
MITF in Normal Melanocytes, Cutaneous and Uveal Melanoma: A Delicate Balance
by
 , , ,
, , ,
Maria Chiara Gelmi
1 ,
,
Laurien E. Houtzagers
1,
Thomas Strub
2,3,
Imène Krossa
2,3 and
Martine J. Jager
1,* 1
Department of Ophthalmology, Leiden University Medical Center, P.O. Box 9600, 2300 RC Leiden, The Netherlands
2
Université Côte d’Azur, 06103 Nice, France
3
Inserm, Biology and Pathologies of Melanocytes, Team1, Equipe Labellisée Ligue 2020, Centre Méditerranéen de Médecine Moléculaire, 06204 Nice, France
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(11), 6001; https://doi.org/10.3390/ijms23116001
Submission received: 17 February 2022
/
Revised: 23 May 2022
/
Accepted: 24 May 2022
/
Published: 26 May 2022
(This article belongs to the Collection Feature Papers in Molecular Pathology, Diagnostics, and Therapeutics)
Abstract
:Microphthalmia-associated transcription factor (MITF) is an important regulator of melanogenesis and melanocyte development. Although it has been studied extensively in cutaneous melanoma, the role of MITF in uveal melanoma (UM) has not been explored in much detail. We review the literature about the role of MITF in normal melanocytes, in cutaneous melanoma, and in UM. In normal melanocytes, MITF regulates melanocyte development, melanin synthesis, and melanocyte survival. The expression profile and the behaviour of MITF-expressing cells suggest that MITF promotes local proliferation and inhibits invasion, inflammation, and epithelial-to-mesenchymal (EMT) transition. Loss of MITF expression leads to increased invasion and inflammation and is more prevalent in malignant cells. Cutaneous melanoma cells switch between MITF-high and MITF-low states in different phases of tumour development. In UM, MITF loss is associated with loss of BAP1 protein expression, which is a marker of poor prognosis. These data indicate a dual role for MITF in benign and malignant melanocytic cells.
1. Introduction
Melanocytes, which originate from the neural crest, are located in many areas of the human body, such as the skin and eye. Their most obvious physiological function is to produce melanin, which gives the skin, hair, and eye their characteristic colours. However, melanocytes have been demonstrated to interact closely with other cell types in the skin and to be involved in immune responses, to act as stress sensors and to have neuroendocrine properties [1,2,3].
The environment around melanocytes in the skin and in the eye is distinct: skin melanocytes are surrounded by keratinocytes, whereas the neighbours of ocular melanocytes can be fibroblasts, endothelial cells or epithelial cells, depending on the tissue (choroid, iris, ciliary body, or conjunctiva). In the eye, there is another type of pigmented cell, called a retinal pigmented epithelial cell, which is derived from the optic neuroepithelium. Their neighbours are photoreceptor cells.
Transformation of melanocytes leads to melanoma. Cutaneous malignant melanoma represents only 5% of skin cancers but is responsible for 75% of deaths by a cutaneous cancer. Its incidence is 30 to 380 cases per million per year and depends on the geographical location and genetic background [4]. Cutaneous malignant melanoma is epidemiologically linked to exposure to ultraviolet (UV) radiation of the solar light [5,6]. However, the relationship between UVR and cutaneous melanoma is not yet fully understood, given the absence of a UV DNA damage signature in genes relevant to cutaneous melanoma such as BRAF, NRAS, or CDKN2A (cyclin dependent kinase inhibitor 2A) [7]. UM (uveal melanoma) is the most common primary intraocular malignancy in adults. Although UM is relatively rare compared to cutaneous melanomas, with an incidence of 1 to 9 cases per million per year [8], depending on the geographical location, it contributes to a large proportion of melanoma death rates. Approximately 95% of ocular melanomas occur in the uvea (of which 90% are in the choroid, 6% in the ciliary body, and 4% in the iris), and 5% in the conjunctiva [8,9,10,11]. Driver mutations in choroidal melanomas are mutations in the heterotrimeric G-protein alpha subunits GNAQ and GNA11, while conjunctival melanomas show more resemblance to skin melanoma with oncogenic mutations in BRAF and NRAS.
Fair skin, blond/red hair, light iris colour and nevi are risk factors for both types of melanomas [12]. Risk factors for cutaneous melanoma also include freckles, intense sun exposure and strong skin reactions to sun as well as genetic mutations in a panel of genes such as CDKN2A, MC1R (melanocortin 1 receptor), MITF (microphthalmia-associated transcription factor) and BAP1 (BRCA1 associated protein 1) [13]. Risk factors for UM include germline mutations in BAP1 [14,15] and single nucleotide polymorphisms (SNP) in pigmentation genes such as HERC2/OCA2 and IRF4 (interferon regulatory factor 4) [16].
As a light iris colour (blue/grey/green) is especially seen in populations in North-Western Europe and worldwide in people of Northern European ancestry, areas inhabited by these populations have the highest number of UM cases [17].
MITF plays a critical role in melanocyte development as well as their function, survival, and proliferation and is considered the master gene of melanocyte homeostasis and a critical factor in melanoma biology (as reviewed by Cheli et al. [18] and by Goding and Arnheiter [19]). MITF is a basic helix-loop-helix-zipper (bHLHZip) transcription factor encoded on chromosome 3p12.3-14.1. It encodes multiple isoforms that show ubiquitous or tissue-specific expression. MITF-M is the main isoform of the melanocyte lineage, while MITF-D and MITF-H are detected in adult retinal pigment epithelium (RPE). MITF-H is also expressed in the heart tissue whereas MITF-A is ubiquitously expressed [19]. MITF has been extensively studied in skin melanocytes and cutaneous melanoma over the past decades.
We reviewed the literature regarding the role and regulation of MITF expression and its function in normal cutaneous melanocytes and compared its expression and function between cutaneous melanoma and UM cells.
2. Normal Regulation
2.1. Role of MITF in Skin Melanocyte Development
During embryonic development, pluripotent neural crest cells give rise to non-pigmented melanoblasts. The melanoblasts then migrate from both the dorsolateral and the ventral pathways to the basal layer of the epidermis and to the hair follicles. In the epidermis, melanoblasts differentiate into mature melanocytes and start producing melanin, which is transferred to surrounding keratinocytes to ensure skin pigmentation. Melanocytes are not present at the dermal–epidermal junction of mouse skin except on the ear, tail, nose, and foot pad. The hair follicle contains several different types of melanocytes with different properties in different locations [20,21]. Melanogenically-active melanocytes can be found in three locations (close to the dermal papilla, in the infundibulum and in the sebaceous glands), while melanogenically-inactive melanocytes are located in the stem cell compartment of the hair bulge, in the outer root sheath, and in the peripheral–proximal hair bulb.
MITF plays a key role in melanocyte development. In humans, mutations in MITF have been identified in patients with Waardenburg syndrome (WS) type 2A and Tietz syndrome, leading to pigmentation defects such as patches of light skin and iris heterochromia and deafness [22,23]. More recently, mutations in MITF have been identified in patients with Coloboma who also show osteopetrosis, microphthalmia, macrocephaly, albinism, and deafness [24]. In mice, heterozygous MITF mutants usually do not show a visible phenotype but some alleles can lead to alterations in pigmentation of the skin including white spots on the belly, head, tail, and a reduction in iris pigmentation. In contrast, absence of MITF leads to a complete lack of neural crest-derived pigment cells with not only a lack of pigmentation but also with ear and eye developmental abnormalities [25]. Defects in mast cells and osteoclasts have been observed in homozygotes carrying certain mutant alleles [25,26].
The essential role of MITF-M in melanocyte development is demonstrated in the black-eyed white Mitfmi-bw mice [27], which harbour the insertion of an L1 retrotransposable element in intron 3, leading to complete repression of MITF-M mRNA expression. These mice exhibit a completely white coat colour due to deficiency of melanocytes in their skin and hair bulbs, with deafness, and black eyes: they lack choroidal melanocytes but their RPE cells are normally developed and pigmented [27,28].
MITF-M is also required for melanoblast survival [29,30]. This can be mediated through induction of the anti-apoptotic target genes BCL-2 (B-cell-lymphoma 2) [31], and of DICER, which was shown to be crucial to melanocyte survival through its function in the processing of miRNAs [32].
During embryonic development, melanocytes undergo epithelial-to-mesenchymal transition (EMT) in order to delaminate from the neural tube [33]. EMT consists of a shift from an epithelial-like, less-motile phenotype to a mesenchymal-like, more-motile phenotype. EMT is regulated by several transcription factors (EMT-TFs) among which are SNAI1 (snail family transcriptional repressor 1), SNAI2 (snail family transcriptional repressor 2), zinc-finger E box-binding homeobox 1 and 2 (ZEB1 and ZEB2), and twist-related protein 1 (TWIST1) [34]. Since melanocytes are not epithelial in nature, but originate from the neural crest, we may refer to EMT in this cell type as pseudo-EMT. A further interesting function of MITF is its interaction with pseudo-EMT, which is involved in pathological processes such as cancer. A mutation in SNAI2 has been observed in two patients with Waardenburg syndrome type 2, and MITF has been shown to induce the transcription of SNAI2, the promoter of which has a potential MITF binding site [35].
2.2. Role of MITF in Melanocyte Function (Melanocytes and Melanin)
MITF has an important role in melanocyte function, since it is involved in several processes, such as melanin synthesis and pigmentation, and melanocyte structure, survival, proliferation, and differentiation.
Skin colour is related to the type of melanin, the size, and the number of melanosomes, and how the melanocytes are dispersed in the keratinocytes. Melanogenesis is under complex regulatory control by multiple agents, interacting via pathways activated by receptor-dependent and -independent mechanisms, in hormonal, auto-, para-, or intracrine fashion or by UV radiation [36]. The most pro-pigmentation pathway involves the binding of α-MSH (α-melanocyte stimulating hormone), produced by the keratinocytes, on its MC1R, which is expressed by melanocytes. Mechanistically, α-MSH binding to MC1R triggers elevation of cAMP and activation of the PKA/CREB (cAMP response element)-signalling pathway that stimulates transcription of a variety of downstream targets, including MITF [37,38].
Melanin is synthetized in specialized lysosomal-related organelles called melanosomes. They can produce two types of melanin: eumelanin is a brownish-black insoluble polymer associated with protection against UV-mediated DNA damage, while pheomelanin is a yellowish-red soluble polymer associated with the induction of genotoxic stress [39]. The main enzymes that drive melanogenesis are TYR (tyrosinase), TYRP-1 (tyrosine-related protein-1) and DCT (dopachrome tautomerase, also known as tyrosine-related protein-2, TYRP-2). The expression of these melanogenic enzymes is regulated by MITF through a direct interaction with an E-box/M-box in their promoters [18,40,41,42]. The activation of the DCT promoter by MITF happens in cooperation with CREB, SOX10 (SRY-box transcription factor 10), or LEF-1 (lymphoid enhancer binding factor 1) [43,44,45]. PAX3 (paired box 3) has been shown to have a dual role: on the one hand it antagonises the MITF-mediated stimulation of DCT, while on the other hand, it enhances MITF expression [46]. Specific MC1R variants have been associated with red hair and a high pheomelanin content [47], but not with iris pigmentation [48]. The cAMP-dependent signalling pathway also operates through the expression of PKCβ (protein kinase C beta) via MITF, which has been reported to phosphorylate and activate TYR [49].
The production of melanin starts with the oxidation of L-tyrosine to L-DOPA and then to DOPAquinone (DQ) by TYR; this is a rate-limiting step (Figure 1). DQ can follow two distinct pathways, leading to either the production of eumelanin or pheomelanin: it can undergo cyclisation and give rise to cyclodopa, followed by a redox reaction that produces dopachrome and dihydroxyphenylalanine (DOPA). While DOPA is converted back to DQ by TYR, dopachrome either undergoes decarboxylation to dihydroxyindole (DHI) or is converted into dihydroxindole-2-carboxylic acid (DHICA) by DCT. Both DHI and DHICA are subsequently oxidised to eumelanin. In the presence of cysteine or glutathione, DQ is converted to 5-S-cysteinyldopa or glutathionyldopa, which are then oxidised to benzothiazine and finally to pheomelanin [50].
