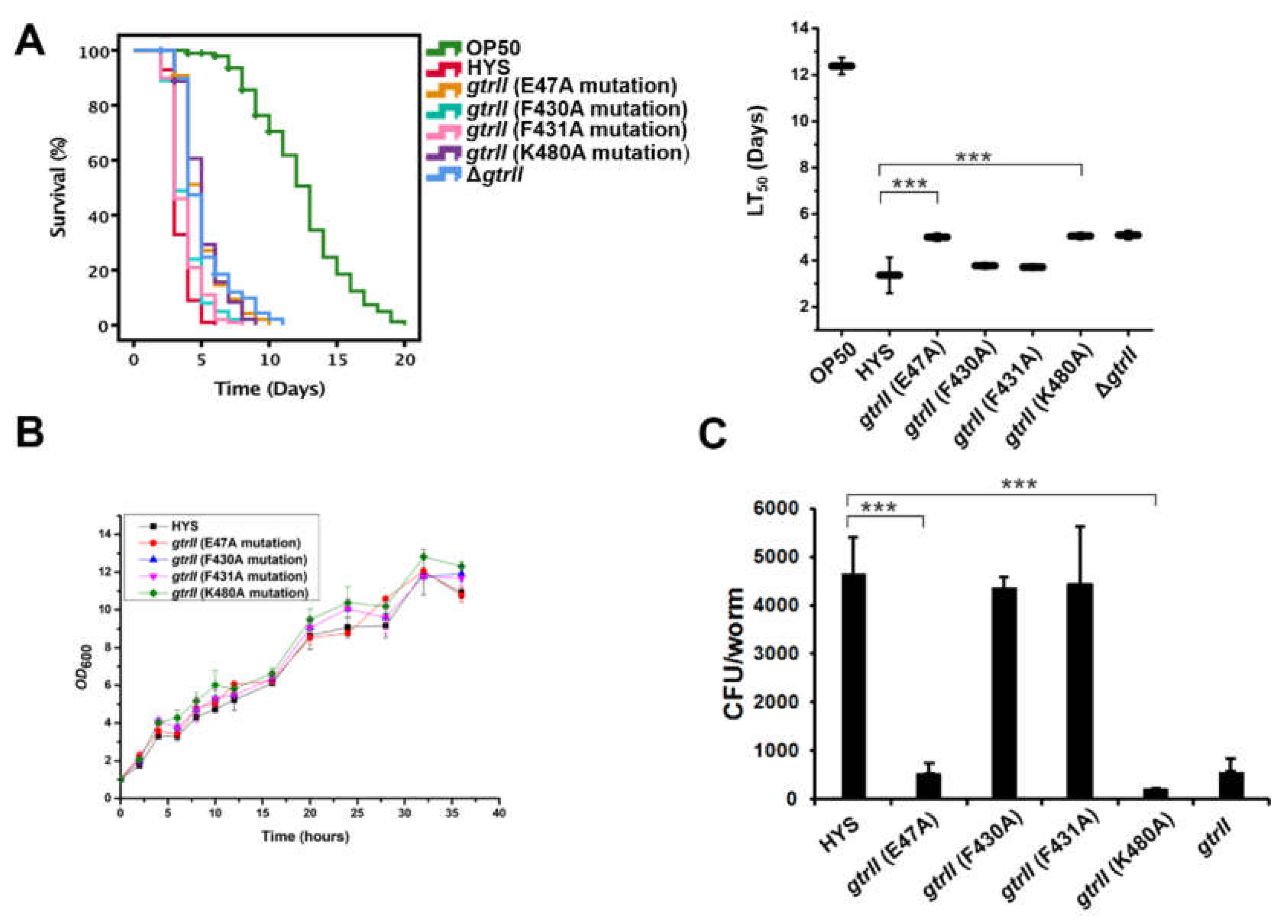
2.4. Identification of Critical GtrII Residues in P. donghuensis HYS
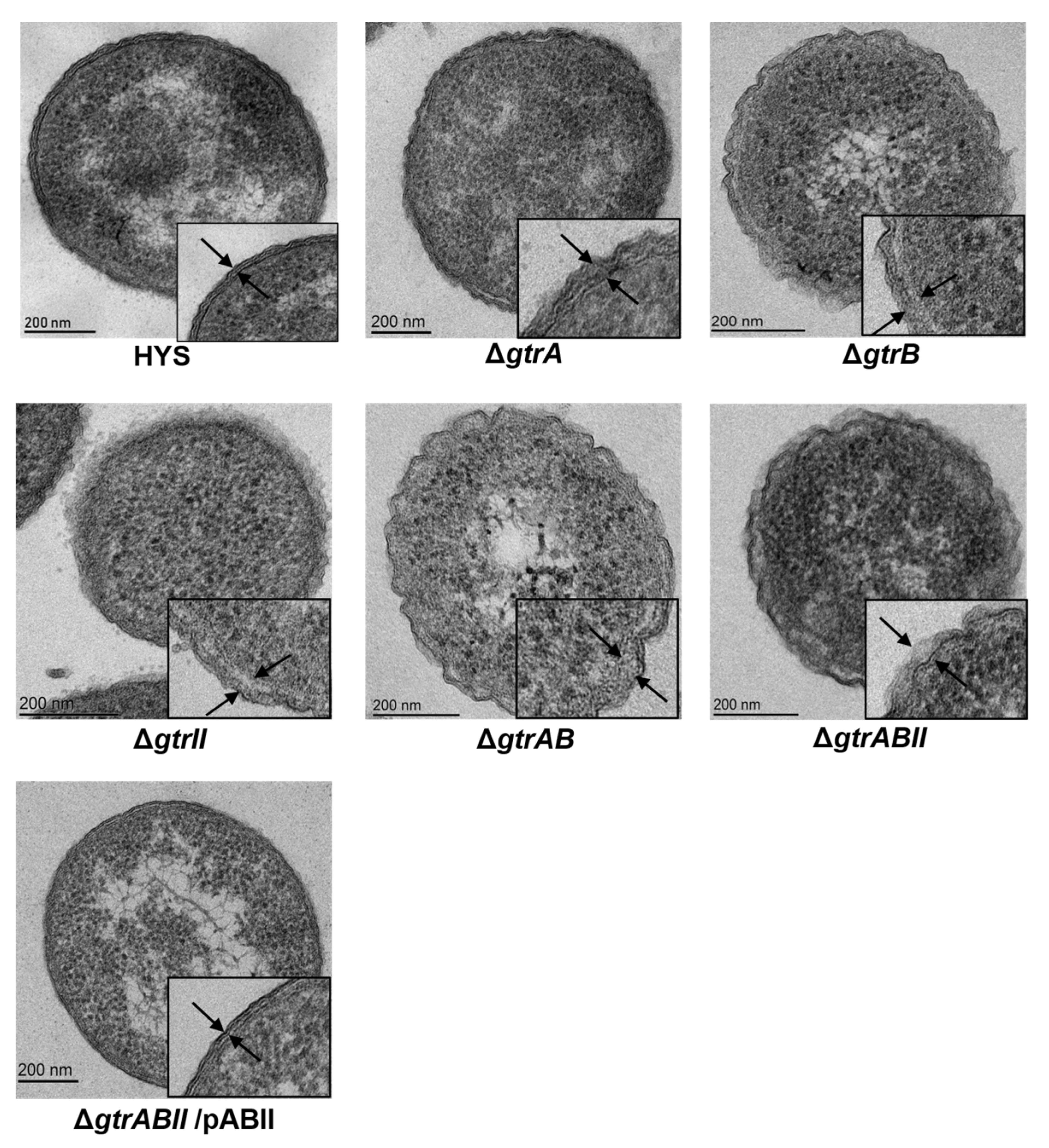
2.5. Surface Topology of Strains Revealed by Transmission Electron Microscopy
2.6. Estimation of P. donghuensis HYS CFU within the C. elegans Gut
2.7. GtrA/B/II-Involved Virulence Is Related to C. elegans Innate Immunity
3. Discussion
4. Materials and Methods
4.1. Bacteria, Nematodes and Cultivation Conditions
4.2. Comparative Genomics Analysis
4.3. Screening of Virulence Factors
4.4. Construction of Mutant and Complement P. donghuensis HYS
4.5. Construction of Point Mutations in P. donghuensis HYS
4.6. Slow-Killing Assays of C. elegans
4.7. Growth Curve Assay
4.8. Transmission Electron Microscopy
4.9. C. elegans Bacterial CFU Analysis
4.10. Microscopy
4.11. Statistical Analysis
4.12. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hornef, M.W.; Wick, M.J.; Rhen, M.; Normark, S. Bacterial strategies for overcoming host innate and adaptive immune responses. Nat. Immunol. 2002, 3, 1033–1040. [Google Scholar] [CrossRef]
- Rohmer, L.; Hocquet, D.; Miller, S.I. Are pathogenic bacteria just looking for food? Metabolism and microbial pathogenesis. Trends Microbiol. 2011, 19, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.R.; Hu, Y.H.; Zhang, W.W.; Sun, L. Construction of an attenuated Pseudomonas fluorescens strain and evaluation of its potential as a cross-protective vaccine. Vaccine 2009, 27, 4047–4055. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.; Porcel, M.; de la Torre, J.; Molina-Henares, M.A.; Daddaoua, A.; Llamas, M.A.; Roca, A.; Carriel, V.; Garzon, I.; Ramos, J.L.; et al. Analysis of the pathogenic potential of nosocomial Pseudomonas putida strains. Front. Microbiol. 2015, 6, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Al-Wrafy, F.; Brzozowska, E.; Gorska, S.; Gamian, A. Pathogenic factors of Pseudomonas aeruginosa—The role of biofilm in pathogenicity and as a target for phage therapy. Postep. Hig. Med. Dosw. 2017, 71, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.F.; He, S.Y. Pseudomonas syringae pv. Tomato DC3000: A model pathogen for probing disease susceptibility and hormone signaling in plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef]
- Hoiby, N. Pseudomonas aeruginosa infection in cystic fibrosis. Relationship between mucoid strains of Pseudomonas aeruginosa and the humoral immune response. Acta Pathol. Microbiol. Scand. 1974, 82, 551–558. [Google Scholar] [CrossRef]
