
miR-22-3p and miR-30e-5p Are Associated with Prognosis in Cervical Squamous Cell Carcinoma

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Differential miRNA Profiles on miRNA Microarray
2.2. Validation of miRNA Expression by qRT-PCR
2.3. Association between miRNA Expression and Clinicopathologic Prognostic Factors
2.4. HPV Infection Pattern and Its Association with miRNA Expression
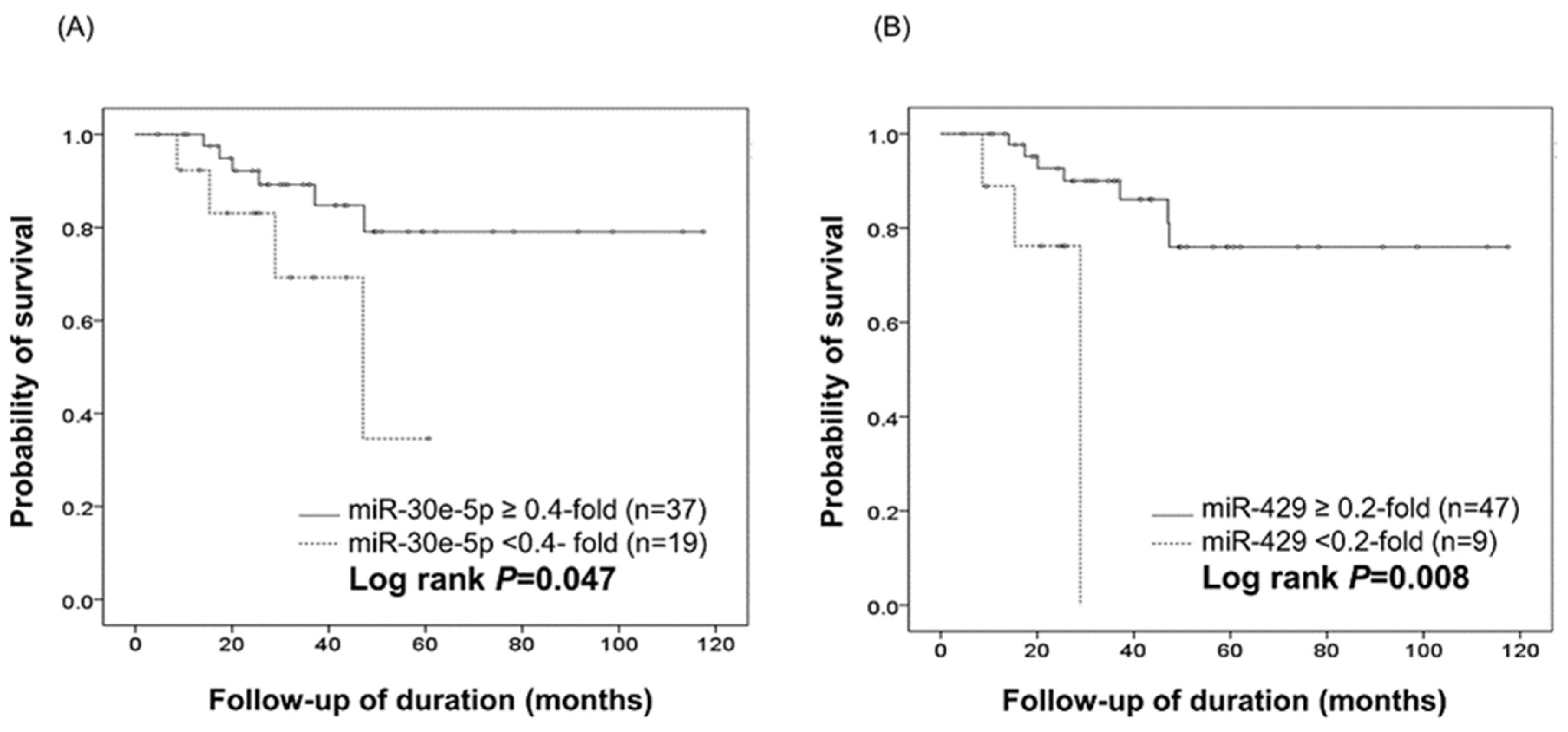
2.5. Survival and Multivariate Cox Analysis
3. Discussion
4. Materials and Methods
4.1. Cervical Tissue Specimens
4.2. HPV Genotyping
4.3. MiRNA Extraction and Microarray Hybridization
4.4. MiRNA Target Prediction and Functional Annotation Analysis
4.5. Validation by qRT-PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Cancer Society. Cancer Facts & Figures 2015. Atlanta: American Cancer Society. 2015. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures.html (accessed on 13 November 2015).