In the skin, each melanocyte is in contact with on average 36 keratinocytes, thereby forming the epidermal melanin unit. Mature melanin-containing melanosomes are thought to be transported to the melanocyte’s dendrite tips in an actin- and microtubule-dependent process that is regulated by Rab27a, melanophilin, and myosin Va [51,52,53,54] and subsequently to be transferred to keratinocytes to ensure homogeneous skin pigmentation. These processes also implicate MITF, which regulates the transcription and expression of genes involved in melanosome functioning (GPR143/OA1, SILV/PMEL/gp100, MLANA/MART-1 [18,55,56]), and transport (Rab27). The mechanisms governing melanosome transfer and positioning are still debated and poorly understood [57,58].
It is worth noting that L-tyrosine and L-dihydroxyphenylalanine (L-DOPA), in addition to serving as substrates and intermediates of melanogenesis, are thought to regulate other melanocyte functions through action on specific receptors or through non-receptor-mediated mechanisms [36,59].
At the same time, there is increasing evidence that melanocytes may have other functions in the skin. Indeed, being capable of secreting a wide range of signalling molecules (cytokines, POMC peptides including α-MSH, catecholamines, and NO), melanocytes could impact on a range of skin cells to maintain epidermal homeostasis [2,60].
As mentioned previously, pigmentation and melanin production are not the only functions of MITF in melanocytes. MITF is also involved in melanocyte survival, proliferation, and cell cycle. It is a positive regulator of the anti-apoptotic gene BCL2 (BCL2 apoptosis regulator) both in normal melanocytes and in human cutaneous melanoma cells [31], and of BIRC7 (baculoviral IAP repeat containing 7) in melanoma cells [61]. Levy et al. reported that MITF regulates DICER, which is a regulator of miRNA processing and is necessary for melanocyte development and survival [32]. Moreover, Du et al. reported that MITF controls the expression of CDK2 (cyclin dependent kinase 2) both in normal melanocytes and in melanoma cells, and that this interaction is relevant for the growth of melanoma cells [62]. Among the targets of MITF we find factors involved in cell cycle-regulation as well: MITF can increase expression of p21 (CDKN1A) and p16 INK4a (CDKN2A), thus causing cell cycle exit and impairing cell growth in melanocytes and in fibroblasts treated with MITF [63,64]. Moreover, both p21 and p16INK4a induce fibroblasts to acquire features typical of melanocyte differentiation.
A summary of the main MITF targets is presented in Figure 2.
2.3. Regulation of MITF Expression in Normal Skin Melanocytes
The MITF-M promoter is upregulated by the transcription factors PAX3 [65] and SOX10. Mutations in PAX3 are responsible for WS types 1 and 3 and mutations in SOX10 for WS type 4 [66]. Moreover, Wnt signalling is required for the expansion and for determining the fate of neural crest derivatives, including melanocytes during early development [67,68,69]. Wnt-3a expression is detected at embryonic day 7.5 [70], which precedes that of MITF expression in neural crest cells (9.5–10.5 days) [30]. Wnt3 regulates β-catenin, which translocates into the nucleus, where it then interacts with LEF/TCF (lymphoid enhancer-binding factor/T cell factor) and stimulates MITF transcription [70,71]. A study on mouse hair follicles and skin showed that ZEB2 knockout greatly impaired melanocyte differentiation and decreased the expression of MITF and other pigment-related genes in mouse melanocyte, suggesting that MITF might be a direct target of EMT factors [72]. As already mentioned, MC1R, through activation of the PKA/CREB module, is an important regulator of MITF transcription in the skin. SOX10 enhances MITF expression and it cooperates with CREB and PAX3, which is particularly active in early melanocyte development and is inhibited by TGFβ (transforming growth factor β) [66,73,74,75]. These transcription factors are thus regulators of MITF, while they themselves are regulated by MITF as well. Knockout of the receptor tyrosine kinase KIT in melanocytes causes a severe white spotting phenotype [76]. Mechanistically, SCF (stem cell factor) binds to c-Kit and stimulates the MAPK (mitogen activated protein kinase) and PI3K (phosphatidyl inositol-3-kinase) pathways. Activation of ERK (extracellular signal regulated kinase) and subsequent phosphorylation of MITF on Ser73 increases MITF activity [77,78,79] (Figure 3). Ser73 phosphorylation is a priming site for GSK3 (glycogen synthase kinase 3) phosphorylation on Ser69 which prevents MITF nuclear export, while phosphorylation on Ser409 may prime for phosphorylation of S405, S401, and S397 by GSK3 [80,81]. Thus, BRAF/MAPK and PI3K/GSK3 signalling converges to control MITF nuclear export and activity. Finally, miR-218 suppressed melanogenesis in human pigmented skin, through MITF repression [82]. In conclusion, in cutaneous melanocytes, MITF expression is regulated through several signalling pathways, both at the mRNA and protein level, in which epigenetic regulation also plays a role.
3. MITF in Cutaneous Melanoma
Cutaneous malignant melanomas are heterogeneous in nature, comprising several cell subpopulations with distinct transcriptomic signatures and behaviours [83]. Melanomas carrying different genetic alterations have different clinical features and different relation with sun exposure. A study of the TCGA database divided melanomas into four genomic subtypes: BRAF mutated (more common in younger patients, associated with intermittent sun exposure), NRAS mutated (more common in older patients, associated with chronic UV damage), NFI mutated (present in acral and mucosal melanomas, associated with chronic UV damage), and triple wild type (lacking a UV signature, including uveal melanoma) [84]. Other mutations and genetic alterations are known to be present in cutaneous melanoma, such as CDKN2A mutation, PTEN loss, and amplification of MITF, EGFR, CCDN1, cMET, and cKIT [85,86,87]. Epigenetic modifications such as changes in the DNA methylation status, chromatin remodelling and non-coding RNA regulation may also influence the behaviour of melanoma cells and the response to therapy [88]. Two main cell populations have been identified: a fast-replicating population with low invasive potential (with high MITF expression) and a slowly proliferating population with high invasive potential (with low MITF expression). MITF-low cutaneous melanoma cells display a higher expression of stem cell markers (OCT4 and NANOG) and are able to produce larger tumours when injected into nude mice [89]. However, both MITF-low and MITF-high cells can give rise to tumours, which then contain both types of cells [89,90]. This finding served as evidence that these two populations of cells are not genetically determined but that the proliferative or invasive potential is likely to be the result of a phenotype switch [89,91]. While it is not yet fully clear what determines the switch between the two phenotypes, factors such as hypoxia and nutrient starvation have been implicated and MITF is thought to play a key role [89,92].
3.1. Cell Proliferation
MITF has been shown to enhance or to inhibit proliferation. On one hand, MITF may behave as a melanocyte specific oncogene. MITF is expressed in about 80% of human melanomas (since it is not frequently expressed in desmoplastic melanomas) [93,94]; it is amplified in 10% of primary and 20% of metastatic cutaneous melanomas and its expression correlates with decreased 5-year overall patient survival [95]. Furthermore, a rare germline variant in the MITF gene (E318K variant) has been linked to a high total nevus count and an increased risk of cutaneous melanoma [96,97]. Another illustration of MITF’s role in proliferation is its ability to control the expression of the cell cycle regulators. MITF is known to regulate transcription and expression of the cyclin dependent kinases CDK2 (cyclin dependent kinase 2) and CDK4 (cyclin dependent kinase 4) [95,98], of TBX2 (T-box transcription factor 2, a transcription factor of the T-box family), that in turn blocks senescence through repression of p21 and p19 [99,100,101,102], and of several genes involved in mitosis [103]. Moreover, MITF exerts a positive control over cell cycle progression through degradation of the growth inhibitor p27 on the one hand, while at the same time it inhibits invasiveness. Both of these functions are carried out through the DIAPH1 (diaphanous related formin 1) gene [104]. MITF also drives expression of a large subset of genes involved in lysosome biogenesis and functioning [81], which triggers increased activity of the lysosome-bound mTORC1 (mTOR complex 1) and global protein synthesis [105]. Likewise, MITF controls expression of the metabolic factor PGC1a (PPARG coactivator 1 alpha) [106,107]. Enhanced levels of protein synthesis and metabolic activities could allow cancer cells to cope with the metabolic demand related to the high proliferative rate associated with MITF. Consequently, MITF knockdown in human cutaneous melanoma cell lines promotes a growth arrest through induction of a senescence-like phenotype [108]. On the other hand, MITF has been reported to exert an antiproliferative effect in cutaneous melanoma cells, essentially via p21 regulation [63].
3.2. Cell Survival
MITF has been shown to promote cell survival in melanoma cells through several mechanisms. MITF binds E-boxes on the promoters of anti-apoptotic target genes BCL2, BCL2A1 (BCL2 related protein A1) [109] and BIRC7 (baculoviral IAP repeat containing 7) [31,61] and participates in transactivation of the receptor tyrosine kinase MET, thereby increasing the anti-apoptotic effect of the MET ligand HGF (hepatocyte growth factor) [110]. Under oxidative stress conditions, MITF activates APE1/Ref1 (apurinic-apyrimidinic endonuclease1⁄redox factor-1), which is a protein involved in DNA repair and in redox regulation [111]. HIF1α is one of the targets of APE1/Ref1 [112]. HIF1α is also a direct target of MITF. Aligned with this, enhanced HIF1α expression impaired staurosporin-induced cell death in cutaneous melanoma cells [113].
3.3. Epithelial-Mesenchymal Transition and Motile Ability
EMT is a complex process in which epithelial cells acquire the characteristics of invasive mesenchymal cells. Melanoma tumour progression and metastasis formation involves a pseudo-EMT process (given the non-epithelial nature of melanoma cells) in which MITF is also involved. Normal cutaneous melanocytes have a high expression of SNAI2 and ZEB2 and a low expression of ZEB1 and TWIST1, while malignant cutaneous melanomas have low SNAI2 and ZEB2 and high ZEB1 and TWIST1 [114]. Survival analysis in cutaneous melanoma patients showed that high TWIST1 and ZEB1 expression was associated with a shorter metastasis-free survival. Moreover, in vitro BRAF activation caused a switch from a ZEB2high/SNAI2high/ZEB1low/TWIST1low state (similar to normal melanocytes) to a ZEB2low/SNAI2low/ZEB1high/TWIST1high state and MEK inhibitors reversed this switch [114]. Gene expression profiling revealed that cell lines with high ZEB1 and TWIST1 had a de-differentiated gene signature characterised by an upregulation of invasion-associated and TGFβ-regulated genes and downregulation of MITF and its target genes [114]. The role of ZEB2 was confirmed in another study, in which human primary cutaneous melanoma samples with high nuclear ZEB2 staining were associated with a better prognosis than tumours with low nuclear ZEB2 staining [72]. ZEB2 knockdown in mouse melanoma cell lines led to a decrease in MITF and its target genes and an increase in ZEB1, thereby leading to a more invasive phenotype. A subsequent study by the same group confirmed that ZEB2 is associated with a proliferative gene signature that includes MITF, while ZEB1 is associated with an invasive gene signature [115]. A schematic representation of the role of MITF in pseudo-EMT is presented in Figure 4.
In conclusion, MITF’s role in melanoma cells is important and complex. The rheostat model proposed by the group of Goding provides explanation to the apparent paradox that MITF controls or represses the proliferation or the motile ability of melanoma cells [104]. MITF expression levels are important but by far not the only criterion in physiology and pathology of melanocytes and melanoma cells. MITF activity also depends on its post-translational modifications (phosphorylation, SUMOylation, ubiquitination) and co-factors (such as p300, BRG1, β-catenin). Hence, cells with low MITF levels are poorly proliferative (likely due to increased p27 expression), dedifferentiated, more mesenchymal and motile [104], whereas cells with high MITF activity are differentiated and growth-arrested in part through p21 induction [63]. Supporting this model, positive MITF staining in the primary tumour was associated with a better survival and lower rates of lymph node involvement in cutaneous melanoma patients [116,117].