- Reynolds, H.Y.; Levine, A.S.; Wood, R.E.; Zierdt, C.H.; Dale, D.C.; Pennington, J.E.; Maryland, B. Pseudomonas aeruginosa infections: Persisting problems and current research to find new therapies. Ann. Intern. Med. 1975, 82, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Dzvova, N.; Colmer-Hamood, J.A.; Griswold, J.A.; Hamood, A.N. Isolation and characterization of HepP: A virulence-related Pseudomonas aeruginosa heparinase. BMC Microbiol. 2017, 17, 233–248. [Google Scholar] [CrossRef]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The antimicrobial volatile power of the rhizospheric isolate Pseudomonas donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef] [Green Version]
- Agaras, B.C.; Iriarte, A.; Valverde, C.F. Genomic insights into the broad antifungal activity, plant-probiotic properties, and their regulation, in Pseudomonas donghuensis strain SVBP6. PLoS ONE 2018, 13, e0194088. [Google Scholar] [CrossRef]
- Gao, J.; Yu, X.; Xie, Z. Draft genome sequence of high-siderophore-yielding Pseudomonas sp. strain HYS. J. Bacteriol. 2012, 194, 4121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Xie, G.; Peng, F.; Xie, Z. Pseudomonas donghuensis sp. nov., exhibiting high-yields of siderophore. Antonie Van Leeuwenhoek 2015, 107, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Y.; Chen, M.; Jiang, Z.; Hu, Y.; Xie, Z.X. The two-component regulators GacS and GacA positively regulate a nonfluorescent siderophore through the Gac/Rsm signaling cascade in high-siderophore-yielding Pseudomonas sp. Strain HYS. J. Bacteriol. 2014, 196, 3259–3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, G.F.; Zeng, M.; You, J.; Xie, Z.X. Pseudomonas donghuensis HYS virulence towards Caenorhabditis elegans is regulated by the Cbr/Crc system. Sci. Rep. 2019, 9, 8772. [Google Scholar] [CrossRef] [PubMed]
- Gui, Z.; You, J.; Xie, G.; Qin, Y.; Wu, T.; Xie, Z. Pseudomonas donghuensis HYS 7-hydroxytropolone contributes to pathogenicity toward Caenorhabditis elegans and is influenced by pantothenic acid. Biochem. Biophys. Res. Commun. 2020, 533, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Yue, Y.; Park, Y. A living model for obesity and aging research: Caenorhabditis elegans. Crit. Rev. Food Sci. Nutr. 2018, 58, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, L.P.; Luke, C.J.; Perlmutter, D.H.; Silverman, G.A.; Pak, S.C. C. elegans in high-throughput drug discovery. Adv. Drug Deliv. Rev. 2014, 69, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Markaki, M.; Tavernarakis, N. Modeling human disease in Caenorhabditis elegans. Biotechnol. J. 2010, 5, 1261–1276. [Google Scholar] [CrossRef]
