
A Review of Prostate Organogenesis and a Role for iPSC-Derived Prostate Organoids to Study Prostate Development and Disease

 ,
,
Abstract
:1. Introduction
2. Prostate Organogenesis
2.1. Budding, Elongation, and Branching of the Prostate
2.2. Ductal Canalisation and Cytodifferentiation
3. Key Molecular Drivers of Prostate Development
3.1. Androgen Receptor
3.2. Homeobox Protein NKX3.1
3.3. Fibroblast Growth Factors
3.4. Wingless-Related Integration Sites
3.5. Homeobox Genes
3.6. Bone Morphogenetic Proteins (BMP)
3.7. Sonic Hedgehog (SHH) Signalling
3.8. Forkhead Box A (FOXA)
3.9. Other Factors
4. Prostate Development and Disease
5. PSC-Derived Prostate Tissue for Developmental and Disease Research
6. Summary
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [Green Version]
- Pomerantz, M.M.; Qiu, X.; Zhu, Y.; Takeda, D.Y.; Pan, W.; Baca, S.C.; Gusev, A.; Korthauer, K.D.; Severson, T.M.; Ha, G.; et al. Prostate cancer reactivates developmental epigenomic programs during metastatic progression. Nat. Genet. 2020, 52, 790–799. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, D.; Spring, D.J.; Depinho, R.A. Genetics and biology of prostate cancer. Genes Dev. 2018, 32, 1105–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, F.U.; Ihsan, A.U.; Khan, H.U.; Jana, R.; Wazir, J.; Khongorzul, P.; Waqar, M.; Zhou, X. Comprehensive overview of prostatitis. Biomed. Pharmacother. 2017, 94, 1064–1076. [Google Scholar] [CrossRef]
- Kwon, O.J.; Zhang, L.; Ittmann, M.M.; Xin, L. Prostatic inflammation enhances basal-to-luminal differentiation and accelerates initiation of prostate cancer with a basal cell origin. Proc. Natl. Acad. Sci. USA 2014, 111, E592–E600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toivanen, R.; Mohan, A.; Shen, M.M. Basal Progenitors Contribute to Repair of the Prostate Epithelium Following Induced Luminal Anoikis. Stem Cell Rep. 2016, 6, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Berry, S.J.; Coffey, D.S.; Walsh, P.C.; Ewing, L.L. The development of human benign prostatic hyperplasia with age. J. Urol. 1984, 132, 474–479. [Google Scholar] [CrossRef]
- Strand, D.W.; Costa, D.N.; Francis, F.; Ricke, W.A.; Roehrborn, C.G. Targeting phenotypic heterogeneity in benign prostatic hyperplasia. Differentiation 2017, 96, 49–61. [Google Scholar] [CrossRef]
- Nicholson, T.M.; Ricke, W.A. Androgens and estrogens in benign prostatic hyperplasia: Past, present and future. Differentiation 2011, 82, 184–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacs, J.T.; Coffey, D.S. Etiology and disease process of benign prostatic hyperplasia. Prostate 1989, 15 (Suppl. 2), 33–50. [Google Scholar] [CrossRef]
- Brennen, W.N.; Isaacs, J.T. Mesenchymal stem cells and the embryonic reawakening theory of BPH. Nat. Rev. Urol. 2018, 15, 703–715. [Google Scholar] [CrossRef]
- McNeal, J.E. Origin and evolution of benign prostatic enlargement. Investig. Urol. 1978, 15, 340–345. [Google Scholar]
- Cunha, G.R. Mesenchymal-epithelial interactions: Past, present, and future. Differentiation 2008, 76, 578–586. [Google Scholar] [CrossRef]
- Xu, H.; Tian, Y.; Yuan, X.; Wu, H.; Liu, Q.; Pestell, R.G.; Wu, K. The role of CD44 in epithelial-mesenchymal transition and cancer development. Onco Targets Ther. 2015, 8, 3783–3792. [Google Scholar] [CrossRef] [Green Version]
- Shang, Z.; Cai, Q.; Zhang, M.; Zhu, S.; Ma, Y.; Sun, L.; Jiang, N.; Tian, J.; Niu, X.; Chen, J.; et al. A switch from CD44(+) cell to EMT cell drives the metastasis of prostate cancer. Oncotarget 2015, 6, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ren, D.; Guo, W.; Huang, S.; Wang, Z.; Li, Q.; Du, H.; Song, L.; Peng, X. N-cadherin promotes epithelial-mesenchymal transition and cancer stem cell-like traits via ErbB signaling in prostate cancer cells. Int. J. Oncol. 2016, 48, 595–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Yang, R.; Gao, W.Q. Contributions of epithelial-mesenchymal transition and cancer stem cells to the development of castration resistance of prostate cancer. Mol. Cancer 2014, 13, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, G.R.; Baskin, L. Development of human male and female urogenital tracts. Differentiation 2018, 103, 1–4. [Google Scholar] [CrossRef]
- Cunha, G.R.; Vezina, C.M.; Isaacson, D.; Ricke, W.A.; Timms, B.G.; Cao, M.; Franco, O.; Baskin, L.S. Development of the human prostate. Differentiation 2018, 103, 24–45. [Google Scholar] [CrossRef] [Green Version]
- Toivanen, R.; Shen, M.M. Prostate organogenesis: Tissue induction, hormonal regulation and cell type specification. Development 2017, 144, 1382–1398. [Google Scholar] [CrossRef] [Green Version]
- El-Alfy, M.; Pelletier, G.; Hermo, L.S.; Labrie, F. Unique features of the basal cells of human prostate epithelium. Microsc. Res. Tech. 2000, 51, 436–446. [Google Scholar] [CrossRef]
- Prins, G.S.; Putz, O. Molecular signaling pathways that regulate prostate gland development. Differentiation 2008, 76, 641–659. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Cunha, G.R.; Shen, J.; Cao, M.; Liu, G.; Sinclair, A.; Baskin, L. Development of the human bladder and ureterovesical junction. Differentiation 2018, 103, 66–73. [Google Scholar] [CrossRef]
- Shen, M.M.; Rubin, M.A. Prostate Cancer Research at the Crossroads. Cold Spring Harb. Perspect Med. 2019, 9, a036277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Hayward, S.W.; Cao, M.; Thayer, K.A.; Cunha, G.R. Cell differentiation lineage in the prostate. Differentiation 2001, 68, 270–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sciavolino, P.J.; Abrams, E.W.; Yang, L.; Austenberg, L.P.; Shen, M.M.; Abate-Shen, C. Tissue-specific expression of murine Nkx3.1 in the male urogenital system. Dev. Dyn. 1997, 209, 127–138. [Google Scholar] [CrossRef]