3.4. Regulation of MITF in Cutaneous Melanoma
Multiple factors and stimuli have been shown to control MITF-M expression in cutaneous melanoma cells. As previously described for skin melanocytes, CREB, PAX3, SOX10, and the Wnt/β-catenin module are also well known upstream regulators of MITF in cutaneous melanoma cells [19]. In contrast, ATF4 (activating transcription factor 4), and JUN, which both integrate stress signals, repress MITF expression and trigger cutaneous melanoma cell dedifferentiation [118,119,120]. Another gene that is able to repress MITF is BRN2, which mediates melanoma cell invasion [121]. This gene is negatively regulated at the post-transcriptional level by miR-211, which is in turn upregulated by MITF. miR-211 is not the only miRNA involved in the regulation of the invasiveness of cutaneous melanoma cells [122]. Data regarding miR-182 have shown conflicting results, with some authors reporting it to be upregulated in advanced melanoma and other authors stating that it is downregulated in cutaneous melanoma samples [123]. Changes in the tumour microenvironment, such as hypoxia or nutrient starvation, may also cause a decrease in MITF expression and greater invasiveness [92,118,124]. Cutaneous melanoma cells exposed to hypoxia have a higher HIF1α expression and lower MITF expression and give rise to larger tumours and more frequent metastases [92]. The downregulation of MITF caused by hypoxia is dependent on HIF1α, which through the transcription factor Bhlhb2 represses the MITF promoter [92,125]. Melanogenesis has been shown to generate an immunosuppressive and mutagenic environment and alters the glycolytic metabolism through HIF1α induction [126,127]. Likewise, glutamine starvation reduces MITF level through ATF4 induction [118]. Additional cues such as TNFα (tumour necrosis factor α) and TGFβ are likely factors that induce the phenotype switch in cutaneous melanoma cells. TNFα can also activate ATF4 and JUN, resulting in dedifferentiated cutaneous melanoma cells [118,119,120]. TGFβ antagonizes MITF function, represses pigmentation and stimulates the motile ability of cutaneous melanoma cells [128,129].
3.5. Clinical Relevance
Both immune checkpoint inhibitors (ICI) and molecularly targeted therapy with BRAF and MEK inhibitors (BRAF/MEKi) are standard options for patients with BRAFV600-mutated, unresectable, or metastatic melanoma. Options in BRAF wild-type melanoma are limited to ICIs. Despite the progress brought by these treatments, about 50% of the patients reach a therapeutic dead end due to primary or secondary resistance. As mentioned above, cutaneous melanomas are heterogeneous tumours comprised of cells with distinct transcriptomic signatures driving specific behaviours. The two most studied types of melanoma cells are those with a proliferative or invasive phenotype. Due to their high intrinsic plasticity, cutaneous melanoma cells can switch back and forth between these two phenotypes. This plasticity is thought to create intratumour heterogeneity which plays a key role in treatment failure and relapse. Thus, it is of paramount importance to better understand the features of these cell subpopulations to improve treatment efficiency. MITF is thought to be the very determinant of melanoma cell plasticity [89,130]. Genomic amplification of the MITF target, BCL2A1, has been implicated in resistance to BRAF inhibitors by Haq et al. [109]. Moreover, macrophage-derived TNFα, through increased MITF expression, provides resistance to MAPK pathway inhibitors [131]. Consequently, inhibition of MITF, by introduction of a dominant-negative MITF mutant in melanoma cells with MITF amplification, or by blocking the TNFα signalling with IκB kinase inhibitors, increased susceptibility of cutaneous melanoma cells to chemotherapeutic agents or MAPK pathway inhibitors, respectively [95,131]. These observations suggest that MITF inhibition may represent an option to increase therapy efficacy. However, low MITF expression in cutaneous melanoma cells has also been linked to phenotype switching and drug resistance. Indeed, MITF knockdown in melanoma cells leads to a senescent-like phenotype that is associated with NFkB (nuclear factor kappa-light-chain-enhancer of activated B cell) activation and production of an inflammatory secretome (CCL2, IL6, IL1). This secretome favours a more mesenchymal phenotype, thereby favouring melanoma progression and metastatic dissemination [132,133]. Aligned with these findings, Konieczkowski et al. showed that BRAF-mutated melanoma cells with intrinsic resistance to MAPK inhibitors display a low MITF and high NFkB expression [134]. Likewise, NFkB induction by TNFα led to a decrease in MITF expression and conferred resistance to MAPK inhibitors [134]. MITF low cells are also associated with an increase in the expression of stem cell genes and reprogramming to a more invasive cell state. Interestingly, MITF activity has been recently shown to be regulated by a direct interaction with RAF proteins in melanoma cells [135]. By triggering a partial relocation of MITF in the cytoplasm, this interaction might reduce nuclear concentrations of MITF, thereby impacting phenotype switching and therapy efficacy. Melanisation level can also affect the therapy and the clinical outcome of advanced pigmented melanomas [136,137,138].
As highlighted in a review by Ballotti et al., in addition to resistance to targeted therapy, downregulation of MITF may be also responsible for resistance to immunotherapy [139] and subsequent melanoma progression [139]. Inflammatory signals produced by low MITF melanoma cells or by the microenvironment can induce de-differentiation and the consequent loss of melanocyte-specific surface antigens [139,140]. Collectively, these observations are in agreement with Müller et al. reporting the existence of two types of resistant cell lines: one with high or normal levels of MITF and one with low MITF level [141]. Nevertheless, they showed that the cell lines with low MITF levels are more resistant to a wider panel and higher concentrations of MAPK pathway inhibitors than the ones with high MITF [141].
4. MITF and the Eye
In the eye, pigment producing cells can be found in different locations. Melanocytes in the choroid, iris, ciliary body, and conjunctiva are derived from the dorsal neural crest and follow the same path as epidermal melanocytes for development, migrating along the dorsolateral pathway from the neural crest to the uvea or conjunctiva. Studies in mice showed that melanoblasts in the eye reach the choroid at an early embryological stage and increase in density in the early postnatal phase, acquiring a perivascular distribution. The RPE, which is composed of a single layer of cells containing pigment granules, is derived from the optic vesicle [142].
The regulation of eye colour is far less understood than skin colour. Both eye melanocytes and RPE cells contain melanin-filled melanosomes. More important than the melanin ratio is the density of melanosomes in the stromal melanocytes, with density increasing from blue to brown eyes [143,144]. A study by Wakamatsu et al. showed that iridal and choroidal melanocytes from eyes with a dark iris had significantly higher total melanin content and a higher eumelanin/pheomelanin ratio than melanocytes from eyes with a light iris [145]. Iris tissue from light eyes also showed a slightly higher pheomelanin content compared to dark eyes, but it did not reach statistical significance. There is no compelling evidence suggesting that the pigment-containing structures are secreted and/or transferred to adjacent cells or that RPE cells produce pheomelanin.
Genetic defects in expression of some melanogenic enzymes (TYR, TYRP1) or other melanin synthesis regulators (P protein, SLC45A2,) result in oculocutaneous albinism (OCA1-OCA4) and visual function impairment, demonstrating the importance of melanin for proper ocular function [146,147]. Single-nucleotide polymorphisms (SNPs) in other genes including HERC2/OCA2, LYST (lysosomal trafficking regulator), IRF4, and SLC24A4 (solute carrier family 24 member 4) have also been implicated in determining eye colour [148,149,150,151]. Simcoe et al. published the results of a very large genome-wide association study involving two European cohorts and two smaller Asian cohorts and reported 50 novel genetic loci related to eye colour (alongside the previously-known loci). Interestingly, MITF and DCT were among the genes containing significantly-associated SNPs [152].
4.1. Expression of MITF in Ocular Tissues
Several transcription factors are involved in the specification of intraocular tissues and MITF plays a crucial role. Initially, MITF-M has been shown to be expressed in melanocytes but not in RPE cell lines [153] while studies have shown important roles of MITF-H and MITF-D in mouse RPE specification and differentiation [39,154]. MITF-H and MITF-D are controlled by the retina-specific transcription factor CHX10 [154]. Absence of H- and D-MITF in MITF red-eyed white (MITFmi-rw) mutant mice, which may present with low and variable levels of pigmentation in the peripheral iris, exhibit severe RPE developmental abnormalities [154]. It is worth noting that MITF-H has the ability to compensate for reduced RPE pigmentation induced by MITF-D loss [155]. Other MITF mutations can cause RPE and retinal dysfunction, degeneration and thus worsen vision [156]. Isoforms MITF-A and MITF-J are more ubiquitous, as they are expressed in melanocytes and RPE cells as well as in retinal cells, but they do not seem to have a significant impact on eye development [154,157] (Table 1). More recently, Esumi and coworkers uncovered that differentiated human adult RPE cells expressed MITF-M [158]. Nevertheless, the black-eyed white Mitfmi-bw mutant mice lack MITF-M expression and melanocytes in the skin and choroid, but retain RPE pigmentation [27,159], indicating that MITF-M is not essential for RPE development or differentiation. Therefore, MITF-M might be functional in human adult RPE, although its functions in RPE cells and whether they are distinct from those of the other MITF isoforms remain to be determined.
It is worth noting that mice with the MITFmi-bw mutation developed a thinner choroid, containing only one vascular layer, as opposed to the normal two layers, indicating that MITF has been observed to have a role in the development of choroidal vessels [160].
4.2. Uveal Nevi
In the choroid, melanocytes that acquire mutations in either GNAQ, GNA11, or CYSLTR2 can give rise to nevi [161,162]. Usually, cutaneous nevi do not carry these mutations, being characterised by BRAF and NRAS mutations [163,164]. However, somatic mutations in GNAQ, GNA11, or CYSLTR2 can be found in dermal blue nevi [165,166]. These mutations are not per se able to induce melanocyte transformation and melanoma development.
The risk of developing a UM is increased in people with an oculodermal pigment aberration, known as nevus of Ota (1 in 400 compared to 1 in 1300 in the general population) [167]. The nevus of Ota may harbour mutations in GNAQ but a study on one patient also revealed the presence of a BAP1 mutation in the primary tumour and TP53 (tumour protein 53) mutations in a local recurrence [168].
4.3. MITF in Ocular Melanoma
It seems eye melanomas are derived exclusively from neural crest melanocytes but never from the neuroepithelium-derived RPE or the posterior layer of the iris. The molecular mechanisms underlying eye melanocyte transformation, especially in the choroid and ciliary body, are not known. In both conjunctival and iris melanoma, molecular findings support the occurrence of UV-induced DNA damage, indicating a link between solar radiation and development of these tumours [169,170]. Clinically and histologically, conjunctival melanomas, which harbour mutations in BRAF and NRAS, show greater similarity to cutaneous melanoma than does UM. Like choroidal nevi, a UM usually carries either a GNAQ or a GNA11 mutation [171,172]. When a second mutation occurs, it usually involves one of three genes: BAP1, EIF1AX, or SF3B1 [8,173].
BAP1 is a high-risk locus in UM and germline BAP1 mutations increase the risk of developing UM and other tumours such as melanocytic skin tumours, mesothelioma, and renal cell carcinoma (known as the BAP1-predisposition syndrome) [8,174]. Alongside BAP1, some other low-risk loci have been linked to the risk of developing UM: specific SNPs in the pigmentation genes HERC2 and OCA2 are associated with a lower risk, and IRF4 is associated with a higher risk, and deleterious germline mutations of MBD4, which give rise to UM with a hypermutator phenotype [8,16].