- Saavedra, K.P.; Brebi, P.M.; Roa, J.C. Epigenetic alterations in preneoplastic and neoplastic lesions of the cervix. Clin. Epigenet. 2012, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.I.; Shiekhattar, R. MicroRNA biogenesis and cancer. Cancer Res. 2005, 65, 3509–3512. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, P.S. Cancer genomics: Small RNAs with big impacts. Nature 2005, 435, 745–746. [Google Scholar] [CrossRef]
- Bagga, S.; Bracht, J.; Hunter, S.; Massirer, K.; Holtz, J.; Eachus, R.; Pasquinelli, A.E. Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation. Cell 2005, 122, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.Y.; Hayes, J.J.; Pruss, D.; Wolffe, A.P. A positive role for histone acetylation in transcription factor access to nucleosomal DNA. Cell 1993, 72, 73–84. [Google Scholar] [CrossRef]
- Lee, J.W.; Choi, C.H.; Choi, J.J.; Park, Y.A.; Kim, S.J.; Hwang, S.Y.; Kim, W.Y.; Kim, T.J.; Lee, J.H.; Kim, B.G.; et al. Altered MicroRNA expression in cervical carcinomas. Clin. Cancer Res. 2008, 14, 2535–2542. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.M.; Marques, J.P.; Soares, A.R.; Carreto, L.; Santos, M.A. MicroRNA expression variability in human cervical tissues. PLoS ONE 2010, 5, e11780. [Google Scholar] [CrossRef] [Green Version]
- Rao, Q.; Shen, Q.; Zhou, H.; Peng, Y.; Li, J.; Lin, Z. Aberrant microRNA expression in human cervical carcinomas. Med. Oncol. 2012, 29, 1242–1248. [Google Scholar] [CrossRef]
- Villegas-Ruiz, V.; Juárez-Méndez, S.; Pérez-González, O.A.; Arreola, H.; Paniagua-García, L.; Parra-Melquiadez, M.; Peralta-Rodríguez, R.; López-Romero, R.; Monroy-García, A.; Mantilla-Morales, A.; et al. Heterogeneity of microRNAs expression in cervical cancer cells: Over-expression of miR-196a. Int. J. Clin. Exp. Pathol. 2014, 7, 1389–1401. [Google Scholar]
- Lopez-Camarillo, C.; Marchat, L.A.; Arechaga-Ocampo, E.; Perez-Plasencia, C.; Moral-Hernandez, O.D.; Castaneda-Ortiz, E.J.; Rodriguez-Cuevas, S. MetastamiRs: Non-coding MicroRNAs driving cancer invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 1347–1379. [Google Scholar] [CrossRef]
- Kwak, P.B.; Iwasaki, S.; Tomari, Y. The microRNA pathway and cancer. Cancer Sci. 2010, 101, 2309–2315. [Google Scholar] [CrossRef]
- Hurst, D.R.; Edmonds, M.D.; Welch, D.R. Metastamir: The field of metastasis-regulatory microRNA is spreading. Cancer Res. 2009, 69, 7495–7498. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [Green Version]
- Bosch, F.X.; Manos, M.M.; Muñoz, N.; Sherman, M.; Jansen, A.M.; Peto, J.; Schiffman, M.H.; Moreno, V.; Kurman, R.; Shan, K.V. Prevalence of human papillomavirus in cervical cancer: A worldwide perspective. International biological study on cervical cancer (IBSCC) Study Group. J. Natl. Cancer Inst. 1995, 87, 796–802. [Google Scholar] [CrossRef]