- Marker, P.C.; Donjacour, A.A.; Dahiya, R.; Cunha, G.R. Hormonal, cellular, and molecular control of prostatic development. Dev. Biol. 2003, 253, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Bhatia-Gaur, R.; Donjacour, A.A.; Sciavolino, P.J.; Kim, M.; Desai, N.; Young, P.; Norton, C.R.; Gridley, T.; Cardiff, R.D.; Cunha, G.R.; et al. Roles for Nkx3.1 in prostate development and cancer. Genes Dev. 1999, 13, 966–977. [Google Scholar] [CrossRef] [Green Version]
- Lowsley, O.S. The development of the human prostate gland with reference to the development of other structures at the neck of the urinary bladder. Am. J. Anat. 1912, 13, 299–349. [Google Scholar] [CrossRef]
- Timms, B.G.; Mohs, T.J.; Didio, L.J.A. Ductal budding and branching patterns in the developing prostate. J. Urol. 1994, 155, 1427–1432. [Google Scholar] [CrossRef]
- Donjacour, A.A.; Thomson, A.A.; Cunha, G.R. FGF-10 plays an essential role in the growth of the fetal prostate. Dev. Biol. 2003, 261, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Pu, Y.; Alam, S.; Birch, L.; Prins, G.S. The role of Fgf10 signaling in branching morphogenesis and gene expression of the rat prostate gland: Lobe-specific suppression by neonatal estrogens. Dev. Biol. 2005, 278, 396–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, A.A.; Cunha, G.R. Prostatic growth and development are regulated by FGF10. Development 1999, 126, 3693–3701. [Google Scholar] [CrossRef]
- Kuslak, S.L.; Marker, P.C. Fibroblast growth factor receptor signaling through MEK-ERK is required for prostate bud induction. Differentiation 2007, 75, 638–651. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, F. FGF signalling in prostate development, tissue homoeostasis and tumorigenesis. Biosci. Rep. 2010, 30, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Sugimura, Y.; Foster, B.A.; Hom, Y.K.; Lipschutz, J.H.; Rubin, J.S.; Finch, P.W.; Aaronson, S.A.; Hayashi, N.; Kawamura, J.; Cunha, G.R. Keratinocyte growth factor (KGF) can replace testosterone in the ductal branching morphogenesis of the rat ventral prostate. Int. J. Dev. Biol. 1996, 40, 941–951. [Google Scholar] [PubMed]
- Huang, L.; Pu, Y.; Hu, W.Y.; Birch, L.; Luccio-Camelo, D.; Yamaguchi, T.; Prins, G.S. The role of Wnt5a in prostate gland development. Dev. Biol. 2009, 328, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Allgeier, S.H.; Lin, T.M.; Vezina, C.M.; Moore, R.W.; Fritz, W.A.; Chiu, S.Y.; Zhang, C.L.; Peterson, R.E. WNT5A selectively inhibits mouse ventral prostate development. Dev. Biol. 2008, 324, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Pu, Y.; Huang, L.; Birch, L.; Prins, G.S. Androgen Regulation of Prostate Morphoregulatory Gene Expression: Fgf10-dependent and -independent pathways. Endocrinology 2007, 148, 1697–1706. [Google Scholar] [CrossRef] [Green Version]
- Lamm, M.L.G.; Podlasek, C.A.; Barnett, D.H.; Lee, J.; Clemens, J.Q.; Hebner, C.M.; Bushman, W. Mesenchymal factor bone morphogenetic protein 4 restricts ductal budding and branching morphogenesis in the developing prostate. Dev. Biol. 2001, 232, 301–314. [Google Scholar] [CrossRef] [Green Version]
- Grishina, I.B.; Kim, S.Y.; Ferrara, C.; Makarenkova, H.P.; Walden, P.D. BMP7 inhibits branching morphogenesis in the prostate gland and interferes with Notch signaling. Dev. Biol. 2005, 288, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Freestone, S.H.; Marker, P.; Grace, O.C.; Tomlinson, D.C.; Cunha, G.R.; Harnden, P.; Thomson, A.A. Sonic hedgehog regulates prostatic growth and epithelial differentiation. Dev. Biol. 2003, 264, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Bushman, W. Differential stage-dependent regulation of prostatic epithelial morphogenesis by Hedgehog signaling. Dev. Biol. 2013, 380, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.M.; Desai, N.; Wang, X.; Karhadkar, S.S.; Reynon, M.; Abate-Shen, C.; Beachy, P.A.; Shen, M.M. Roles for Hedgehog signaling in androgen production and prostate ductal morphogenesis. Dev. Biol. 2004, 267, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Hayward, S.W.; Baskin, L.S.; Haughney, P.C.; Foster, B.A.; Cunha, A.R.; Dahiya, R.; Prins, G.S.; Cunha, G.R. Stromal development in the ventral prostate, anterior prostate and seminal vesicle of the rat. Cells Tissues Organs 1996, 155, 94–103. [Google Scholar] [CrossRef]
- Hudson, D.L.; Guy, A.T.; Fry, P.; O’Hare, M.J.; Watt, F.M.; Masters, J.R.W. Epithelial cell differentiation pathways in the human prostate: Identification of intermediate phenotypes by keratin expression. J. Histochem. Cytochem. 2001, 49, 271–278. [Google Scholar] [CrossRef] [Green Version]
- Szczyrba, J.; Niesen, A.; Wagner, M.; Wandernoth, P.M.; Aumüller, G.; Wennemuth, G. Neuroendocrine cells of the prostate derive from the neural crest. J. Biol. Chem. 2017, 292, 2021–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ousset, M.; Van Keymeulen, A.; Bouvencourt, G.; Sharma, N.; Achouri, Y.; Simons, B.D.; Blanpain, C. Multipotent and unipotent progenitors contribute to prostate postnatal development. Nat. Cell Biol. 2012, 14, 1131–1138. [Google Scholar] [CrossRef]
- Hayward, S.W.; Haughney, P.C.; Rosen, M.A.; Greulich, K.M.; Weier, H.U.G.; Dahiya, R.; Cunha, G.R. Interactions between adult human prostatic epithelium and rat urogenital sinus mesenchyme in a tissue recombination model. Differentiation 1998, 63, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.Y.; Leav, I.; Lau, K.M.; Ho, S.M.; Pflueger, S.M.V. Expression of estrogen receptor beta in the fetal, neonatal, and prepubertal human prostate. Prostate 2002, 52, 69–81. [Google Scholar] [CrossRef]
- Pritchard, C.C.; Nelson, P.S. Gene expression profiling in the developing prostate. Differentiation 2008, 76, 624–640. [Google Scholar] [CrossRef] [PubMed]