- Sorathia, N.; Rajadhyaksha, M.S. Caenorhabditis elegans: A Model for Studying Human Pathogen Biology. Recent Pat. Biotechnol. 2016, 10, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; Veen, J.A.; Elsas, J.D. Assessment of the diversity, and antagonism towards Rhizoctonia solani AG3, of Pseudomonas species in soil from different agricultural regimes. FEMS Microbiol. Ecol. 2004, 47, 51–64. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Sun, Q.; Senchenkova, S.N.; Perepolov, A.V.; Shashkov, A.S.; Xu, J. O-antigen modifications providing antigenic diversity of Shigella flexneri and underlying genetic mechanisms. Biochemistry 2015, 80, 901–914. [Google Scholar] [CrossRef]
- Kolly, G.S.; Mukherjee, R.; Kilacskova, E.; Abriata, L.A.; Raccaud, M.; Blasko, J.; Sala, C.; Dal, P.M.; Mikusova, K.; Cole, S.T. GtrA protein RV3789 is required for arabinosylation of arabinogalactan in Mycobacterium tuberculosis. J. Bacteriol. 2015, 197, 3686–3697. [Google Scholar] [CrossRef] [Green Version]
- Ardiccioni, C.; Clarke, O.B.; Tomasek, D.; Issa, H.A.; von Alpen, D.C.; Pond, H.L.; Banerjee, S.; Rajashankar, K.R.; Liu, Q.; Guan, Z.; et al. Structure of the polyisoprenyl-phosphate glycosyltransferase GtrB and insights into the mechanism of catalysis. Nat. Commun. 2016, 7, 10175–10183. [Google Scholar] [CrossRef] [Green Version]
- Okada, N.; Sasakawa, C.; Tobe, T.; Yamada, M.; Nagai, S.; Talukder, K.A.; Komatsu, K.; Kanegasaki, S.; Yoshikawa, M. Virulence-associated chromosomal loci of Shigella flexneri identified by random Tn5 insertion mutagenesis. Mol. Microbiol. 1991, 5, 187–195. [Google Scholar] [CrossRef]
- Guan, S.; Bastin, D.A.; Verma, N.K. Functional analysis of the O antigen glucosylation gene cluster of Shigella flexneri bacteriophage SfX. Microbiology 1999, 145, 1263–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavris, M.; Manning, P.A.; Morona, R. Mechanism of bacteriophage SfII-mediated serotype conversion in Shigella flexneri. Mol. Microbiol. 1997, 26, 939–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, N.O.; Riccio, C.; Dallaire, A.; Price, J.; Jenkins, B.; Koulman, A.; Miska, E.A. Cysteine synthases CYSL-1 and CYSL-2 mediate C. elegans heritable adaptation to P. vranovensis infection. Nat. Commun. 2020, 11, 1741–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, C.; Huptas, C.; von Neubeck, M.; Scherer, S.; Wenning, M.; Lücking, G. Genetic Organization of the aprX-lipA2 Operon Affects the Proteolytic Potential of Pseudomonas Species in Milk. Front. Microbiol. 2020, 11, 1190–1203. [Google Scholar] [CrossRef]
- Khan, A.H.; Anees, M.; Arshad, M.; Muhammad, Y.S.; Iqbal, M.; Yousaf, S. Effects of illuminance and nutrients on bacterial photo-physiology of hydrocarbon degradation. Sci. Total Environ. 2016, 557, 705–711. [Google Scholar] [CrossRef]