- Boyer, S.N.; Wazer, D.E.; Band, V. E7 protein of human papilloma virus-16 induces degradation of retinoblastoma protein through the ubiquitin-proteasome pathway. Cancer Res. 1996, 56, 4620–4624. [Google Scholar]
- Scheffner, M.; Werness, B.A.; Huibregtse, J.M.; Levine, A.J.; Howley, P.M. The E6 oncoprotein encoded by human papillomavirus types 16 and 18 promotes the degradation of p53. Cell 1990, 63, 1129–1136. [Google Scholar] [CrossRef]
- Antinore, M.J.; Birrer, M.J.; Patel, D.; Nader, L.; McCance, D.J. The human papillomavirus type 16 E7 gene product interacts with and trans-activates the AP1 family of transcription factors. EMBO J. 1996, 15, 1950–1960. [Google Scholar] [CrossRef]
- Lui, W.O.; Pourmand, N.; Patterson, B.K.; Fire, A. Patterns of known and novel small RNAs in human cervical cancer. Cancer Res. 2007, 67, 6031–6043. [Google Scholar] [CrossRef] [Green Version]
- Martinez, I.; Gardiner, A.S.; Board, K.F.; Monzon, F.A.; Edwards, R.P.; Khan, S.A. Human papillomavirus type 16 reduces the expression of microRNA-218 in cervical carcinoma cells. Oncogene 2008, 27, 2575–2582. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tang, S.; Le, S.Y.; Lu, R.; Rader, J.S.; Meyers, C.; Zheng, Z.M. Aberrant expression of oncogenic and tumor-suppressive microRNAs in cervical cancer is required for cancer cell growth. PLoS ONE 2008, 3, e2557. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Yao, D.-S.; Chen, J.-Y.; Ding, N. Aberrant Expression of miR-20a and miR-203 in Cervical Cancer. Asian Pac. J. Cancer Prev. 2013, 14, 2289–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Schwarz, J.K.; Lewis, J.S.; Huettner, P.C.; Rader, J.S.; Deasy, J.O.; Grigsby, P.W.; Wang, X. A microRNA expression signature for cervical cancer prognosis. Cancer Res. 2010, 70, 1441–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.N.; Wang, L.F.; Jia, Y.F.; Hao, Y.Q.; Zhang, L.; Wang, H. Upregulation of microRNA-224 is associated with aggressive progression and poor prognosis in human cervical cancer. Diagn. Pathol. 2013, 8, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Meyers, C.; Guo, M.; Zheng, Z.M. Upregulation of p18Ink4c expression by oncogenic HPV E6 via p53-miR-34a pathway. Int. J. Cancer 2011, 129, 1362–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, R.; Wang, F.; Shi, L.Y.; Liu, M.; Chen, S.; Wan, H.Y.; Li, Y.X.; Li, X.; Gao, S.Y.; Sun, B.C.; et al. Plexin-B1 is a target of miR-214 in cervical cancer and promotes the growth and invasion of HeLa cells. Int. J. Biochem. Cell Biol. 2011, 43, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Lin, J.X.; Yu, Y.H.; Zhang, M.Y.; Wang, H.Y.; Zheng, M. Downregulation of six microRNAs is associated with advanced stage, lymph node metastasis and poor prognosis in small cell carcinoma of the cervix. PLoS ONE 2012, 7, e33762. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yao, D.; Li, Y.; Chen, H.; He, C.; Ding, N.; Lu, Y.; Ou, T.; Zhao, S.; Li, L.; et al. Serum microRNA expression levels can predict lymph node metastasis in patients with early-stage cervical squamous cell carcinoma. Int. J. Mol. Med. 2013, 32, 557–567. [Google Scholar] [CrossRef] [Green Version]