- Aaron, L.T.; Franco, O.E.; Hayward, S.W. Review of Prostate Anatomy and Embryology and the Etiology of Benign Prostatic Hyperplasia. Urol. Clin. N. Am. 2016, 43, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeal, J.E. The zonal anatomy of the prostate. Prostate 1981, 2, 35–49. [Google Scholar] [CrossRef]
- Francis, J.C.; Swain, A. Prostate organogenesis. Cold Spring Harb. Perspect. Med. 2018, 8, a030353. [Google Scholar] [CrossRef]
- Baulieu, E.E.; Lasnitzki, I.; Robel, P. Metabolism of testosterone and action of metabolites on prostate glands grown in organ culture. Nature 1968, 219, 1155–1156. [Google Scholar] [CrossRef]
- Lyon, M.F.; Hawkes, S.G. X-linked Gene for testicular feminization in the mouse. Nature 1970, 227, 1217–1219. [Google Scholar] [CrossRef]
- Takeda, H.; Lasnitzki, I.; Mizuno, T. Analysis of prostatic bud induction by brief androgen treatment in the fetal rat urogenital sinus. J. Endocrinol. 1986, 110, 467–470. [Google Scholar] [CrossRef]
- Cunha, G.R.; Cao, M.; Franco, O.; Baskin, L.S. A comparison of prostatic development in xenografts of human fetal prostate and human female fetal proximal urethra grown in dihydrotestosterone-treated hosts. Differentiation 2020, 115, 37–52. [Google Scholar] [CrossRef]
- Charest, N.J.; Zhou, Z.x.; Lubahn, D.B.; Olsen, K.L.; Wilson, E.M.; French, F.S. A frameshift mutation destabilizes androgen receptor messenger RNA in the Tfm mouse. Mol. Endocrinol. 1991, 5, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Bardin, C.W.; Bullock, L.P.; Jänne, O.; Jacob, S.T. Genetic regulation of the androgen receptor-A study of testicular feminization in the mouse. J. Steroid Biochem. 1975, 6, 515–520. [Google Scholar] [CrossRef]
- Cunha, G.R.; Lung, B. The possible influence of temporal factors in androgenic responsiveness of urogenital tissue recombinants from wild-type and androgen-insensitive (Tfm) Mice. J. Exp. Zool. 1978, 205, 181–193. [Google Scholar] [CrossRef]
- Donjacour, A.A.A.; Cunha, G.R. Assessment of prostatic protein secretion in tissue recombinants made of urogenital sinus mesenchyme and urothelium from normal or androgen-insensitive mice. Endocrinology 1993, 132, 2342–2350. [Google Scholar] [CrossRef]
- Cunha, G.R.; Battle, E.; Young, P.; Brody, J.; Donjacour, A.; Hayashi, N.; Kinbara, H. Role of epithelial-mesenchymal interactions in the differentiation and spatial organization of visceral smooth muscle. Epithel. Cell Biol. 1992, 1, 76–83. [Google Scholar]
- Thomson, A.A. Mesenchymal mechanisms in prostate organogenesis. Differentiation 2008, 76, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Kruithof-de Julio, M.; Shibata, M.; Desai, N.; Reynon, M.; Halili, M.V.; Hu, Y.P.; Price, S.M.; Abate-Shen, C.; Shen, M.M. Canonical Wnt signaling regulates Nkx3.1 expression and luminal epithelial differentiation during prostate organogenesis. Dev. Dyn. 2013, 242, 1160–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, A.; Le Magnen, C.; Mitrofanova, A.; Ouyang, X.; Califano, A.; Abate-Shen, C. Identification of an NKX3.1-G9a-UTY transcriptional regulatory network that controls prostate differentiation. Science 2016, 352, 1576–1580. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Luo, Y.; Kan, M.; McKeehan, W.L. Fibroblast growth factor-10: A second candidate stromal to epithelial cell andromedin in prostate. J. Biol. Chem. 1999, 274, 12827–12834. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.; Fukabori, Y.; Nikolaropoulos, S.; Wang, F.; McKeehan, W.L. Heparin-binding keratinocyte growth factor is a candidate stromal-to-epithelial-cell andromedin. Mol. Endocrinol. 1992, 6, 2123–2128. [Google Scholar] [CrossRef] [PubMed]
- Finch, P.W.; Cunha, G.R.; Rubin, J.S.; Wong, J.; Ron, D. Pattern of keratinocyte growth factor and keratinocyte growth factor receptor expression during mouse fetal development suggests a role in mediating morphogenetic mesenchymal-epithelial interactions. Dev. Dyn. 1995, 203, 223–240. [Google Scholar] [CrossRef]
- Mehta, V.; Abler, L.L.; Keil, K.P.; Schmitz, C.T.; Joshi, P.S.; Vezina, C.M. Atlas of Wnt and R-spondin gene expression in the developing male mouse lower urogenital tract. Dev. Dyn. 2011, 240, 2548–2560. [Google Scholar] [CrossRef] [Green Version]
- Tanner, M.J.; Welliver, R.C.; Chen, M.; Shtutman, M.; Godoy, A.; Smith, G.; Mian, B.M.; Buttyan, R. Effects of androgen receptor and androgen on gene expression in prostate stromal fibroblasts and paracrine signaling to prostate cancer cells. PLoS ONE 2011, 6, e16027. [Google Scholar] [CrossRef] [PubMed]
- Podlasek, C.A.; Duboule, D.; Bushman, W. Male accessory sex organ morphogenesis is altered by loss of function of Hoxd-13. Dev. Dyn. 1997, 208, 454–465. [Google Scholar] [CrossRef]
- Podlasek, C.A.; Clemens, J.Q.; Bushman, W. HOXA-13 gene mutation results in abnormal seminal vesicle and prostate development. J. Urol. 1999, 161, 1655–1661. [Google Scholar] [CrossRef]
- Podlasek, C.A.; Seo, R.M.; Quentin Clemens, J.; Ma, L.; Maas, R.L.; Bushman, W. Hoxa-10 deficient male mice exhibit abnormal development of the accessory sex organs. Dev. Dyn. 1999, 214, 1–12. [Google Scholar] [CrossRef]
- Meeks, J.J.; Schaeffer, E.M. Genetic regulation of prostate development. J. Androl. 2011, 32, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Economides, K.D.; Capecchi, M.R. Hoxb13 is required for normal differentiation and secretory function of the ventral prostate. Development 2003, 130, 2061–2069. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Pu, Y.; Hepps, D.; Danielpour, D.; Prins, G.S. Posterior Hox Gene Expression and Differential Androgen Regulation in the Developing and Adult Rat Prostate Lobes. Endocrinology 2007, 148, 1235–1245. [Google Scholar] [CrossRef]