Only a few studies analysed the presence and function of MITF in UM and, as for cutaneous melanoma, its role seems to be complex: some studies classify MITF as pro-oncogenic, and others mention its expression as a feature of low-risk tumours. Mouriaux et al. showed that 65% of UM were positive for nuclear MITF staining. They found a negative association between MITF staining and tumour pigmentation but a positive correlation between MITF staining and proliferative activity [175]. These observations indicate that, like in cutaneous melanoma, MITF might be linked to the proliferative ability of UM cells. Hippo-YAP/TAZ also emerged as an important signalling pathway downstream of GNAQ/11 that controls UM cell proliferation. Downstream of this module lies PAX3, which controls MITF expression [176]. As such, YAP inhibition suppressed the growth of UM [177,178]. In contrast, MITF has been shown to increase p16 expression in UM, where CDKN2A mutations have rarely been described, supporting the idea that MITF can induce cell cycle arrest and behave as a tumour suppressor gene [64]. Similar to the situation in cutaneous melanoma [122,123], miRNAs and epigenetic mechanisms are involved in MITF regulation in UM cells: miR-137 through downregulation of MITF, MET, and CDK6 (cyclin dependent kinase 6) and miR-182 through inhibition of MITF, MET, BCL2, and cyclin D2, causing G1 cell cycle arrest, thereby reducing the number of metabolically-active UM cells [179,180]. In addition, miR-182 (which is activated by p53) causes apoptosis of UM cells [180]. Increase in miR-137 by the DNA hypomethylating agent 5-aza-2′-deoxycytidine or the histone deacetylase inhibitor trichostatin A (TSA) also represents a therapeutic opportunity to impair UM cell proliferation through MITF inhibition [179]. However, as in cutaneous melanoma, MITF inhibition might be associated with a stem cell-like phenotype. Matatall et al. showed that BAP1 knockdown was associated with an increase in expression of stem cell markers (NANOG), and loss of pigmentation markers (MITF, TYR, and DCT) and with the capacity of UM cells to form anchorage-independent colonies. These data suggest that BAP1 loss induced a dedifferentiated and a stem-like phenotype in UM cells, although this remains to be fully demonstrated [181]. Like cutaneous melanoma, MITF inhibition may activate the NFkB pathway. Stimulation of the NFkB pathway has been reported to favour tumour aggressiveness in UM cells [182]. A comparison of gene expression in normal choroidal melanocytes and primary UM showed downregulation of MITF target genes involved in pigmentation (TYRP1, DCT) and the detection of stemness markers including OCT4 in primary UM cells [183]. MITF high and MITF low melanoma cells can coexist in a UM and contribute to intra-tumoural heterogeneity, as has recently been described [184,185,186].
4.4. MITF as a Therapeutic Target
Currently, there are no available direct MITF inhibitors. Therefore, it is important to continue to understand how MITF is regulated and functions in order to identify potential upstream regulators and/or downstream effectors that may be targeted. In line with that, MITF has been reported as a critical downstream target of the HAT (histone acetyl transferase) p300 promoting human cutaneous melanoma growth [187]. A small molecule p300/CBP inhibitor dramatically reduced the number of human cutaneous melanoma cells in vitro. However, inhibitors of HDAC (histone deacetylase) have also been shown to decrease MITF expression and to inhibit tumour growth in a human cutaneous melanoma xenograft model [188]. Aligned with that, the HDAC trichostatin A (TSA) increases miR-137, which in turn reduces MITF expression in uveal melanoma cells [179]. A recent study showed that both the HDAC inhibitor ACY-1215 and a small molecule inhibitor of the MITF pathway (ML329) were able to reduce the proliferation of a metastatic UM cell line in vitro and caused regression of tumours derived from this cell line in zebrafish, but did not influence tumour dissemination [189]. Moreover, UM cells treated with ACY-1215 showed a concomitant decrease in the protein expression of MITF and several of its targets [189]. This apparent contradiction, i.e., inhibition of HAT and HDAC leading to reduction of MITF activity, remains to be elucidated. The DNA hypomethylating agent 5-aza-2′-deoxycytidine, which is FDA approved for some cancer treatments, has also been described to increase miR-137 in uveal melanoma cells and thus could be therapeutically exploited [179].
However, data connecting MITF expression with a highly proliferative, yet more differentiated and less invasive phenotype raise the question of whether downregulating MITF, which may trigger a phenotype switch to a slowly-proliferating and highly-invasive melanoma cell state, will actually be beneficial in the long term.
5. Conclusions
MITF has several functions in normal melanocytes: it promotes melanocyte development, differentiation, proliferation, and survival. Its role in the biology of eye melanocytes has been less studied but mouse models show that it plays a key role in visual function. Although not yet fully defined, MITF has roles in melanoma development and progression. Its activity depends on its level of expression, cofactors, and post-translational modifications as proposed by Goding [19].
Given that cutaneous and uveal melanoma are governed by mutations in different genes and are found in different environments, the role of MITF might differ between skin and eye melanomas, although we see many similarities.
Author Contributions
M.C.G., L.E.H., T.S., I.K. and M.J.J. collaboratively wrote this review. All authors have read and agreed to the published version of the manuscript.
Funding
M.C. Gelmi was funded by the Bontius Foundation, Oogfonds, the Sam Fund, the LUF, the P.A. Jager-van Gelder Fund, the Blinden-Penning foundation, and ASROO (Associazione Scientifica Retinoblastoma ed Oncologia Oculare). The sponsor or funding organisation had no role in the design or conduct of this research.
Acknowledgments
T.S. acknowledges the financial support provided by La Fondation pour la Recherche Médicale (FRM ARF201809006989) and the Fondation de France (00120250/WB-2021-33281). I.K. is a fellowship from Région Provence-Alpes-Côte d’Azur “Emplois jeunes doctorants”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Plonka, P.M.; Passeron, T.; Brenner, M.; Tobin, D.J.; Shibahara, S.; Thomas, A.; Slominski, A.; Kadekaro, A.L.; Hershkovitz, D.; Peters, E.; et al. What are melanocytes really doing all day long...? Exp. Dermatol. 2009, 18, 799–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.; Paus, R.; Schadendorf, D. Melanocytes as “sensory” and regulatory cells in the epidermis. J. Theor. Biol. 1993, 164, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A. Neuroendocrine activity of the melanocyte. Exp. Dermatol. 2009, 18, 760–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saginala, K.; Barsouk, A.; Aluru, J.S.; Rawla, P.; Barsouk, A. Epidemiology of Melanoma. Med. Sci. 2021, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.F.; Hodis, E.; Heffernan, T.P.; Deribe, Y.L.; Lawrence, M.S.; Protopopov, A.; Ivanova, E.; Watson, I.R.; Nickerson, E.; Ghosh, P.; et al. Melanoma genome sequencing reveals frequent PREX2 mutations. Nature 2012, 485, 502–506. [Google Scholar] [CrossRef]
- Krauthammer, M.; Kong, Y.; Ha, B.H.; Evans, P.; Bacchiocchi, A.; McCusker, J.P.; Cheng, E.; Davis, M.J.; Goh, G.; Choi, M.; et al. Exome sequencing identifies recurrent somatic RAC1 mutations in melanoma. Nat. Genet. 2012, 44, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Craig, S.; Earnshaw, C.H.; Virós, A. Ultraviolet light and melanoma. J. Pathol. 2018, 244, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Jager, M.J.; Shields, C.L.; Cebulla, C.M.; Abdel-Rahman, M.H.; Grossniklaus, H.E.; Stern, M.-H.; Carvajal, R.D.; Belfort, R.N.; Jia, R.; Shields, J.A.; et al. Uveal melanoma. Nat. Rev. Dis. Prim. 2020, 6, 24. [Google Scholar] [CrossRef]
- Shields, C.L.; Kaliki, S.; Furuta, M.; Mashayekhi, A.; Shields, J.A. Clinical spectrum and prognosis of uveal melanoma based on age at presentation in 8033 cases. Retina 2012, 32, 1363–1372. [Google Scholar] [CrossRef]
- McLaughlin, C.C.; Wu, X.-C.; Jemal, A.; Martin, H.J.; Roche, L.M.; Chen, V.W. Incidence of noncutaneous melanomas in the U.S. Cancer 2005, 103, 1000–1007. [Google Scholar] [CrossRef]
- Isager, P.; Østerlind, A.; Engholm, G.; Heegaard, S.; Lindegaard, J.; Overgaard, J.; Storm, H.H. Uveal and conjunctival malignant melanoma in Denmark, 1943–1997: Incidence and validation study. Ophthalmic Epidemiol. 2005, 12, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Weis, E.; Shah, C.P.; Lajous, M.; Shields, J.A.; Shields, C.L. The Association Between Host Susceptibility Factors and Uveal Melanoma. Arch. Ophthalmol. 2006, 124, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paillerets, B.B.-D.; Lesueur, F.; Bertolotto, C. A germline oncogenic MITF mutation and tumor susceptibility. Eur. J. Cell Biol. 2014, 93, 71–75. [Google Scholar] [CrossRef]
- Walpole, S.; Pritchard, A.L.; Cebulla, C.M.; Pilarski, R.; Stautberg, M.; Davidorf, F.H.; De La Fouchardière, A.; Cabaret, O.; Golmard, L.; Stoppa-Lyonnet, D.; et al. Comprehensive Study of the Clinical Phenotype of Germline BAP1 Variant-Carrying Families Worldwide. J. Natl. Cancer Inst. 2018, 110, 1328–1341. [Google Scholar] [CrossRef] [PubMed]
- Derrien, A.-C.; Rodrigues, M.; Eeckhoutte, A.; Dayot, S.; Houy, A.; Mobuchon, L.; Gardrat, S.; Lequin, D.; Ballet, S.; Pierron, G.; et al. Germline MBD4 Mutations and Predisposition to Uveal Melanoma. J. Natl. Cancer Inst. 2020, 113, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, R.; Vogelsang, M.; Ucisik-Akkaya, E.; Rai, K.; Pilarski, R.; Martinez, C.N.; Rendleman, J.; Kazlow, E.; Nagdimov, K.; Osman, I.; et al. Genetic markers of pigmentation are novel risk loci for uveal melanoma. Sci. Rep. 2016, 6, 31191. [Google Scholar] [CrossRef]
- Houtzagers, L.E.; Wierenga, A.P.A.; Ruys, A.A.M.; Luyten, G.P.M.; Jager, M.J. Iris Colour and the Risk of Developing Uveal Melanoma. Int. J. Mol. Sci. 2020, 21, 7172. [Google Scholar] [CrossRef]
- Cheli, Y.; Ohanna, M.; Ballotti, R.; Bertolotto, C. Fifteen-year quest for microphthalmia-associated transcription factor target genes. Pigment Cell Melanoma Res. 2010, 23, 27–40. [Google Scholar] [CrossRef]