- White, R.A.; Neiman, J.M.; Reddi, A.; Han, G.; Birlea, S.; Mitra, D.; Dionne, L.; Fernandez, P.; Murao, K.; Bian, L.; et al. Epithelial stem cell mutations that promote squamous cell carcinoma metastasis. J. Clin. Investig. 2013, 123, 4390–4404. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Kinoshita, T.; Nohata, N.; Yoshino, H.; Itesako, T.; Fujimura, L.; Mitsuhashi, A.; Usui, H.; Enokida, H.; Nakagawa, M.; et al. Tumor-suppressive microRNA-29a inhibits cancer cell migration and invasion via targeting HSP47 in cervical squamous cell carcinoma. Int. J. Oncol. 2013, 43, 1855–1863. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Dua, P.; Agarwal, S.M. A Comprehensive Review of Dysregulated miRNAs Involved in Cervical Cancer. Curr. Genom. 2014, 15, 310–323. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Wu, Y.-L.; Wang, Y.-X.; Zhu, F.-F. Characterization of the MicroRNA Expression Profile of Cervical Squamous Cell Carcinoma Metastases. Asian Pac. J. Cancer Prev. 2014, 15, 1675–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Wong, H.K.; Gu, W.; Yu, M.Y.; To, K.F.; Wang, C.C.; Wong, Y.F.; Cheung, T.H.; Chung, T.K.H.; Choy, K.W. MicroRNA-182 plays an onco-miRNA role in cervical cancer. Gynecol. Oncol. 2013, 129, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hu, Y.; Ye, F.; Li, Y.; Lv, W.; Xie, X. Reduced miR-34a expression in normal cervical tissues and cervical lesions with high-risk human papillomavirus infection. Int. J. Gynecol Cancer 2010, 20, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Gocze, K.; Gombos, K.; Juhasz, K.; Kovacs, K.; Kajtar, B.; Benczik, M.; Gocze, P.; Patczai, B.; Arany, I.; Ember, I. Unique microRNA expression profiles in cervical cancer. Anticancer Res. 2013, 33, 2561–2567. [Google Scholar]
- Wang, G.; Shen, N.; Cheng, L.; Lin, J.; Li, K. Downregulation of miR-22 acts as an unfavorable prognostic biomarker in osteosarcoma. Tumor Biol. 2015, 36, 7891–7895. [Google Scholar] [CrossRef]
- Li, J.; Liang, S.; Yu, H.; Zhang, J.; Ma, D.; Lu, X. An inhibitory effect of miR-22 on cell migration and invasion in ovarian cancer. Gynecol. Oncol. 2010, 119, 543–548. [Google Scholar] [CrossRef]
- Wan, W.N.; Zhang, Y.Q.; Wang, X.M.; Liu, Y.J.; Zhang, Y.X.; Que, Y.H.; Zhao, W.J.; Li, P. Down-regulated miR-22 as predictive biomarkers for prognosis of epithelial ovarian cancer. Diagn. Pathol. 2014, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; He, J.; Zhang, Y. MicroRNA-22 expression in hepatocellular carcinoma and its correlation with ezrin protein. J. Int. Med. Res. 2013, 41, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Xia, S.; Tian, H.; Liu, Z.; Zhou, T. Clinical significance of miR-22 expression in patients with colorectal cancer. Med. Oncol. 2012, 29, 3108–3112. [Google Scholar] [CrossRef]