- McMullin, R.P.; Dobi, A.; Mutton, L.N.; Orosz, A.; Maheshwari, S.; Shashikant, C.S.; Bieberich, C.J. A FOXA1-binding enhancer regulates Hoxb13 expression in the prostate gland. Proc. Natl. Acad. Sci. USA 2010, 107, 98–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, J.D.; Chang, C.Y.; Wittmann, B.M.; Kunder, R.S.; Cui, H.; Fan, D.; Joseph, J.D.; McDonnell, D.P. The Homeodomain Protein HOXB13 Regulates the Cellular Response to Androgens. Mol. Cell 2009, 36, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Cook, C.; Vezina, C.M.; Allgeier, S.H.; Shaw, A.; Yu, M.; Peterson, R.E.; Bushman, W. Noggin is required for normal lobe patterning and ductal budding in the mouse prostate. Dev. Biol. 2007, 312, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Lamm, M.L.G.; Catbagan, W.S.; Laciak, R.J.; Barnett, D.H.; Hebner, C.M.; Gaffield, W.; Walterhouse, D.; Iannaccone, P.; Bushman, W. Sonic hedgehog activates mesenchymal Gli1 expression during prostate ductal bud formation. Dev. Biol. 2002, 249, 349–366. [Google Scholar] [CrossRef] [Green Version]
- Pu, Y.; Huang, L.; Prins, G.S. Sonic hedgehog-patched Gli signaling in the developing rat prostate gland: Lobe-specific suppression by neonatal estrogens reduces ductal growth and branching. Dev. Biol. 2004, 273, 257–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podlasek, C.A.; Barnett, D.H.; Clemens, J.Q.; Bak, P.M.; Bushman, W. Prostate development requires sonic hedgehog expressed by the urogenital sinus epithelium. Dev. Biol. 1999, 209, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.K.; Taipale, J.; Cooper, M.K.; Beachy, P.A. Inhibition of Hedgehog signaling by direct binding of cyclopamine to Smoothened. Genes Dev. 2002, 16, 2743–2748. [Google Scholar] [CrossRef] [Green Version]
- Mirosevich, J.; Gao, N.; Matusik, R.J. Expression of Foxa transcription factors in the developing and adult murine prostate. Prostate 2005, 62, 339–352. [Google Scholar] [CrossRef]
- Gao, N.; Zhang, J.; Rao, M.A.; Case, T.C.; Mirosevich, J.; Wang, Y.; Jin, R.; Gupta, A.; Rennie, P.S.; Matusik, R.J. The role of hepatocyte nuclear factor-3α (forkhead box A1) and androgen receptor in transcriptional regulation of prostatic genes. Mol. Endocrinol. 2003, 17, 1484–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamat, M.; Chew, K.Y.; Shaw, G.; Renfree, M.B. FOXA1 and SOX9 expression in the developing urogenital sinus of the tammar wallaby (Macropus eugenii). Sex. Dev. 2015, 9, 216–228. [Google Scholar] [CrossRef]
- Wang, D.; Garcia-Bassets, I.; Benner, C.; Li, W.; Su, X.; Zhou, Y.; Qiu, J.; Liu, W.; Kaikkonen, M.U.; Ohgi, K.A.; et al. Reprogramming transcription by distinct classes of enhancers functionally defined by eRNA. Nature 2011, 474, 390–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopachik, W.; Hayward, S.W.; Cunha, G.R. Expression of Hepatocyte Nuclear Factor-3alpha in Rat Prostate, Seminal Vesicle, and Bladder. Dev. Dyn. 1998, 211, 131–140. [Google Scholar] [CrossRef]
- Gao, N.; Ishii, K.; Mirosevich, J.; Kuwajima, S.; Oppenheimer, S.R.; Roberts, R.L.; Jiang, M.; Yu, X.; Shappell, S.B.; Caprioli, R.M.; et al. Forkhead box A1 regulates prostate ductal morphogenesis and promotes epithelial cell maturation. Development 2005, 132, 3431–3443. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, D.C.; Ruiz i Altaba, A.; Chen, W.S.; Hoodless, P.; Prezioso, V.R.; Jessell, T.M.; Darnell, J.E. The winged-helix transcription factor HNF-3β is required for notochord development in the mouse embryo. Cell 1994, 78, 575–588. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, L.; Wu, X.; Li, R.; Wen, J.; Sha, J.; Wen, X. The PI3K/AKT pathway in the pathogenesis of prostate cancer. Front. Biosci. (Landmark Ed.) 2016, 21, 1084–1091. [Google Scholar] [CrossRef] [Green Version]
- Risbridger, G.P.; Toivanen, R.; Taylor, R.A. Preclinical models of prostate cancer: Patient-derived xenografts, organoids, and other explant models. Cold Spring Harb. Perspect. Med. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, G.R. Epithelio-mesenchymal interactions in primordial glandstructures which become responsive to androgenic stimulation. Anat. Rec. 1972, 172, 179–195. [Google Scholar] [CrossRef]
- Taylor, R.A.; Cowin, P.A.; Cunha, G.R.; Pera, M.; Trounson, A.O.; Pedersen, J.; Risbridger, G.P. Formation of human prostate tissue from embryonic stem cells. Nat. Methods 2006, 3, 179–181. [Google Scholar] [CrossRef]
- Ferreira, C.; Lobo, J.; Antunes, L.; Lopes, P.; Jeronimo, C.; Henrique, R. Differential expression of E-cadherin and P-cadherin in pT3 prostate cancer: Correlation with clinical and pathological features. Virchows Arch. 2018, 473, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashley, G.R.; Grace, C.O.; Vanpoucke, C.; Thomson, A.A. Identification of EphrinB1 expression in prostatic mesenchyme and a role for EphB-EphrinB signalling in prostate development. Differentiation 2010, 80, 89–98. [Google Scholar] [CrossRef]
- Lv, L.; Yang, Z.; Ma, T.; Xuan, Y. Gli1, a potential cancer stem cell marker, is strongly associated with prognosis in prostate cancer. Int. J. Clin. Exp. Pathol. 2018, 11, 4957–4966. [Google Scholar]
- Xia, L.; Bouamar, H.; Gu, X.; Zeballos, C.; Qin, T.; Wang, B.; Zhou, Y.; Wang, Y.; Yang, J.; Zhu, H.; et al. Gli2 mediates the development of castrationresistant prostate cancer. Int. J. Oncol. 2020, 57, 100–112. [Google Scholar] [CrossRef]