- Goding, C.R.; Arnheiter, H. MITF—The first 25 years. Genes Dev. 2019, 33, 983–1007. [Google Scholar] [CrossRef] [Green Version]
- Tobin, D.J. The cell biology of human hair follicle pigmentation. Pigment Cell Melanoma Res. 2010, 24, 75–88. [Google Scholar] [CrossRef]
- Casalou, C.; Moreiras, H.; Mayatra, J.M.; Fabre, A.; Tobin, D.J. Loss of ‘Epidermal Melanin Unit’ Integrity in Human Skin during Melanoma-Genesis. Front. Oncol. 2022, 12, 878336. [Google Scholar] [CrossRef] [PubMed]
- Tassabehji, M.; Newton, V.E.; Read, A.P. Waardenburg syndrome type 2 caused by mutations in the human microphthalmia (MITF) gene. Nat. Genet. 1994, 8, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D.; Kelley, P.M.; Kenyon, J.B.; Hoover, D. Tietz syndrome (hypopigmentation/deafness) caused by mutation of MITF. J. Med. Genet. 2000, 37, 446–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, A.; Zand, D.J.; Hufnagel, R.B.; Sharma, R.; Sergeev, Y.V.; Legare, J.M.; Rice, G.M.; Schwoerer, J.A.S.; Rius, M.; Tetri, L.; et al. Biallelic Mutations in MITF Cause Coloboma, Osteopetrosis, Microphthalmia, Macrocephaly, Albinism, and Deafness. Am. J. Hum. Genet. 2016, 99, 1388–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgkinson, C.A.; Moore, K.J.; Nakayama, A.; Steingrímsson, E.; Copeland, N.G.; Jenkins, N.A.; Arnheiter, H. Mutations at the mouse microphthalmia locus are associated with defects in a gene encoding a novel basic-helix-loop-helix-zipper protein. Cell 1993, 74, 395–404. [Google Scholar] [CrossRef]
- Silvers, W.K. The Coat Colors of Mice—A Model for Mammalian Gene Action and Interaction; Springer: New York, NY, USA, 1979. [Google Scholar]
- Yajima, I.; Sato, S.; Kimura, T.; Yasumoto, K.; Shibahara, S.; Goding, C.; Yamamoto, H. An L1 element intronic insertion in the black-eyed white (Mitf[mi-bw]) gene: The loss of a single Mitf isoform responsible for the pigmentary defect and inner ear deafness. Hum. Mol. Genet. 1999, 8, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, H.; Hozawa, K.; Oshima, T.; Takeuchi, T.; Takasaka, T. Dysgenesis of melanocytes and cochlear dysfunction in mutant microphthalmia (mi) mice. Hear. Res. 1994, 80, 10–20. [Google Scholar] [CrossRef]
- Opdecamp, K.; Nakayama, A.; Nguyen, M.T.; Hodgkinson, C.A.; Pavan, W.J.; Arnheiter, H. Melanocyte development in vivo and in neural crest cell cultures: Crucial dependence on the Mitf basic-helix-loop-helix-zipper transcription factor. Development 1997, 124, 2377–2386. [Google Scholar] [CrossRef]
- Nakayama, A.; Nguyen, M.-T.T.; Chen, C.C.; Opdecamp, K.; Hodgkinson, C.A.; Arnheiter, H. Mutations in microphthalmia, the mouse homolog of the human deafness gene MITF, affect neuroepithelial and neural crest-derived melanocytes differently. Mech. Dev. 1998, 70, 155–166. [Google Scholar] [CrossRef]
- McGill, G.G.; Horstmann, M.A.; Widlund, H.; Du, J.; Motyckova, G.; Nishimura, E.K.; Lin, Y.-L.; Ramaswamy, S.; Avery, W.; Ding, H.-F.; et al. Bcl2 Regulation by the Melanocyte Master Regulator Mitf Modulates Lineage Survival and Melanoma Cell Viability. Cell 2002, 109, 707–718. [Google Scholar] [CrossRef] [Green Version]
- Levy, C.; Khaled, M.; Robinson, K.C.; Veguilla, R.A.; Chen, P.-H.; Yokoyama, S.; Makino, E.; Lu, J.; Larue, L.; Beermann, F.; et al. Lineage-specific transcriptional regulation of DICER by MITF in melanocytes. Cell 2010, 141, 994–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mort, R.L.; Jackson, I.J.; Patton, E.E. The melanocyte lineage in development and disease. Development 2015, 142, 620–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto, M.A.; Huang, R.Y.-J.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Martín, M.; Rodríguez-García, A.; Pérez-Losada, J.; Sagrera, A.; Read, A.P.; Sánchez-García, I. SLUG (SNAI2) deletions in patients with Waardenburg disease. Hum. Mol. Genet. 2002, 11, 3231–3236. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Bertolotto, C.; Abbe, P.; Hemesath, T.J.; Bille, K.; Fisher, D.E.; Ortonne, J.-P.; Ballotti, R. Microphthalmia Gene Product as a Signal Transducer in cAMP-Induced Differentiation of Melanocytes. J. Cell Biol. 1998, 142, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Cui, R.; Widlund, H.R.; Feige, E.; Lin, J.Y.; Wilensky, D.L.; Igras, V.E.; D’Orazio, J.; Fung, C.Y.; Schanbacher, C.F.; Granter, S.R.; et al. Central Role of p53 in the Suntan Response and Pathologic Hyperpigmentation. Cell 2007, 128, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Wakamatsu, K. Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: A comparative review. Pigment Cell Res. 2003, 16, 523–531. [Google Scholar] [CrossRef]
- Aoki, H.; Moro, O. Involvement of microphthalmia-associated transcription factor (MITF) in expression of human melanocortin-1 receptor (MC1R). Life Sci. 2002, 71, 2171–2179. [Google Scholar] [CrossRef]
- Galibert, M.D.; Carreira, S.; Goding, C.R. The Usf-1 transcription factor is a novel target for the stress-responsive p38 kinase and mediates UV-induced Tyrosinase expression. EMBO J. 2001, 20, 5022–5031. [Google Scholar] [CrossRef] [Green Version]
- de la Serna, I.L.; Ohkawa, Y.; Higashi, C.; Dutta, C.; Osias, J.; Kommajosyula, N.; Tachibana, T.; Imbalzano, A.N. The microphthalmia-associated transcription factor requires SWI/SNF enzymes to activate melanocyte-specific genes. J. Biol. Chem. 2006, 281, 20233–20241. [Google Scholar] [CrossRef] [Green Version]
- Bertolotto, C.; Buscà, R.; Abbe, P.; Bille, K.; Aberdam, E.; Ortonne, J.-P.; Ballotti, R. Different cis-acting elements are involved in the regulation of TRP1 and TRP2 promoter activities by cyclic AMP: Pivotal role of M boxes (GTCATGTGCT) and of microphthalmia. Mol. Cell. Biol. 1998, 18, 694–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasumoto, K.; Takeda, K.; Saito, H.; Watanabe, K.; Takahashi, K.; Shibahara, S. Microphthalmia-associated transcription factor interacts with LEF-1, a mediator of Wnt signaling. EMBO J. 2002, 21, 2703–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, A.; Rehberg, S.; Wegner, M. Melanocyte-specific expression of dopachrome tautomerase is dependent on synergistic gene activation by the Sox10 and Mitf transcription factors. FEBS Lett. 2004, 556, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Lang, D.; Lu, M.M.; Huang, L.; Engleka, K.A.; Zhang, M.; Chu, E.Y.; Lipner, S.; Skoultchi, A.; Millar, S.E.; Epstein, J.A. Pax3 functions at a nodal point in melanocyte stem cell differentiation. Nature 2005, 433, 884–887. [Google Scholar] [CrossRef]
- Valverde, P.; Healy, E.; Jackson, I.J.; Rees, J.L.; Thody, A.J. Variants of the melanocyte–stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat. Genet. 1995, 11, 328–330. [Google Scholar] [CrossRef]
- Metzelaar-Blok, J.A.W.; Ter Huurne, J.A.C.; Hurks, H.M.H.; Keunen, J.E.; Jager, M.J.; Gruis, N.A. Characterization of melanocortin-1 receptor gene variants in uveal melanoma patients. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1951–1954. [Google Scholar]
- Park, H.-Y.; Wu, C.; Yonemoto, L.; Murphy-Smith, M.; Wu, H.; Stachur, C.M.; Gilchrest, B.A. MITF mediates cAMP-induced protein kinase C-β expression in human melanocytes. Biochem. J. 2006, 395, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Manickam, M.; Jung, S.-H. Recent development of signaling pathways inhibitors of melanogenesis. Cell. Signal 2017, 40, 99–115. [Google Scholar] [CrossRef]
- Chiaverini, C.; Beuret, L.; Flori, E.; Busca, R.; Abbe, P.; Bille, K.; Bahadoran, P.; Ortonne, J.-P.; Bertolotto, C.; Ballotti, R. Microphthalmia-associated Transcription Factor Regulates RAB27A Gene Expression and Controls Melanosome Transport. J. Biol. Chem. 2008, 283, 12635–12642. [Google Scholar] [CrossRef] [Green Version]
- Oberhofer, A.; Spieler, P.; Rosenfeld, Y.; Stepp, W.L.; Cleetus, A.; Hume, A.N.; Mueller-Planitz, F.; Ökten, Z. Myosin Va’s adaptor protein melanophilin enforces track selection on the microtubule and actin networks in vitro. Proc. Natl. Acad. Sci. USA 2017, 114, E4714–E4723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passeron, T.; Bahadoran, P.; Bertolotto, C.; Chiaverini, C.; Buscà, R.; Valony, G.; Bille, K.; Ortonne, J.P.; Ballotti, R. Cyclic AMP promotes a peripheral distribution of melanosomes and stimulates melanophilin/Slac2-a and actin association. FASEB J. 2004, 18, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, P.; Aberdam, E.; Mantoux, F.; Buscà, R.; Bille, K.; Yalman, N.; de Saint-Basile, G.; Casaroli-Marano, R.; Ortonne, J.P.; Ballotti, R. Rab27a: A key to melanosome transport in human melanocytes. J. Cell Biol. 2001, 152, 843–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Miller, A.J.; Widlund, H.R.; Horstmann, M.A.; Ramaswamy, S.; Fisher, D.E. MLANA/MART1 and SILV/PMEL17/GP100 are transcriptionally regulated by MITF in melanocytes and melanoma. Am. J. Pathol. 2003, 163, 333–343. [Google Scholar] [CrossRef]
- Vetrini, F.; Auricchio, A.; Du, J.; Angeletti, B.; Fisher, D.E.; Ballabio, A.; Marigo, V. The microphthalmia transcription factor (Mitf) controls expression of the ocular albinism type 1 gene: Link between melanin synthesis and melanosome biogenesis. Mol. Cell. Biol. 2004, 24, 6550–6559. [Google Scholar] [CrossRef] [Green Version]
- Moreiras, H.; Pereira, F.J.C.; Neto, M.V.; Lopes, L.; Festas, T.C.; Seabra, M.C.; Barral, D.C. The exocyst is required for melanin exocytosis from melanocytes and transfer to keratinocytes. Pigment Cell Melanoma Res. 2020, 33, 366–371. [Google Scholar] [CrossRef]
- Benito-Martínez, S.; Salavessa, L.; Raposo, G.; Marks, M.S.; Delevoye, C. Melanin Transfer and Fate within Keratinocytes in Human Skin Pigmentation. Integr. Comp. Biol. 2021, 61, 1546–1555. [Google Scholar] [CrossRef]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators of melanocyte functions. Pigment Cell Melanoma Res. 2012, 25, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Tsatmali, M.; Ancans, J.; Thody, A.J. Melanocyte Function and Its Control by Melanocortin Peptides. J. Histochem. Cytochem. 2002, 50, 125–133. [Google Scholar] [CrossRef]