- Yang, C.; Ning, S.; Li, Z.; Qin, X.; Xu, W. miR-22 is down-regulated in esophageal squamous cell carcinoma and inhibits cell migration and invasion. Cancer Cell Int. 2014, 14, 138. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.M.; Liao, C.G.; Zhang, Y.; Xu, J.; Li, Y.; Huang, W.; Zhang, Y.; Bian, H.; Chen, Z.N. A regulatory loop involving miR-22, Sp1, and c-Myc modulates CD147 expression in breast cancer invasion and metastasis. Cancer Res. 2014, 74, 3764–3778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Liu, X.; Su, B.; Zhang, Z.; Zeng, X.; Lei, Y.; Shan, J.; Wu, Y.; Tang, H.; Su, Q. microRNA-22 acts as a metastasis suppressor by targeting metadherin in gastric cancer. Mol. Med. Rep. 2015, 11, 454–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, K.T.; Liu, Z.; Feng, J.; Zhao, W.; Hao, T.; Ding, W.Y.; Chu, J.P.; Gao, L.J. MiR-22-3p regulates cell proliferation and inhibits cell apoptosis through targeting the eIF4EBP3 gene in human cervical squamous carcinoma cells. Int. J. Med. Sci. 2018, 15, 142–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lin, C. Exosomes miR-22-3p derived from mesenchymal stem cells suppress colorectal cancer cell proliferation and invasion by regulating RAP2B and PI3K/AKT pathway. J. Oncol. 2021, 2021, 3874478. [Google Scholar] [CrossRef]
- Lajer, C.B.; Garnaes, E.; Friis-Hansen, L.; Norrild, B.; Therkildsen, M.H.; Glud, M.; Rossing, M.; Lajer, H.; Svane, D.; Skotte, L.; et al. The role of miRNAs in human papilloma virus (HPV)-associated cancers: Bridging between HPV-related head and neck cancer and cervical cancer. Br. J. Cancer 2012, 106, 1526–1534. [Google Scholar] [CrossRef] [PubMed]
- Higgins, G.D.; Davy, M.; Roder, D.; Uzelin, D.M.; Phillips, G.E.; Burrell, C.J. Increased age and mortality associated with cervical carcinomas negative for human papillomavirus RNA. Lancet 1991, 338, 910–913. [Google Scholar] [CrossRef]
- Markou, A.; Sourvinou, I.; Vorkas, P.A.; Yousef, G.M.; Lianidou, E. Clinical evaluation of microRNA expression profiling in non small cell lung cancer. Lung Cancer 2013, 81, 388–396. [Google Scholar] [CrossRef]
- Sugihara, H.; Ishimoto, T.; Watanabe, M.; Sawayama, H.; Iwatsuki, M.; Baba, Y.; Komohara, Y.; Takeya, M.; Baba, H. Identification of miR-30e* regulation of Bmi1 expression mediated by tumor-associated macrophages in gastrointestinal cancer. PLoS ONE 2013, 8, e81839. [Google Scholar] [CrossRef]
- Zhang, S.; Li, G.; Liu, C.; Lu, S.; Jing, Q.; Chen, X.; Zheng, H.; Ma, H.; Zhang, D.; Ren, S.; et al. miR-30e-5p represses angiogenesis and metastasis by directly targeting AEG-1 in squamous cell carcinoma of the head and neck. Cancer Sci. 2020, 111, 356–368. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| A. Upregulated miRNAs | B. Downregulated miRNAs | ||||