- Shiota, M.; Bishop, J.L.; Nip, K.M.; Zardan, A.; Takeuchi, A.; Cordonnier, T.; Beraldi, E.; Bazov, J.; Fazli, L.; Chi, K.; et al. Hsp27 regulates epithelial mesenchymal transition, metastasis, and circulating tumor cells in prostate cancer. Cancer Res. 2013, 73, 3109–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciocca, D.R.; Fanelli, M.A.; Cuello-Carrión, F.D.; Castro, G.N. Heat shock proteins in prostate cancer: From tumorigenesis to the clinic. Int. J. Hyperth. 2010, 26, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Culig, Z.; Puhr, M. Interleukin-6 and prostate cancer: Current developments and unsolved questions. Mol. Cell. Endocrinol. 2018, 462, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Dahl, H.C.; Kanchwala, M.; Thomas-Jardin, S.E.; Sandhu, A.; Kanumuri, P.; Nawas, A.F.; Xing, C.; Lin, C.; Frigo, D.E.; Delk, N.A. Chronic IL-1 exposure drives LNCaP cells to evolve androgen and AR independence. PLoS ONE 2020, 15, e0242970. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, Y.; Ling, Z.; Li, Y.; Ren, X.; Yang, J.; Wang, Z.; Xia, J.; Zhang, W.; Cheng, B. R-spondin 2-LGR4 system regulates growth, migration and invasion, epithelial-mesenchymal transition and stem-like properties of tongue squamous cell carcinoma via Wnt/beta-catenin signaling. EBioMedicine 2019, 44, 275–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Dong, B.; Yan, Y.; Hu, G.; Xu, Y. Association between MMP-2 expression and prostate cancer: A meta-analysis. Biomed. Rep. 2015, 4, 241–245. [Google Scholar] [CrossRef]
- Quigley, D.A.; Dang, H.X.; Zhao, S.G.; Lloyd, P.; Aggarwal, R.; Alumkal, J.J.; Foye, A.; Kothari, V.; Perry, M.D.; Bailey, A.M.; et al. Genomic Hallmarks and Structural Variation in Metastatic Prostate Cancer. Cell 2018, 174, 758–769.e759. [Google Scholar] [CrossRef] [Green Version]
- Samoylenko, O.A.; Stakhovsky, E.O.; Vitruk, Y.V.; Shlyakhovenko, V.O. Ornithine decarboxylase activity in prostate cancer. Exp. Oncol. 2021, 43, 46–51. [Google Scholar]
- Khoubehi, B.; Kessling, A.M.; Adshead, J.M.; Smith, G.L.; Smith, R.D.; Ogden, C.W. Expression of the developmental and oncogenic PAX2 gene in human prostate cancer. J. Urol. 2001, 165, 2115–2120. [Google Scholar] [CrossRef]
- Downing, S.R.; Russell, P.J.; Jackson, P. Alterations of p53 are common in early stage prostate cancer. Can. J. Urol. 2003, 10, 1924–1933. [Google Scholar]
- Dhillon, P.K.; Barry, M.; Stampfer, M.J.; Perner, S.; Fiorentino, M.; Fornari, A.; Ma, J.; Fleet, J.; Kurth, T.; Rubin, M.A.; et al. Aberrant cytoplasmic expression of p63 and prostate cancer mortality. Cancer Epidemiol. Biomark. Prev. 2009, 18, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Shen, M.; Hsu, E.-C.; Zhang, C.A.; Garcia-Marques, F.; Nolley, R.; Koul, K.; Rice, M.A.; Aslan, M.; Pitteri, S.J.; et al. Discovery of PTN as a serum-based biomarker of pro-metastatic prostate cancer. Br. J. Cancer 2020, 124, 896–900. [Google Scholar] [CrossRef]
- Wise, H.M.; Hermida, M.A.; Leslie, N.R. Prostate cancer, PI3K, PTEN and prognosis. Clin. Sci. (Lond.) 2017, 131, 197–210. [Google Scholar] [CrossRef]
- Mesci, A.; Lucien, F.; Huang, X.; Wang, E.H.; Shin, D.; Meringer, M.; Hoey, C.; Ray, J.; Boutros, P.C.; Leong, H.S.; et al. RSPO3 is a prognostic biomarker and mediator of invasiveness in prostate cancer. J. Transl. Med. 2019, 17, 125. [Google Scholar] [CrossRef]
- Zheng, L.; Sun, D.; Fan, W.; Zhang, Z.; Li, Q.; Jiang, T. Diagnostic Value of SFRP1 as a Favorable Predictive and Prognostic Biomarker in Patients with Prostate Cancer. PLoS ONE 2015, 10, e0118276. [Google Scholar] [CrossRef] [PubMed]
- Francis, J.; Capper, A.; Ning, J.; Knight, E.; De Bono, J.; Swain, A. SOX9 is a driver of aggressive prostate cancer by promoting invasion, cell fate and cytoskeleton alterations and epithelial to mesenchymal transition. Oncotarget 2018, 9, 7604–7615. [Google Scholar] [CrossRef] [Green Version]
- Bret, C.; Moreaux, J.; Schved, J.-F.; Hose, D.; Klein, B. SULFs in human neoplasia: Implication as progression and prognosis factors. J. Transl. Med. 2011, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Ahel, J.; Hudorovic, N.; Vicic-Hudorovic, V.; Nikles, H. Tgf-Beta in the Natural History of Prostate Cancer. Acta Clin. Croat 2019, 58, 128–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, J.M.; Heinlein, C.; Kim, T.; Hernandez, S.A.; Malik, M.S.; True, L.D.; Morrissey, C.; Corey, E.; Montgomery, B.; Mostaghel, E.; et al. The androgen-regulated protease TMPRSS2 activates a proteolytic cascade involving components of the tumor microenvironment and promotes prostate cancer metastasis. Cancer Discov. 2014, 4, 1310–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; De, S.; Brainard, J.; Byzova, T.V. Metastatic properties of prostate cancer cells are controlled by VEGF. Cell Commun. Adhes 2004, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Singhal, U.; Qiao, Y.; Kasputis, T.; Chung, J.-S.; Zhao, H.; Chammaa, F.; Belardo, J.A.; Roth, T.M.; Zhang, H.; et al. Wnt Signaling Drives Prostate Cancer Bone Metastatic Tropism and Invasion. Transl. Oncol. 2020, 13, 100747. [Google Scholar] [CrossRef]
- Barclay, W.W.; Woodruff, R.D.; Hall, M.C.; Cramer, S.D. A system for studying epithelial-stromal interactions reveals distinct inductive abilities of stromal cells from benign prostatic hyperplasia and prostate cancer. Endocrinology 2005, 146, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Dhanasekaran, S.M.; Dash, A.; Yu, J.; Maine, I.P.; Laxman, B.; Tomlins, S.A.; Creighton, C.J.; Menon, A.; Rubin, M.A.; Chinnaiyan, A.M. Molecular profiling of human prostate tissues: Insights into gene expression patterns of prostate development during puberty. FASEB J. 2005, 19, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, B.; Riddick, A.C.; Stewart, G.D.; Anderson, R.A.; Franco, O.E.; Hayward, S.W.; Thomson, A.A. Identification of stromally expressed molecules in the prostate by tag-profiling of cancer-associated fibroblasts, normal fibroblasts and fetal prostate. Oncogene 2012, 31, 1130–1142. [Google Scholar] [CrossRef] [Green Version]