- Dynek, J.N.; Chan, S.M.; Liu, J.; Zha, J.; Fairbrother, W.J.; Vucic, D. Microphthalmia-Associated Transcription Factor Is a Critical Transcriptional Regulator of Melanoma Inhibitor of Apoptosis in Melanomas. Cancer Res. 2008, 68, 3124–3132. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Widlund, H.R.; Horstmann, M.A.; Ramaswamy, S.; Ross, K.; Huber, W.E.; Nishimura, E.K.; Golub, T.R.; Fisher, D.E. Critical role of CDK2 for melanoma growth linked to its melanocyte-specific transcriptional regulation by MITF. Cancer Cell 2004, 6, 565–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreira, S.; Goodall, J.; Aksan, I.; La Rocca, S.A.; Galibert, M.-D.; Denat, L.; Larue, L.; Goding, C.R. Mitf cooperates with Rb1 and activates p21Cip1 expression to regulate cell cycle progression. Nature 2005, 433, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Loercher, A.E.; Tank, E.M.; Delston, R.B.; Harbour, J.W. MITF links differentiation with cell cycle arrest in melanocytes by transcriptional activation of INK4A. J. Cell Biol. 2005, 168, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, A.; Takeda, K.; Ploplis, B.; Tachibana, M. Epistatic relationship between Waardenburg Syndrome genes MITF and PAX3. Nat. Genet. 1998, 18, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Bondurand, N.; Pingault, V.; Goerich, D.E.; Lemort, N.; Sock, E.; Le Caignec, C.; Wegner, M.; Goossens, M. Interaction among SOX10, PAX3 and MITF, three genes altered in Waardenburg syndrome. Hum. Mol. Genet. 2000, 9, 1907–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Jeannet, J.-P.; He, X.; Varmus, H.E.; Dawid, I.B. Regulation of dorsal fate in the neuraxis by Wnt-1 and Wnt-3a. Proc. Natl. Acad. Sci. USA 1997, 94, 13713–13718. [Google Scholar] [CrossRef] [Green Version]
- Ikeya, M.; Lee, S.M.K.; Johnson, J.E.; McMahon, A.P.; Takada, S. Wnt signalling required for expansion of neural crest and CNS progenitors. Nature 1997, 389, 966–970. [Google Scholar] [CrossRef]
- Dorsky, R.I.; Moon, R.T.; Raible, D.W. Control of neural crest cell fate by the Wnt signalling pathway. Nature 1998, 396, 370–373. [Google Scholar] [CrossRef]
- Takeda, K.; Yasumoto, K.-I.; Takada, R.; Takada, S.; Watanabe, K.-I.; Udono, T.; Saito, H.; Takahashi, K.; Shibahara, S. Induction of Melanocyte-specific Microphthalmia-associated Transcription Factor by Wnt-3a. J. Biol. Chem. 2000, 275, 14013–14016. [Google Scholar] [CrossRef] [Green Version]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [Green Version]
- Denecker, G.; Vandamme, N.; Akay, O.; Koludrovic, D.; Taminau, J.; Lemeire, K.; Gheldof, A.; De Craene, B.; Van Gele, M.; Brochez, L.; et al. Identification of a ZEB2-MITF-ZEB1 transcriptional network that controls melanogenesis and melanoma progression. Cell Death Differ. 2014, 21, 1250–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Li, Y.; Nishimura, E.K.; Xin, H.; Zhou, A.; Guo, Y.; Dong, L.; Denning, M.F.; Nickoloff, B.J.; Cui, R. Inhibition of PAX3 by TGF-β Modulates Melanocyte Viability. Mol. Cell 2008, 32, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.E.; Price, E.R.; Widlund, H.R.; Du, J.; Davis, I.J.; Wegner, M.; Fisher, D.E. A tissue-restricted cAMP transcriptional response: SOX10 modulates alpha-melanocyte-stimulating hormone-triggered expression of microphthalmia-associated transcription factor in melanocytes. J. Biol. Chem. 2003, 278, 45224–45230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubic, J.D.; Young, K.P.; Plummer, R.S.; Ludvik, A.E.; Lang, D. Pigmentation PAX-ways: The role of Pax3 in melanogenesis, melanocyte stem cell maintenance, and disease. Pigment Cell Melanoma Res. 2008, 21, 627–645. [Google Scholar] [CrossRef] [Green Version]
- Aoki, H.; Tomita, H.; Hara, A.; Kunisada, T. Conditional Deletion of Kit in Melanocytes: White Spotting Phenotype Is Cell Autonomous. J. Investig. Dermatol. 2015, 135, 1829–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemesath, T.J.; Price, E.R.; Takemoto, C.M.; Badalian, T.; Fisher, D.E. MAP kinase links the transcription factor Microphthalmia to c-Kit signalling in melanocytes. Nature 1998, 391, 298–301. [Google Scholar] [CrossRef]
- Wu, M.; Hemesath, T.J.; Takemoto, C.M.; Horstmann, M.A.; Wells, A.G.; Price, E.R.; Fisher, D.Z.; Fisher, D.E. c-Kit triggers dual phosphorylations, which couple activation and degradation of the essential melanocyte factor Mi. Genes Dev. 2000, 14, 301–312. [Google Scholar] [CrossRef]
- Phung, B.; Sun, J.; Schepsky, A.; Steingrimsson, E.; Rönnstrand, L. C-KIT signaling depends on microphthalmia-associated transcription factor for effects on cell proliferation. PLoS ONE 2011, 6, e24064. [Google Scholar] [CrossRef] [Green Version]
- Ngeow, K.C.; Friedrichsen, H.J.; Li, L.; Zeng, Z.; Andrews, S.; Volpon, L.; Brunsdon, H.; Berridge, G.; Picaud, S.; Fischer, R.; et al. BRAF/MAPK and GSK3 signaling converges to control MITF nuclear export. Proc. Natl. Acad. Sci. USA 2018, 115, E8668–E8677. [Google Scholar] [CrossRef] [Green Version]
- Ploper, D.; Taelman, V.F.; Robert, L.; Perez, B.S.; Titz, B.; Chen, H.-W.; Graeber, T.G.; von Euw, E.; Ribas, A.; De Robertis, E.M. MITF drives endolysosomal biogenesis and potentiates Wnt signaling in melanoma cells. Proc. Natl. Acad. Sci. USA 2015, 112, E420–E429. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Zhang, J.-F.; Wang, W.-M.; Cheung, F.W.-K.; Lu, Y.-F.; Ng, C.-F.; Kung, H.-F.; Liu, W.-K. MicroRNA-218 inhibits melanogenesis by directly suppressing microphthalmia-associated transcription factor expression. RNA Biol. 2014, 11, 732–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambow, F.; Rogiers, A.; Marin-Bejar, O.; Aibar, S.; Femel, J.; Dewaele, M.; Karras, P.; Brown, D.; Chang, Y.H.; Debiec-Rychter, M.; et al. Toward Minimal Residual Disease-Directed Therapy in Melanoma. Cell 2018, 174, 843–855.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Cancer Genome Atlas Network. Genomic Classification of Cutaneous Melanoma. Cell 2015, 161, 1681–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sini, M.C.; Doneddu, V.; Paliogiannis, P.; Casula, M.; Colombino, M.; Manca, A.; Botti, G.; Ascierto, P.A.; Lissia, A.; Cossu, A.; et al. Genetic alterations in main candidate genes during melanoma progression. Oncotarget 2018, 9, 8531–8541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, E.J.; Johnson, D.B.; Sosman, J.A.; Chandra, S. Melanoma: What do all the mutations mean? Cancer 2018, 124, 3490–3499. [Google Scholar] [CrossRef]
- Rabbie, R.; Ferguson, P.; Molina-Aguilar, C.; Adams, D.J.; Robles-Espinoza, C.D. Melanoma subtypes: Genomic profiles, prognostic molecular markers and therapeutic possibilities. J. Pathol. 2019, 247, 539–551. [Google Scholar] [CrossRef]
- Giunta, E.F.; Arrichiello, G.; Curvietto, M.; Pappalardo, A.; Bosso, D.; Rosanova, M.; Diana, A.; Giordano, P.; Petrillo, A.; Federico, P.; et al. Epigenetic Regulation in Melanoma: Facts and Hopes. Cells 2021, 10, 2048. [Google Scholar] [CrossRef]
- Cheli, Y.; Guiliano, S.; Botton, T.; Rocchi, S.; Hofman, V.; Hofman, P.; Bahadoran, P.; Bertolotto, C.; Ballotti, R. Mitf is the key molecular switch between mouse or human melanoma initiating cells and their differentiated progeny. Oncogene 2011, 30, 2307–2318. [Google Scholar] [CrossRef] [Green Version]
- Hoek, K.S.; Eichhoff, O.M.; Schlegel, N.C.; Döbbeling, U.; Kobert, N.; Schaerer, L.; Hemmi, S.; Dummer, R. In vivo switching of human melanoma cells between proliferative and invasive states. Cancer Res. 2008, 68, 650–656. [Google Scholar] [CrossRef] [Green Version]
- Pinner, S.; Jordan, P.; Sharrock, K.; Bazley, L.; Collinson, L.; Marais, R.; Bonvin, E.; Goding, C.; Sahai, E. Intravital imaging reveals transient changes in pigment production and Brn2 expression during metastatic melanoma dissemination. Cancer Res. 2009, 69, 7969–7977. [Google Scholar] [CrossRef] [Green Version]
- Cheli, Y.; Giuliano, S.; Fenouille, N.; Allegra, M.; Hofman, V.; Hofman, P.; Bahadoran, P.; Lacour, J.-P.; Tartare-Deckert, S.; Bertolotto, C.; et al. Hypoxia and MITF control metastatic behaviour in mouse and human melanoma cells. Oncogene 2012, 31, 2461–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, R.; Googe, P.B.; Weilbaecher, K.N.; Mihm, M.C., Jr.; Fisher, D.E. Microphthalmia Transcription Factor Expression in Cutaneous Benign, Malignant Melanocytic, and Nonmelanocytic Tumors. Am. J. Surg. Pathol. 2001, 25, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Granter, S.R.; Weilbaecher, K.N.; Quigley, C.; Fletcher, C.D.; Fisher, D.E. Microphthalmia transcription factor: Not a sensitive or specific marker for the diagnosis of desmoplastic melanoma and spindle cell (non-desmoplastic) melanoma. Am. J. Dermatopathol. 2001, 23, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Garraway, L.A.; Widlund, H.R.; Rubin, M.A.; Getz, G.; Berger, A.J.; Ramaswamy, S.; Beroukhim, R.; Milner, J.D.A.; Granter, S.R.; Du, J.; et al. Integrative genomic analyses identify MITF as a lineage survival oncogene amplified in malignant melanoma. Nature 2005, 436, 117–122. [Google Scholar] [CrossRef]
- Bertolotto, C.; Lesueur, F.; Giuliano, S.; Strub, T.; De Lichy, M.; Bille, K.; Dessen, P.; D’Hayer, B.; Mohamdi, H.; Remenieras, A.; et al. A SUMOylation-defective MITF germline mutation predisposes to melanoma and renal carcinoma. Nature 2011, 480, 94–98. [Google Scholar] [CrossRef]
- Ainger, S.A.; Jagirdar, K.; Lee, K.J.; Soyer, H.P.; Sturm, R.A. Skin Pigmentation Genetics for the Clinic. Dermatology 2017, 233, 1–15. [Google Scholar] [CrossRef]
- Wellbrock, C.; Arozarena, I. Microphthalmia-associated transcription factor in melanoma development and MAP-kinase pathway targeted therapy. Pigment. Cell Melanoma Res. 2015, 28, 390–406. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Teja, M.; Aladowicz, E.; Lanfrancone, L.; Goding, C.R. Tbx3 represses E-cadherin expression and enhances melanoma invasiveness. Cancer Res. 2008, 68, 7872–7881. [Google Scholar] [CrossRef] [Green Version]
- Carreira, S.; Liu, B.; Goding, C.R. The gene encoding the T-box factor Tbx2 is a target for the microphthalmia-associated transcription factor in melanocytes. J. Biol. Chem. 2000, 275, 21920–21927. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J.J.L.; Keblusek, P.; Robanus-Maandag, E.; Kristel, P.; Lingbeek, M.; Nederlof, P.M.; Van Welsem, T.; van de Vijver, M.J.; Koh, E.Y.; Daley, G.Q.; et al. Senescence bypass screen identifies TBX2, which represses Cdkn2a (p19ARF) and is amplified in a subset of human breast cancers. Nat. Genet. 2000, 26, 291–299. [Google Scholar] [CrossRef]