|---|---|---|---|---|---|
| miRNAs | Fold Change | p-Value | miRNAs | Fold Change | p-Value |
| hsa-miR-134 | 2.019 | 0.023 | hsa-miR-210 | −7.229 | <0.001 |
| hsa-miR-1185-2-3p | 2.066 | 0.017 | hsa-miR-142-3p | −5.740 | 0.002 |
| hsa-miR-4532 | 2.093 | 0.027 | hsa-miR-1260a | −5.345 | 0.004 |
| hsa-miR-652-5p | 2.142 | 0.024 | hsa-miR-429 | −5.062 | 0.025 |
| hsa-miR-155-5p | 2.311 | 0.002 | hsa-miR-15a-5p | −4.607 | 0.004 |
| hsa-miR-6717-5p | 2.478 | 0.011 | hsa-miR-19a-3p | −4.577 | 0.005 |
| hsa-miR-6131 | 2.583 | 0.006 | hsa-miR-141-3p | −3.743 | 0.011 |
| hsa-miR-1185-1-3p | 2.804 | 0.003 | hsa-miR-30e-5p | −3.593 | 0.015 |
| hsa-miR-6126 | 2.833 | 0.003 | hsa-miR-16-5p | −3.470 | 0.043 |
| hsa-miR-4734 | 2.997 | <0.001 | hsa-miR-200b-3p | −3.142 | 0.002 |
| hsa-miR-3198 | 3.085 | 0.005 | hsa-miR-103a-3p | −3.044 | 0.004 |
| hsa-miR-4713-3p | 3.125 | 0.011 | hsa-miR-107 | −2.998 | 0.005 |
| hsa-miR-5581-5p | 3.189 | 0.013 | hsa-miR-15b-5p | −2.899 | 0.015 |
| hsa-miR-874 | 3.220 | 0.004 | hsa-miR-22-3p | −2.618 | 0.003 |
| hsa-miR-6132 | 3.267 | 0.027 | hsa-miR-106b-5p | −2.426 | 0.008 |
| hsa-miR-1233-1-5p | 4.516 | <0.001 | hsa-miR-34a-5p | −2.290 | 0.003 |
| hsa-miR-4485 | 4.679 | 0.032 | hsa-miR-342-3p | −2.226 | <0.001 |
| hsa-miR-4695-5p | 5.950 | 0.017 | hsa-miR-93-5p | −2.123 | 0.020 |
| hsa-miR-4497 | 6.501 | 0.009 | hsa-miR-25-3p | −2.088 | 0.017 |
| hsa-miR-4430 | 6.522 | 0.042 | |||
| hsa-miR-4653-3p | 16.451 | 0.022 | |||
| miRNA | Cervical SqC (n = 65) | p-Value | Cervical SqC without Metastasis (n = 29) | Cervical SqC with Metastasis (n = 27) | Metastatic SqC from Cervix (n = 9) | p-Value |
|---|---|---|---|---|---|---|
| miR-22-3p | 1.079 ± 1.191 | 0.228 | 1.609 ± 1.415 | 0.819 ± 0.881 | 0.266 ± 0.181 | <0.001 * |
| miR-30e-5p | 0.857 ± 0.842 | 0.002 * | 0.891 ± 0.672 | 0.651 ± 0.600 | 1.370 ± 1.577 | 0.054 |
| miR-429 | 0.696 ± 0.839 | <0.001* | 0.634 ± 0.752 | 0.693 ± 0.780 | 0.900 ± 1.270 | 0.744 |
| miR-134 | 1.264 ± 1.319 | 0.429 | 1.359 ± 1.651 | 1.277 ± 0.902 | 0.930 ± 0.782 | 0.407 |
| miR-420 § | miR-22-3p § | miR-30e-5p § | miR-134 § | |||||
|---|---|---|---|---|---|---|---|---|
| Down-Regulation | p-Value | Down-Regulation | p-Value | Down-Regulation | p-Value | Up-Regulation | p-Value | |
| Age | ||||||||
| <60 | 9/49 (18.4%) | 0.583 | 13/49 (26.5%) | 0.666 | 13/49 (26.5%) | 0.182 | 36/49 (73.5%) | 0.182 |
| ≥60 | 0/7 (0.0%) | 1/7 (14.3%) | 0/7 (0.0%) | 3/7 (42.9%) | ||||
| Clinical stage | ||||||||
| I | 4/27 (14.8%) | 1.000 | 5/27 (18.5%) | 0.280 | 3/27 (11.1%) | 0.038 * | 20/27 (74.1%) | 0.487 |
| II, III and IV | 5/29 (17.2%) | 9/29 (31.0%) | 10/29 (34.5%) | 19/29 (65.5%) | ||||
| Nodal metastasis | ||||||||