- Nash, C.; Boufaied, N.; Badescu, D.; Wang, Y.C.; Paliouras, M.; Trifiro, M.; Ragoussis, I.; Thomson, A.A. Genome-wide analysis of androgen receptor binding and transcriptomic analysis in mesenchymal subsets during prostate development. DMM Dis. Models Mech. 2019, 12, dmm039297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, E.M.; Marchionni, L.; Huang, Z.; Simons, B.; Blackman, A.; Yu, W.; Parmigiani, G.; Berman, D.M. Androgen-induced programs for prostate epithelial growth and invasion arise in embryogenesis and are reactivated in cancer. Oncogene 2008, 27, 7180–7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, C.; Mecham, B.; Dumpit, R.; Coleman, U.; Bhattacharjee, M.; Chen, Q.; Sikes, R.A.; Nelson, P.S. Conserved gene expression programs integrate mammalian prostate development and tumorigenesis. Cancer Res. 2009, 69, 1739–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Skoog, L.; Hellborg, H.; Jonstam, G.; Wingmo, I.L.; Hjalm-Eriksson, M.; Harmenberg, U.; Cedermark, G.C.; Andersson, K.; Ahrlund-Richter, L.; et al. An expression signature at diagnosis to estimate prostate cancer patients’ overall survival. Prostate Cancer Prostatic Dis. 2014, 17, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef]
- Hepburn, A.; Curry, E.; Moad, M.; Steele, R.; Franco, O.; Wilson, L.; Singh, P.; Crawford, S.; Gaughan, L.; Mills, I.; et al. High-throughput propagation of human prostate tissue from induced-pluripotent stem cells. bioRxiv 2019. [Google Scholar] [CrossRef]
- Keil, K.P.; Abler, L.L.; Mehta, V.; Altmann, H.M.; Laporta, J.; Plisch, E.H.; Suresh, M.; Hernandez, L.L.; Vezina, C.M. DNA methylation of E-cadherin is a priming mechanism for prostate development. Dev. Biol. 2014, 387, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Jerde, T.J.; Bushman, W. IL-1 induces IGF-dependent epithelial proliferation in prostate development and reactive hyperplasia. Sci. Signal. 2009, 2, ra49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadi, M.V.; Walsh, P.C.; Barrack, E.R. Immunohistochemical study of androgen receptors in metastatic prostate cancer. Comparison of receptor content and response to hormonal therapy. Cancer 1991, 67, 3057–3064. [Google Scholar] [CrossRef]
- Ruizeveld de Winter, J.A.; Janssen, P.J.; Sleddens, H.M.; Verleun-Mooijman, M.C.; Trapman, J.; Brinkmann, A.O.; Santerse, A.B.; Schroder, F.H.; van der Kwast, T.H. Androgen receptor status in localized and locally progressive hormone refractory human prostate cancer. Am. J. Pathol. 1994, 144, 735–746. [Google Scholar]
- Attard, G.; Swennenhuis, J.F.; Olmos, D.; Reid, A.H.; Vickers, E.; A’Hern, R.; Levink, R.; Coumans, F.; Moreira, J.; Riisnaes, R.; et al. Characterization of ERG, AR and PTEN gene status in circulating tumor cells from patients with castration-resistant prostate cancer. Cancer Res. 2009, 69, 2912–2918. [Google Scholar] [CrossRef] [Green Version]
- Stein, M.E.; Bernstein, Z.; Abacioglu, U.; Sengoz, M.; Miller, R.C.; Meirovitz, A.; Zouhair, A.; Freixa, S.V.; Poortmans, P.H.; Ash, R.; et al. Small cell (neuroendocrine) carcinoma of the prostate: Etiology, diagnosis, prognosis, and therapeutic implications—A retrospective study of 30 patients from the rare cancer network. Am. J. Med. Sci. 2008, 336, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Gillard, M.; Javier, R.; Ji, Y.; Zheng, S.L.; Xu, J.; Brendler, C.B.; Crawford, S.E.; Pierce, B.L.; Griend, D.J.V.; Franco, O.E. Elevation of Stromal-Derived Mediators of Inflammation Promote Prostate Cancer Progression in African-American Men. Cancer Res. 2018, 78, 6134–6145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiro, N.; Fernandez-Gomez, J.; Sacristan, R.; Fernandez-Garcia, B.; Lobo, B.; Gonzalez-Suarez, J.; Quintas, A.; Escaf, S.; Vizoso, F.J. Stromal factors involved in human prostate cancer development, progression and castration resistance. J. Cancer Res. Clin. Oncol. 2017, 143, 351–359. [Google Scholar] [CrossRef]
- Memarzadeh, S.; Xin, L.; Mulholland, D.J.; Mansukhani, A.; Wu, H.; Teitell, M.A.; Witte, O.N. Enhanced Paracrine FGF10 Expression Promotes Formation of Multifocal Prostate Adenocarcinoma and an Increase in Epithelial Androgen Receptor. Cancer Cell 2007, 12, 572–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolia, A.; Cieslik, M.; Chu, S.C.; Xiao, L.; Ouchi, T.; Zhang, Y.; Wang, X.; Vats, P.; Cao, X.; Pitchiaya, S.; et al. Distinct structural classes of activating FOXA1 alterations in advanced prostate cancer. Nature 2019, 571, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.; Zhou, S.; Cai, C.; Lupien, M.; He, H.H. Pioneer of prostate cancer: Past, present and the future of FOXA1. Protein Cell 2021, 12, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, Y.; de Velasco, M.A.; Oki, T.; Shimizu, N.; Nozawa, M.; Yoshimura, K.; Yoshikawa, K.; Nishio, K.; Uemura, H. HOXA10 expression profiling in prostate cancer. Prostate 2019, 79, 554–563. [Google Scholar] [CrossRef]
- Long, Z.; Li, Y.; Gan, Y.; Zhao, D.; Wang, G.; Xie, N.; Lovnicki, J.M.; Fazli, L.; Cao, Q.; Chen, K.; et al. Roles of the HOXA10 gene during castrate-resistant prostate cancer progression. Endocr. Relat Cancer 2019, 26, 279–292. [Google Scholar] [CrossRef]
- Warot, X.; Fromental-Ramain, C.; Fraulob, V.; Chambon, P.; Dollé, P. Gene dosage-dependent effects of the Hoxa-13 and Hoxd-13 mutations on morphogenesis of the terminal parts of the digestive and urogenital tracts. Development 1997, 124, 4781–4791. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Cai, Y.; Liu, B.; Jiao, X.; Li, Z.T.; Guo, D.Y.; Li, X.W.; Wang, Y.J.; Yang, D.K. HOXA13 is associated with unfavorable survival and acts as a novel oncogene in prostate carcinoma. Future Oncol. 2017, 13, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Ewing, C.M.; Ray, A.M.; Lange, E.M.; Zuhlke, K.A.; Robbins, C.M.; Tembe, W.D.; Wiley, K.E.; Isaacs, S.D.; Johng, D.; Wang, Y.; et al. Germline mutations in HOXB13 and prostate-cancer risk. N. Engl. J. Med. 2012, 366, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouhtit, A.; Al-Kindi, M.N.; Kumar, P.R.; Gupta, I.; Shanmuganathan, S.; Tamimi, Y. Hoxb13, a potential prognostic biomarker for prostate cancer. Front. Biosci. (Elite Ed.) 2016, 8, 40–45. [Google Scholar] [CrossRef]