- Prince, S.; Carreira, S.; Vance, K.W.; Abrahams, A.; Goding, C.R. Tbx2 Directly Represses the Expression of the p21WAF1Cyclin-Dependent Kinase Inhibitor. Cancer Res. 2004, 64, 1669–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strub, T.; Giuliano, S.; Ye, T.; Bonet, C.; Keime, C.; Kobi, D.; Le Gras, S.; Cormont, M.; Ballotti, R.; Bertolotto, C.; et al. Essential role of microphthalmia transcription factor for DNA replication, mitosis and genomic stability in melanoma. Oncogene 2011, 30, 2319–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreira, S.; Goodall, J.; Denat, L.; Rodriguez, M.; Nuciforo, P.; Hoek, K.S.; Testori, A.; LaRue, L.; Goding, C.R. Mitf regulation of Dia1 controls melanoma proliferation and invasiveness. Genes Dev. 2006, 20, 3426–3439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoncu, R.; Bar-Peled, L.; Efeyan, A.; Wang, S.; Sancak, Y.; Sabatini, D.M. mTORC1 senses lysosomal amino acids through an inside-out mechanism that requires the vacuolar H(+)-ATPase. Science 2011, 334, 678–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haq, R.; Shoag, J.; Andreu-Perez, P.; Yokoyama, S.; Edelman, H.; Rowe, G.C.; Frederick, D.T.; Hurley, A.D.; Nellore, A.; Kung, A.L.; et al. Oncogenic BRAF regulates oxidative metabolism via PGC1α and MITF. Cancer Cell 2013, 23, 302–315. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, F.; Lim, J.-H.; Chim, H.; Bhalla, K.; Girnun, G.; Pierce, K.; Clish, C.B.; Granter, S.R.; Widlund, H.R.; Spiegelman, B.M.; et al. PGC1α expression defines a subset of human melanoma tumors with increased mitochondrial capacity and resistance to oxidative stress. Cancer Cell 2013, 23, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, S.; Cheli, Y.; Ohanna, M.; Bonet, C.; Beuret, L.; Bille, K.; Loubat, A.; Hofman, V.; Hofman, P.; Ponzio, G.; et al. Microphthalmia-Associated Transcription Factor Controls the DNA Damage Response and a Lineage-Specific Senescence Program in Melanomas. Cancer Res. 2010, 70, 3813–3822. [Google Scholar] [CrossRef] [Green Version]
- Haq, R.; Yokoyama, S.; Hawryluk, E.B.; Jönsson, G.B.; Frederick, D.T.; McHenry, K.; Porter, D.; Tran, T.-N.; Love, K.T.; Langer, R.; et al. BCL2A1 is a lineage-specific antiapoptotic melanoma oncogene that confers resistance to BRAF inhibition. Proc. Natl. Acad. Sci. USA 2013, 110, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Beuret, L.; Flori, E.; Denoyelle, C.; Bille, K.; Busca, R.; Picardo, M.; Bertolotto, C.; Ballotti, R. Up-regulation of MET Expression by α-Melanocyte-stimulating Hormone and MITF Allows Hepatocyte Growth Factor to Protect Melanocytes and Melanoma Cells from Apoptosis. J. Biol. Chem. 2007, 282, 14140–14147. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Fu, Y.; Meyskens, F.L., Jr. MiTF Regulates Cellular Response to Reactive Oxygen Species through Transcriptional Regulation of APE-1/Ref-1. J. Investig. Dermatol. 2009, 129, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Logsdon, D.P.; Grimard, M.; Luo, M.; Shahda, S.; Jiang, Y.; Tong, Y.; Yu, Z.; Zyromski, N.; Schipani, E.; Carta, F.; et al. Regulation of HIF1α under Hypoxia by APE1/Ref-1 Impacts CA9 Expression: Dual Targeting in Patient-Derived 3D Pancreatic Cancer Models. Mol. Cancer Ther. 2016, 15, 2722–2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buscà, R.; Berra, E.; Gaggioli, C.; Khaled, M.; Bille, K.; Marchetti, B.; Thyss, R.; Fitsialos, G.; Larribère, L.; Bertolotto, C.; et al. Hypoxia-inducible factor 1α is a new target of microphthalmia-associated transcription factor (MITF) in melanoma cells. J. Cell Biol. 2005, 170, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Caramel, J.; Papadogeorgakis, E.; Hill, L.; Browne, G.J.; Richard, G.; Wierinckx, A.; Saldanha, G.; Osborne, J.; Hutchinson, P.; Tse, G.; et al. A Switch in the Expression of Embryonic EMT-Inducers Drives the Development of Malignant Melanoma. Cancer Cell 2013, 24, 466–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandamme, N.; Denecker, G.; Bruneel, K.; Blancke, G.; Akay, O.; Taminau, J.; De Coninck, J.; De Smedt, E.; Skrypek, N.; Van Loocke, W.; et al. The EMT Transcription Factor ZEB2 Promotes Proliferation of Primary and Metastatic Melanoma While Suppressing an Invasive, Mesenchymal-Like Phenotype. Cancer Res. 2020, 80, 2983–2995. [Google Scholar] [CrossRef] [PubMed]
- Naffouje, S.; Naffouje, R.; Bhagwandin, S.; Salti, G.I. Microphthalmia transcription factor in malignant melanoma predicts occult sentinel lymph node metastases and survival. Melanoma Res. 2015, 25, 496–502. [Google Scholar] [CrossRef]
- Salti, G.I.; Manougian, T.; Farolan, M.; Shilkaitis, A.; Majumdar, D.; Das Gupta, T.K. Micropthalmia transcription factor: A new prognostic marker in intermediate-thickness cutaneous malignant melanoma. Cancer Res. 2000, 60, 5012–5016. [Google Scholar]
- Falletta, P.; del Campo, L.S.; Chauhan, J.; Effern, M.; Kenyon, A.; Kershaw, C.J.; Siddaway, R.; Lisle, R.J.; Freter, R.; Daniels, M.J.; et al. Translation reprogramming is an evolutionarily conserved driver of phenotypic plasticity and therapeutic resistance in melanoma. Genes Dev. 2017, 31, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Landsberg, J.; Kohlmeyer, J.; Renn, M.; Bald, T.; Rogava, M.; Cron, M.; Fatho, M.; Lennerz, V.; Wölfel, T.; Hölzel, M.; et al. Melanomas resist T-cell therapy through inflammation-induced reversible dedifferentiation. Nature 2012, 490, 412–416. [Google Scholar] [CrossRef]
- Riesenberg, S.; Groetchen, A.; Siddaway, R.; Bald, T.; Reinhardt, J.; Smorra, D.; Kohlmeyer, J.; Renn, M.; Phung, B.; Aymans, P.; et al. MITF and c-Jun antagonism interconnects melanoma dedifferentiation with pro-inflammatory cytokine responsiveness and myeloid cell recruitment. Nat. Commun. 2015, 6, 8755. [Google Scholar] [CrossRef]
- Goodall, J.; Carreira, S.; Denat, L.; Kobi, D.; Davidson, I.; Nuciforo, P.; Sturm, R.A.; LaRue, L.; Goding, C.R. Brn-2 represses microphthalmia-associated transcription factor expression and marks a distinct subpopulation of microphthalmia-associated transcription factor-negative melanoma cells. Cancer Res. 2008, 68, 7788–7794. [Google Scholar] [CrossRef] [Green Version]
- Boyle, G.M.; Woods, S.L.; Bonazzi, V.F.; Stark, M.S.; Hacker, E.; Aoude, L.G.; Dutton-Regester, K.; Cook, A.L.; Sturm, R.A.; Hayward, N.K. Melanoma cell invasiveness is regulated by miR-211 suppression of the BRN2 transcription factor. Pigment Cell Melanoma Res. 2011, 24, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.F.; Hanniford, D.; Menendez, S.; Reavie, L.; Zou, X.; Alvarez-Diaz, S.; Zakrzewski, J.; Blochin, E.; Rose, A.; Bogunovic, D.; et al. Aberrant miR-182 expression promotes melanoma metastasis by repressing FOXO3 and microphthalmia-associated transcription factor. Proc. Natl. Acad. Sci. USA 2009, 106, 1814–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivas-García, Y.; Falletta, P.; Liebing, J.; Louphrasitthiphol, P.; Feng, Y.; Chauhan, J.; Scott, D.A.; Glodde, N.; Calvo, A.C.; Bonham, S.; et al. Lineage-Restricted Regulation of SCD and Fatty Acid Saturation by MITF Controls Melanoma Phenotypic Plasticity. Mol. Cell 2019, 77, 120–137.e9. [Google Scholar] [CrossRef] [PubMed]
- Feige, E.; Yokoyama, S.; Levy, C.; Khaled, M.; Igras, V.; Lin, R.J.; Lee, S.; Widlund, H.R.; Granter, S.R.; Kung, A.L.; et al. Hypoxia-induced transcriptional repression of the melanoma-associated oncogene MITF. Proc. Natl. Acad. Sci. USA 2011, 108, E924–E933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.; Zbytek, B.; Slominski, R. Inhibitors of melanogenesis increase toxicity of cyclophosphamide and lymphocytes against melanoma cells. Int. J. Cancer 2009, 124, 1470–1477. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.; Kim, T.-K.; Brożyna, A.; Janjetovic, Z.; Brooks, D.; Schwab, L.; Skobowiat, C.; Jóźwicki, W.; Seagroves, T. The role of melanogenesis in regulation of melanoma behavior: Melanogenesis leads to stimulation of HIF-1α expression and HIF-dependent attendant pathways. Arch. Biochem. Biophys. 2014, 563, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Esparza, M.; Jiménez-Cervantes, C.; Beermann, F.; Aparicio, P.; Lozano, J.A.; Garcia-Borron, J.C. Transforming Growth Factor-β1 Inhibits Basal Melanogenesis in B16/F10 Mouse Melanoma Cells by Increasing the Rate of Degradation of Tyrosinase and Tyrosinase-related Protein-1. J. Biol. Chem. 1997, 272, 3967–3972. [Google Scholar] [CrossRef] [Green Version]
- Hoek, K.S.; Schlegel, N.C.; Brafford, P.; Sucker, A.; Ugurel, S.; Kumar, R.; Weber, B.L.; Nathanson, K.L.; Phillips, D.J.; Herlyn, M.; et al. Metastatic potential of melanomas defined by specific gene expression profiles with no BRAF signature. Pigment Cell Res. 2006, 19, 290–302. [Google Scholar] [CrossRef]
- Rambow, F.; Marine, J.-C.; Goding, C.R. Melanoma plasticity and phenotypic diversity: Therapeutic barriers and opportunities. Genes Dev. 2019, 33, 1295–1318. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.P.; Sanchez-Laorden, B.; O’Brien, K.; Brunton, H.; Ferguson, J.; Young, H.; Dhomen, N.; Flaherty, K.T.; Frederick, D.T.; Cooper, Z.A.; et al. The immune microenvironment confers resistance to MAPK pathway inhibitors through macrophage-derived TNFalpha. Cancer Discov. 2014, 4, 1214–1229. [Google Scholar] [CrossRef] [Green Version]
- Ohanna, M.; Giuliano, S.; Bonet, C.; Imbert, V.; Hofman, V.; Zangari, J.; Bille, K.; Robert, C.; Paillerets, B.B.-D.; Hofman, P.; et al. Senescent cells develop a PARP-1 and nuclear factor-κB-associated secretome (PNAS). Genes Dev. 2011, 25, 1245–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohanna, M.; Cheli, Y.; Bonet, C.; Bonazzi, V.F.; Allegra, M.; Giuliano, S.; Bille, K.; Bahadoran, P.; Giacchero, D.; Lacour, J.-P.; et al. Secretome from senescent melanoma engages the STAT3 pathway to favor reprogramming of naive melanoma towards a tumor-initiating cell phenotype. Oncotarget 2013, 4, 2212–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konieczkowski, D.J.; Johannessen, C.M.; Abudayyeh, O.; Kim, J.W.; Cooper, Z.A.; Piris, A.; Frederick, D.T.; Barzily-Rokni, M.; Straussman, R.; Haq, R.; et al. A Melanoma Cell State Distinction Influences Sensitivity to MAPK Pathway Inhibitors. Cancer Discov. 2014, 4, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada, C.; Mirabal-Ortega, L.; Méry, L.; Dingli, F.; Besse, L.; Messaoudi, C.; Loew, D.; Pouponnot, C.; Bertolotto, C.; Eychène, A.; et al. MITF activity is regulated by a direct interaction with RAF proteins in melanoma cells. Commun. Biol. 2022, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; Jóźwicki, W.; Carlson, J.A.; Slominski, A.T. Melanogenesis affects overall and disease-free survival in patients with stage III and IV melanoma. Hum. Pathol. 2013, 44, 2071–2074. [Google Scholar] [CrossRef] [Green Version]