- Tribelli, P.M.; Raiger Iustman, L.J.; Catone, M.V.; Di Martino, C.; Revale, S.; Méndez, B.S.; López, N.I. Genome sequence of the polyhydroxybutyrate producer Pseudomonas extremaustralis, a highly stress-resistant Antarctic bacterium. J. Bacteriol. 2012, 194, 2381–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehane, A.M.; Korres, H.; Verma, N.K. Bacteriophage-encoded glucosyltransferase GtrII of Shigella flexneri: Membrane topology and identification of critical residues. Biochem. J. 2005, 389, 137–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, N.P.; Sansonetti, P.; Mounier, J.; Exley, R.M.; Parsot, C.; Guadagnini, S.; Prevost, M.C.; Prochnicka-Chalufour, A.; Delepierre, M.; Tanguy, M.; et al. Optimization of virulence functions through glucosylation of Shigella LPS. Science 2005, 307, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.J.; Imberty, A.; Phalipon, A.; Perez, S.; Simenel, C.; Mulard, L.A.; Delepierre, M. Conformational studies of the O-specific polysaccharide of Shigella flexneri 5a and of four related synthetic pentasaccharide fragments using NMR and molecular modeling. J. Biol. Chem. 2003, 278, 47928–47936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blocker, A.; Gounon, P.; Larquet, E.; Niebuhr, K.; Cabiaux, V.; Parsot, C.; Sansonetti, P. The tripartite type III secreton of Shigella flexneri inserts IpaB and IpaC into host membranes. J. Cell. Biol. 1999, 147, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Kusmierek, M.; Dersch, P. Regulation of host-pathogen interactions via the post-transcriptional Csr/Rsm system. Curr. Opin. Microbiol. 2018, 41, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Pukkila-Worley, R.; Ausubel, F.M. Immune defense mechanisms in the Caenorhabditis elegans intestinal epithelium. Curr. Opin. Immunol. 2012, 24, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansonetti, P.J. Pathogenesis of shigellosis: From molecular and cellular biology of epithelial cell invasion to tissue inflammation and vaccine development. Jpn. J. Med. Sci. Biol. 1998, 51, S69–S80. [Google Scholar] [CrossRef] [Green Version]
- Bernardes, E.V.T.; Charron-Mazenod, L.; Reading, D.J.; Reckseidler-Zenteno, S.L.; Lewenza, S. Exopolysaccharide-Repressing Small Molecules with Antibiofilm and Antivirulence Activity against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2017, 61, e01997-16. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Pan, W.; Kim, W.; White, A.; Li, S.; Li, H.; Lee, K.; Fuchs, B.B.; Zeng, K.; Mylonakis, E. Caenorhabditis elegans mounts a p38 MAPK pathway-mediated defence to Cutibacterium acnes infection. Cell. Microbiol. 2020, 22, e13234. [Google Scholar] [CrossRef]