- Le Magnen, C.; Virk, R.K.; Dutta, A.; Kim, J.Y.; Panja, S.; Lopez-Bujanda, Z.A.; Califano, A.; Drake, C.G.; Mitrofanova, A.; Abate-Shen, C. Cooperation of loss of NKX3.1 and inflammation in prostate cancer initiation. Dis. Model. Mech. 2018, 11, dmm035139. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.C.; Yu-Lee, L.Y.; Lin, S.H. Osteoblastic Factors in Prostate Cancer Bone Metastasis. Curr. Osteoporos. Rep. 2018, 16, 642–647. [Google Scholar] [CrossRef]
- Lee, Y.C.; Cheng, C.J.; Bilen, M.A.; Lu, J.F.; Satcher, R.L.; Yu-Lee, L.Y.; Gallick, G.E.; Maity, S.N.; Lin, S.H. BMP4 promotes prostate tumor growth in bone through osteogenesis. Cancer Res. 2011, 71, 5194–5203. [Google Scholar] [CrossRef] [Green Version]
- Madueke, I.C.; Hu, W.-y.; Huang, L.; Prins, G.S. WNT2 is necessary for normal prostate gland cyto-differentiation and modulates prostate growth in an FGF10 dependent manner. Am. J. Clin. Exp. Urol. 2018, 6, 154–163. [Google Scholar]
- Katoh, M. Frequent up-regulation of WNT2 in primary gastric cancer and colorectal cancer. Int. J. Oncol. 2001, 19, 1003–1007. [Google Scholar] [CrossRef]
- Lee, G.T.; Kwon, S.J.; Kim, J.; Kwon, Y.S.; Lee, N.; Hong, J.H.; Jamieson, C.; Kim, W.J.; Kim, I.Y. WNT5A induces castration-resistant prostate cancer via CCL2 and tumour-infiltrating macrophages. Br. J. Cancer 2018, 118, 670–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, I.; Quintela-Vieira, F.; Lobo, J.; Moreira-Barbosa, C.; Menezes, F.D.; Martins, A.T.; Oliveira, J.; Silva, R.; Jeronimo, C.; Henrique, R. Expression of EMT-Related Genes CAMK2N1 and WNT5A is increased in Locally Invasive and Metastatic Prostate Cancer. J. Cancer 2019, 10, 5915–5925. [Google Scholar] [CrossRef]
- Madueke, I.; Hu, W.Y.; Hu, D.; Swanson, S.M.; Vander Griend, D.; Abern, M.; Prins, G.S. The role of WNT10B in normal prostate gland development and prostate cancer. Prostate 2019, 79, 1692–1704. [Google Scholar] [CrossRef]
- Lin, T.M.; Rasmussen, N.T.; Moore, R.W.; Albrecht, R.M.; Peterson, R.E. Region-specific inhibition of prostatic epithelial bud formation in the urogenital sinus of C57BL/6 mice exposed in utero to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Sci. 2003, 76, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Branam, A.M.; Davis, N.M.; Moore, R.W.; Schneider, A.J.; Vezina, C.M.; Peterson, R.E. TCDD inhibition of canonical wnt signaling disrupts prostatic bud formation in mouse urogenital sinus. Toxicol. Sci. 2013, 133, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Roe, J.S.; Hwang, C.I.; Somerville, T.D.D.; Milazzo, J.P.; Lee, E.J.; Da Silva, B.; Maiorino, L.; Tiriac, H.; Young, C.M.; Miyabayashi, K.; et al. Enhancer Reprogramming Promotes Pancreatic Cancer Metastasis. Cell 2017, 170, 875–888.e820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacs, J.T. Prostate stem cells and benign prostatic hyperplasia. Prostate 2008, 68, 1025–1034. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, G.H.; Malewska, A.; Joseph, D.B.; Malladi, V.S.; Lee, J.; Torrealba, J.; Mauck, R.J.; Gahan, J.C.; Raj, G.V.; Roehrborn, C.G.; et al. A Cellular Anatomy of the Normal Adult Human Prostate and Prostatic Urethra. Cell Rep. 2018, 25, 3530–3542.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 12 October 2021).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyuga, T.; Alcantara, M.; Kajioka, D.; Haraguchi, R.; Suzuki, K.; Miyagawa, S.; Kojima, Y.; Hayashi, Y.; Yamada, G. Hedgehog Signaling for Urogenital Organogenesis and Prostate Cancer: An Implication for the Epithelial–Mesenchyme Interaction (EMI). Int. J. Mol. Sci. 2019, 21, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.A.; Zhao, J.; Andarmani, S.; Kakitani, M.; Oshima, T.; Binnerts, M.E.; Abo, A.; Tomizuka, K.; Funk, W.D. R-Spondin proteins: A novel link to beta-catenin activation. Cell Cycle 2006, 5, 23–26. [Google Scholar] [CrossRef]
- Chaudhry, F.; Isherwood, J.; Bawa, T.; Patel, D.; Gurdziel, K.; Lanfear, D.E.; Ruden, D.M.; Levy, P.D. Single-Cell RNA Sequencing of the Cardiovascular System: New Looks for Old Diseases. Front. Cardiovasc. Med. 2019, 6, 173. [Google Scholar] [CrossRef]
- Nagle, R.B.; Knox, J.D.; Wolf, C.; Bowden, G.T.; Cress, A.E. Adhesion molecules, extracellular matrix, and proteases in prostate carcinoma. J. Cell Biochem. Suppl. 1994, 19, 232–237. [Google Scholar]
- Yu, Y.; Jiang, W. Pluripotent stem cell differentiation as an emerging model to study human prostate development. Stem Cell Res. Ther. 2020, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Lindstrom, N.O.; Ransick, A.; De Sena Brandine, G.; Guo, Q.; Kim, A.D.; Der, B.; Peti-Peterdi, J.; Smith, A.D.; Thornton, M.; et al. In Vivo Developmental Trajectories of Human Podocyte Inform In Vitro Differentiation of Pluripotent Stem Cell-Derived Podocytes. Dev. Cell 2019, 50, 102–116.e106. [Google Scholar] [CrossRef]
- Kanton, S.; Boyle, M.J.; He, Z.; Santel, M.; Weigert, A.; Sanchis-Calleja, F.; Guijarro, P.; Sidow, L.; Fleck, J.S.; Han, D.; et al. Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature 2019, 574, 418–422. [Google Scholar] [CrossRef]
- Hepburn, A.C.; Curry, E.L.; Moad, M.; Steele, R.E.; Franco, O.E.; Wilson, L.; Singh, P.; Buskin, A.; Crawford, S.E.; Gaughan, L.; et al. Propagation of human prostate tissue from induced pluripotent stem cells. Stem Cells Transl. Med. 2020, 9, 734–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepburn, A.C.; Sims, C.H.C.; Buskin, A.; Heer, R. Engineering Prostate Cancer from Induced Pluripotent Stem Cells-New Opportunities to Develop Preclinical Tools in Prostate and Prostate Cancer Studies. Int. J. Mol. Sci. 2020, 21, 905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moad, M.; Pal, D.; Hepburn, A.C.; Williamson, S.C.; Wilson, L.; Lako, M.; Armstrong, L.; Hayward, S.W.; Franco, O.E.; Cates, J.M.; et al. A novel model of urinary tract differentiation, tissue regeneration, and disease: Reprogramming human prostate and bladder cells into induced pluripotent stem cells. Eur. Urol. 2013, 64, 753–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