| absent | 4/31 (12.9%) | 0.493 | 4/31 (12.9%) | 0.020 * | 4/31 (12.9%) | 0.042 * | 24/31 (77.4%) | 0.159 |
| present | 5/25 (20.0%) | 10/25 (40.0%) | 9/25 (36.0%) | 15/25 (60.0%) | ||||
| Lymphovascular invasion | ||||||||
| absent | 1/14 (7.1%) | 0.424 | 1/14 (7.1%) | 0.151 | 0/14 (0%) | 0.025 * | 1/14 (7.1%) | 0.043 * |
| present | 8/42 (19.0%) | 13/42 (31.0%) | 13/42 (31.0%) | 16/42 (38.1%) | ||||
| Distant metastasis | ||||||||
| absent | 9/50 (18.0%) | 0.575 | 13/50 (26.0%) | 1.000 | 12/50 (24.0%) | 1.000 | 36/50 (72.0%) | 0.354 |
| present | 0/6 (0.0%) | 1/6 (16.7%) | 1/6 (16.7%) | 3/6 (50.0%) | ||||
| Recurrence | ||||||||
| absent | 6/42 (14.3%) | 0.676 | 9/42 (21.4%) | 0.304 | 8/42 (19.0%) | 0.274 | 9/42 (21.4%) | 0.019 * |
| present | 3/14 (21.4%) | 5/14 (35.7%) | 5/14 (35.7%) | 8/14 (57.1%) | ||||
| HPV16 Positive | p-Value | |
| miR-22-3p up-regulation § | 11/12 (91.7%) | 0.021 * |
| no up-regulation | 24/44 (54.5%) | |
| HPV16 or 18 Positive | p-Value | |
| miR-22-3p up-regulation § | 12/12 (100.0%) | 0.005 * |
| no up-regulation | 25/44 (56.8%) |
| Case (n = 56) | Death | Overall Survival (Mon) | Hazard Ratio | p-Value | ||
|---|---|---|---|---|---|---|
| Age | <60 | 49 | 8 | 39.2 ± 26.1 | 34.14 | 0.026 * |
| ≥60 | 7 | 2 | 35.7 ± 18.0 | |||
| Clinical stage | I | 27 | 3 | 46.6 ± 30.7 | 0.16 | 0.235 |
| II/III/IV | 29 | 7 | 1.6 ± 15.9 | |||
| Nodal metastasis | absent | 31 | 1 | 39.7 ± 25.9 | 0.05 | 0.219 |
| present | 25 | 9 | 37.5 ± 24.7 | |||
| Lymphovascular invasion | absent | 14 | 0 | 38.9 ± 27.3 | 7.11 × 104 | 0.94 |
| present | 42 | 10 | 38.7 ± 24.8 | |||
| Distant metastasis | absent | 50 | 6 | 39.8 ± 26.0 | 123.76 | 0.025 * |
| present | 6 | 4 | 30.4 ± 15.1 | |||
| Recurrence | absent | 42 | 0 | 42.7 ± 27.1 | 1.73 × 107 | 0.865 |
| present | 14 | 10 | 26.8 ± 12.3 | |||
| miR-30e-5p § | up-regulation | 43 | 6 | 42.0 ± 26.7 | 289.66 | 0.040 * |
| down-regulation | 13 | 4 | 28.1 ± 15.7 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, A.-Y.; Jeong, J.-Y.; Park, H.; Hwang, S.; Kim, G.; Kang, H.; Heo, J.-H.; Lee, H.J.; Kim, T.-H.; An, H.J. miR-22-3p and miR-30e-5p Are Associated with Prognosis in Cervical Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 5623. https://doi.org/10.3390/ijms23105623
Kwon A-Y, Jeong J-Y, Park H, Hwang S, Kim G, Kang H, Heo J-H, Lee HJ, Kim T-H, An HJ. miR-22-3p and miR-30e-5p Are Associated with Prognosis in Cervical Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2022; 23(10):5623. https://doi.org/10.3390/ijms23105623
Chicago/Turabian StyleKwon, Ah-Young, Ju-Yeon Jeong, Hyun Park, Sohyun Hwang, Gwangil Kim, Haeyoun Kang, Jin-Hyung Heo, Hye Jin Lee, Tae-Heon Kim, and Hee Jung An. 2022. "miR-22-3p and miR-30e-5p Are Associated with Prognosis in Cervical Squamous Cell Carcinoma" International Journal of Molecular Sciences 23, no. 10: 5623. https://doi.org/10.3390/ijms23105623