- Nystul, T.G.; Roth, M.B. Carbon monoxide-induced suspended animation protects against hypoxic damage in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2004, 101, 9133–9136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, T.T.; Karkho-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Meisel, J.D.; Panda, O.; Mahanti, P.; Schroeder, F.C.; Kim, D.H. Chemosensation of bacterial secondary metabolites modulates neuroendocrine signaling and behavior of C. elegans. Cell 2014, 159, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, R.; Priefer, U.; Puhler, A. A broad host range mobilization system for in vivo genetic engineering transposon mutagenesis in gram negative bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Tan, M.W.; Mahajan-Miklos, S.; Ausubel, F.W. Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to medel mammalian bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.; Taubert, S.; Crawford, D.; Libina, N.; Lee, S.-J.; Kenyon, C. Lifespan extension by conditions that inhibit translation in Caenorhabditis elegans. Aging Cell 2007, 6, 95–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezzoagli, C.; Granato, E.T.; Kummerli, R. In-vivo microscopy reveals the impact of Pseudomonas aeruginosa social interactions on host colonization. ISME J. 2019, 13, 2403–2414. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Version | Isolation Source | Isolation Country | Reference |
|---|---|---|---|---|
| P. donghuensis HYS | NZ_AJJP00000000.1 | Lake | China | Gao et al., 2012 [12] |
| P. vranovensis | NZ_MOAM00000000.1 | River | USA | Tao et al., 2016 [30] |
| Pseudomonas sp. WS 5059 | NZ_JAAQWO010000002.1 | Raw milk | Germany | Maier et al., 2020 [31] |
| P. poae | NZ_PCQN01000008.1 | Rotting apple | Germany | Schulenburg et al., 2017 [32] |
| P. extremaustralis | NZ_AHIP01000006.1 | Temporary water pond in Antarctica | Argentina | Tribelli et al., 2012 [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; Wang, P.; Zhu, X.; Xie, Z. Pseudomonas donghuensis HYS gtrA/B/II Gene Cluster Contributes to Its Pathogenicity toward Caenorhabditis elegans. Int. J. Mol. Sci. 2021, 22, 10741. https://doi.org/10.3390/ijms221910741
Xiao Y, Wang P, Zhu X, Xie Z. Pseudomonas donghuensis HYS gtrA/B/II Gene Cluster Contributes to Its Pathogenicity toward Caenorhabditis elegans. International Journal of Molecular Sciences. 2021; 22(19):10741. https://doi.org/10.3390/ijms221910741
Chicago/Turabian StyleXiao, Yaqian, Panning Wang, Xuesi Zhu, and Zhixiong Xie. 2021. "Pseudomonas donghuensis HYS gtrA/B/II Gene Cluster Contributes to Its Pathogenicity toward Caenorhabditis elegans" International Journal of Molecular Sciences 22, no. 19: 10741. https://doi.org/10.3390/ijms221910741