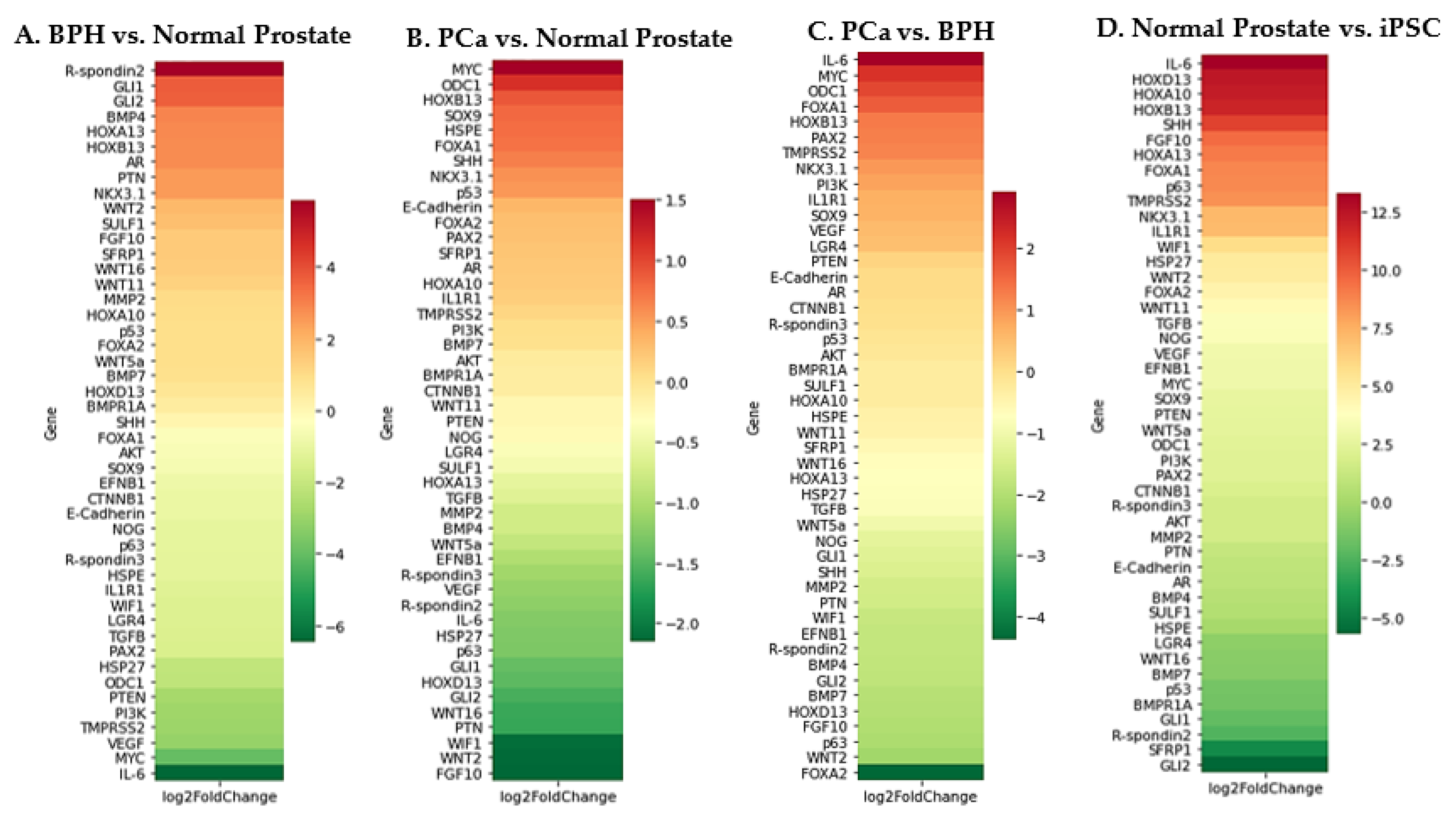{kind=link}
{kind=link}
| Molecules of Interest | Development | Refs | Cancer | Refs |
|---|---|---|---|---|
| AR | Essential for prostate development at all stages; no prostate development in androgen/AR absence | [57,61,62] | Expressed throughout the course of the disease, except in neuroendocrine tumours | [132,133,134,135] |
| FGF7 | Upregulation in rat P0 VP explant cultures resulted in increased budding and branching; Inhibition resulted in decreased branching and epithelial growth | [35,36] | Has been associated with prostate cancer progression; Highly expressed in fibroblasts from localised tumours | [136,137] |
| FGF10 | Upregulation in rat P0 VP explant cultures resulted in epithelial proliferation, branching, and differentiation; Null mutation resulted in reduced budding, branching, and differentiation in UGS explant cultures | [31,32,33,34] | Enhanced mesenchymal expression of FGF10 leads to the formation of prostate cancer | [138] |
| FOXA1 | Null mutation in mice resulted in incomplete lumenization with reduced basal cell hyperplasia and luminal secretory cells | [87,89,90] | Some mutations in FOXA1 in early prostate cancer resulted in enhanced chromatin mobility and activation of a luminal androgen receptor (AR) program of prostate oncogenesis; Other FOXA1 mutations acquired in metastatic prostate cancers; Resulted in dominant chromatin binding by increasing DNA affinity, promoting WNT-pathway driven metastasis; Duplications and translocations in metastatic prostate cancers, within the FOXA1 locus, resulted in overexpression of FOXA1 or other oncogenes | [139,140] |
| HOXA10 | Null mouse mutation resulted in decreased AP size and branching | [74,77] | Evidence of tumor suppressive roles for HOXA10 in the context of prostate cancer; Downregulation of HOXA10 gene expression resulted in PCa cell growth and tumor progression to castrate-resistant stage | [141,142] |
| HOXA13 | Null mouse mutation resulted in decreased DLP and VP size and branching;Double null mutations of Hoxd13 and Hoxa13 resulted in absence of AP and reduced epithelial proliferation | [74,77,143] | HOXA13 is an oncogene for prostate cancer and its overexpression resulted in prostate carcinoma tissues | [144] |
| HOXB13 | Null mouse mutation resulted in loss of VP secretory proteins and abnormal VP luminal cells | [76,77] | Mutations in HOXB13 resulted in significantly increased risk of hereditary prostate cancer; HOXB13 overexpression resulted in during malignant progression of the prostatic tissue and is suspected to contribute to the pathogenesis of the prostate gland | [145,146] |
| NKX3.1 | Null mutant mice had decreased branching, lumenization and prostate secretory product production, and defects in cytodifferentiation with epithelial hyperplasia | [28,34] | Loss of function of NKX3.1 accelerated inflammation-driven prostate cancer initiation potentially via aberrant cellular plasticity and impairment of cellular differentiation | [147] |
| BMP4 | Upregulation in mouse UGS explant cultures reduced epithelial proliferation and branching; Deficiency (Bmp4+/−) in mice led to increased branching | [34,40] | Involved in prostate tumour growth in bone and bone metastasis | [148,149] |
| BMP7 | Upregulation in mouse UGS reduced budding and branching; Null mouse mutation resulted in increased branching | [34,41] | Acted as inhibitor of prostate cancer bone metastasis | [41] |
| WNT2 (canonical) | Upregulation in rat P0 VP explant cultures resulted in decreased size; Null mouse mutant UGS renal grafts led to defective luminal cell differentiation | [150] | Overexpressed in prostate cancer | [151] |
| WNT5A (non-canonical) | Null mouse mutation led to defects in bud positioning in UGS explant cultures, but development proceeded; Upregulation in mouse UGS explant cultures led to reduced budding, ductal elongation, epithelial proliferation, and branching | [37,38,39] | WNT5A was overexpressed in locally invasive and metastatic prostate cancer; WNT5A may be a key gene that induces CRPC in the bone niche | [152,153] |
| WNT10B (canonical) | Upregulation in rat VP (P0) led to decreased ductal elongation and branching | [70,154] | Decreased WNT10B levels in localized cancer let to a hyperproliferative state, whereas increased levels in advanced disease conferred a stemness and malignant propensity due to activation of epithelial to mesenchymal transition genes | [154] |
| RSPO2 and RSPO3 | Inhibition in mouse UGS explant cultures led to reduced and mis-positioned budding with the complete absence of budding in the VP | [70,155,156] | Lower RSPO3 expression resulted in greater metastatic capacity and invasiveness | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buskin, A.; Singh, P.; Lorenz, O.; Robson, C.; Strand, D.W.; Heer, R. A Review of Prostate Organogenesis and a Role for iPSC-Derived Prostate Organoids to Study Prostate Development and Disease. Int. J. Mol. Sci. 2021, 22, 13097. https://doi.org/10.3390/ijms222313097
Buskin A, Singh P, Lorenz O, Robson C, Strand DW, Heer R. A Review of Prostate Organogenesis and a Role for iPSC-Derived Prostate Organoids to Study Prostate Development and Disease. International Journal of Molecular Sciences. 2021; 22(23):13097. https://doi.org/10.3390/ijms222313097
Chicago/Turabian StyleBuskin, Adriana, Parmveer Singh, Oliver Lorenz, Craig Robson, Douglas W. Strand, and Rakesh Heer. 2021. "A Review of Prostate Organogenesis and a Role for iPSC-Derived Prostate Organoids to Study Prostate Development and Disease" International Journal of Molecular Sciences 22, no. 23: 13097. https://doi.org/10.3390/ijms222313097