- Brożyna, A.A.; Jóźwicki, W.; Roszkowski, K.; Filipiak, J.; Slominski, A.T. Melanin content in melanoma metastases affects the outcome of radiotherapy. Oncotarget 2016, 7, 17844–17853. [Google Scholar] [CrossRef] [Green Version]
- Slominski, R.M.; Sarna, T.; Płonka, P.M.; Raman, C.; Brożyna, A.A.; Slominski, A.T. Melanoma, Melanin, and Melanogenesis: The Yin and Yang Relationship. Front. Oncol. 2022, 12, 842496. [Google Scholar] [CrossRef]
- Ballotti, R.; Cheli, Y.; Bertolotto, C. The complex relationship between MITF and the immune system: A Melanoma ImmunoTherapy (response) Factor? Mol. Cancer 2020, 19, 170. [Google Scholar] [CrossRef]
- Arts, N.; Cané, S.; Hennequart, M.; Lamy, J.; Bommer, G.; Van Den Eynde, B.; De Plaen, E. microRNA-155, induced by interleukin-1ss, represses the expression of microphthalmia-associated transcription factor (MITF-M) in melanoma cells. PLoS ONE 2015, 10, e0122517. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Krijgsman, O.; Tsoi, J.; Robert, L.; Hugo, W.; Song, C.; Kong, X.; Possik, P.A.; Cornelissen-Steijger, P.D.; Foppen, M.H.G.; et al. Low MITF/AXL ratio predicts early resistance to multiple targeted drugs in melanoma. Nat. Commun. 2014, 5, 5712. [Google Scholar] [CrossRef]
- Fuhrmann, S.; Zou, C.; Levine, E.M. Retinal pigment epithelium development, plasticity, and tissue homeostasis. Exp. Eye Res. 2013, 123, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorgaleleh, S.; Naghipoor, K.; Barahouie, A.; Dastaviz, F.; Oladnabi, M. Molecular and biochemical mechanisms of human iris color: A comprehensive review. J. Cell. Physiol. 2020, 235, 8972–8982. [Google Scholar] [CrossRef] [PubMed]
- Sturm, R.A.; Larsson, M. Genetics of human iris colour and patterns. Pigment Cell Melanoma Res. 2009, 22, 544–562. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Hu, D.-N.; McCormick, S.A.; Ito, S. Characterization of melanin in human iridal and choroidal melanocytes from eyes with various colored irides. Pigment Cell Melanoma Res. 2008, 21, 97–105. [Google Scholar] [CrossRef]
- Sturm, R.A.; Frudakis, T.N. Eye colour: Portals into pigmentation genes and ancestry. Trends Genet. 2004, 20, 327–332. [Google Scholar] [CrossRef]
- Grønskov, K.; Ek, J.; Brondum-Nielsen, K. Oculocutaneous albinism. Orphanet J. Rare Dis. 2007, 2, 43. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; van Duijn, K.; Vingerling, J.R.; Hofman, A.; Uitterlinden, A.G.; Janssens, A.C.J.; Kayser, M. Eye color and the prediction of complex phenotypes from genotypes. Curr. Biol. 2009, 19, R192–R193. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wollstein, A.; Hysi, P.G.; Ankra-Badu, G.A.; Spector, T.D.; Park, D.; Zhu, G.; Larsson, M.; Duffy, D.L.; Montgomery, G.W.; et al. Digital quantification of human eye color highlights genetic association of three new loci. PLoS Genet. 2010, 6, e1000934. [Google Scholar] [CrossRef] [Green Version]
- Kayser, M.; Liu, F.; Janssens, A.C.J.; Rivadeneira, F.; Lao, O.; van Duijn, K.; Vermeulen, M.; Arp, P.; Jhamai, M.M.; van Ijcken, W.F.; et al. Three genome-wide association studies and a linkage analysis identify HERC2 as a human iris color gene. Am. J. Hum. Genet. 2008, 82, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Sulem, P.; Gudbjartsson, D.F.; Stacey, S.N.; Helgason, A.; Rafnar, T.; Magnusson, K.P.; Manolescu, A.; Karason, A.; Palsson, A.; Thorleifsson, G.; et al. Genetic determinants of hair, eye and skin pigmentation in Europeans. Nat. Genet. 2007, 39, 1443–1452. [Google Scholar] [CrossRef]
- Simcoe, M.; Valdes, A.; Liu, F.; Furlotte, N.A.; Evans, D.M.; Hemani, G.; Ring, S.M.; Smith, G.D.; Duffy, D.L.; Zhu, G.; et al. Genome-wide association study in almost 195,000 individuals identifies 50 previously unidentified genetic loci for eye color. Sci. Adv. 2021, 7, eabd1239. [Google Scholar] [CrossRef] [PubMed]
- Shibahara, S.; Takeda, K.; Yasumoto, K.-I.; Udono, T.; Watanabe, K.-I.; Saito, H.; Takahashi, K. Microphthalmia-associated transcription factor (MITF): Multiplicity in structure, function, and regulation. J. Investig. Dermatol. Symp. Proc. 2001, 6, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharti, K.; Liu, W.; Csermely, T.; Bertuzzi, S.; Arnheiter, H. Alternative promoter use in eye development: The complex role and regulation of the transcription factor MITF. Development 2008, 135, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Bharti, K.; Gasper, M.; Ou, J.; Brucato, M.; Clore-Gronenborn, K.; Pickel, J.; Arnheiter, H. A regulatory loop involving PAX6, MITF, and WNT signaling controls retinal pigment epithelium development. PLoS Genet. 2012, 8, e1002757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Llorca, A.; Aspelund, S.G.; Ogmundsdottir, M.H.; Steingrimsson, E.; Eysteinsson, T. The microphthalmia-associated transcription factor (Mitf) gene and its role in regulating eye function. Sci. Rep. 2019, 9, 15386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinisalo, M.; Putula, J.; Mannermaa, E.; Urtti, A.; Honkakoski, P. Regulation of the human tyrosinase gene in retinal pigment epithelium cells: The significance of transcription factor orthodenticle homeobox 2 and its polymorphic binding site. Mol. Vis. 2012, 18, 38–54. [Google Scholar] [PubMed]
- Maruotti, J.; Thein, T.; Zack, D.J.; Esumi, N. MITF-M, a ‘melanocyte-specific’ isoform, is expressed in the adult retinal pigment epithelium. Pigment Cell Melanoma Res. 2012, 25, 641–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flesher, J.L.; Paterson-Coleman, E.K.; Vasudeva, P.; Ruiz-Vega, R.; Marshall, M.; Pearlman, E.; MacGregor, G.R.; Neumann, J.; Ganesan, A.K. Delineating the role of MITF isoforms in pigmentation and tissue homeostasis. Pigment Cell Melanoma Res. 2020, 33, 279–292. [Google Scholar] [CrossRef]
- Shibuya, H.; Watanabe, R.; Maeno, A.; Ichimura, K.; Tamura, M.; Wakana, S.; Shiroishi, T.; Ohba, K.; Takeda, K.; Tomita, H.; et al. Melanocytes contribute to the vasculature of the choroid. Genes Genet. Syst. 2018, 93, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Vader, M.J.C.; Madigan, M.C.; Versluis, M.; Suleiman, H.M.; Gezgin, G.; Gruis, N.A.; Out-Luiting, J.J.; Bergman, W.; Verdijk, R.M.; Jager, M.J.; et al. GNAQ and GNA11 mutations and downstream YAP activation in choroidal nevi. Br. J. Cancer 2017, 117, 884–887. [Google Scholar] [CrossRef] [Green Version]
- Nell, R.J.; Menger, N.V.; Versluis, M.; Luyten, G.P.M.; Verdijk, R.M.; Madigan, M.C.; Jager, M.J.; Van der Velden, P.A. Involvement of mutant and wild-type CYSLTR2 in the development and progression of uveal and melanoma. BMC Cancer 2021, 21, 164. [Google Scholar] [CrossRef] [PubMed]
- Uribe, P.; Wistuba, I.; Gonzalez, S. BRAF Mutation: A Frequent Event in Benign, Atypical, and Malignant Melanocytic Lesions of the Skin. Am. J. Dermatopathol. 2003, 25, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Poynter, J.N.; Elder, J.T.; Fullen, D.R.; Nair, R.P.; Soengas, M.S.; Johnson, T.M.; Redman, B.; Thomas, N.E.; Gruber, S.B. BRAF and NRAS mutations in melanoma and melanocytic nevi. Melanoma Res. 2006, 16, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Emley, A.; Nguyen, L.P.; Yang, S.; Mahalingam, M. Somatic mutations in GNAQ in amelanotic/hypomelanotic blue nevi. Hum. Pathol. 2011, 42, 136–140. [Google Scholar] [CrossRef]
- Rodríguez-Jiménez, P.; Mayor-Sanabria, F.; Rütten, A.; Fraga, J.; Llamas-Velasco, M. Agminated blue nevus: GNAQ mutations and beyond. Actas Dermo-Sifiliográficas (Engl. Ed.) 2021, 112, 95–97. [Google Scholar] [CrossRef]
- Singh, A.D.; De Potter, P.; Fijal, B.A.; Shields, C.L.; Shields, J.A.; Elston, R.C. Lifetime prevalence of uveal melanoma in white patients with oculo(dermal) melanocytosis. Ophthalmology 1998, 105, 195–198. [Google Scholar] [CrossRef]
- Vivancos, A.; Caratú, G.; Matito, J.; Muñoz, E.; Ferrer, B.; Hernandez-Losa, J.; Bodet, D.; Pérez-Alea, M.; Cortes, J.; Garcia-Patos, V.; et al. Genetic evolution of nevus of Ota reveals clonal heterogeneity acquiring BAP1 andTP53mutations. Pigment Cell Melanoma Res. 2016, 29, 247–253. [Google Scholar] [CrossRef]
- Rivolta, C.; Royer-Bertrand, B.; Rimoldi, D.; Schalenbourg, A.; Zografos, L.; Leyvraz, S.; Moulin, A. UV light signature in conjunctival melanoma; not only skin should be protected from solar radiation. J. Hum. Genet. 2016, 61, 361–362. [Google Scholar] [CrossRef] [Green Version]
- Johansson, P.A.; Brooks, K.; Newell, F.; Palmer, J.M.; Wilmott, J.S.; Pritchard, A.L.; Broit, N.; Wood, S.; Carlino, M.S.; Leonard, C.; et al. Whole genome landscapes of uveal melanoma show an ultraviolet radiation signature in iris tumours. Nat. Commun. 2020, 11, 2408. [Google Scholar] [CrossRef]
- Van Raamsdonk, C.D.; Bezrookove, V.; Green, G.; Bauer, J.; Gaugler, L.; O’Brien, J.M.; Simpson, E.M.; Barsh, G.S.; Bastian, B.C. Frequent somatic mutations of GNAQ in uveal melanoma and blue naevi. Nature 2009, 457, 599–602. [Google Scholar] [CrossRef] [Green Version]
- Van Raamsdonk, C.D.; Griewank, K.G.; Crosby, M.B.; Garrido, M.C.; Vemula, S.; Wiesner, T.; Obenauf, A.C.; Wackernagel, W.; Green, G.; Bouvier, N.; et al. Mutations in GNA11 in Uveal Melanoma. N. Engl. J. Med. 2010, 363, 2191–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, K.N.; Jager, M.J.; de Klein, A.; Kiliҫ, E. Uveal melanoma: Towards a molecular understanding. Prog. Retin. Eye Res. 2020, 75, 100800. [Google Scholar] [CrossRef] [PubMed]
- Harbour, J.W.; Onken, M.D.; Roberson, E.D.O.; Duan, S.; Cao, L.; Worley, L.A.; Council, M.L.; Matatall, K.A.; Helms, C.; Bowcock, A.M. Frequent Mutation of BAP1 in Metastasizing Uveal Melanomas. Science 2010, 330, 1410–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouriaux, F.; Vincent, S.; Kherrouche, Z.; Maurage, C.-A.; Planque, N.; Monté, D.; Labalette, P.; Saule, S. Microphthalmia transcription factor analysis in posterior uveal melanomas. Exp. Eye Res. 2003, 76, 653–661. [Google Scholar] [CrossRef]
- Manderfield, L.J.; Engleka, K.A.; Aghajanian, H.; Gupta, M.; Yang, S.; Li, L.; Baggs, J.E.; Hogenesch, J.B.; Olson, E.N.; Epstein, J.A. Pax3 and Hippo Signaling Coordinate Melanocyte Gene Expression in Neural Crest. Cell Rep. 2014, 9, 1885–1895. [Google Scholar] [CrossRef] [Green Version]
- Lyubasyuk, V.; Ouyang, H.; Yu, F.-X.; Guan, K.-L.; Zhang, K. YAP inhibition blocks uveal melanogenesis driven by GNAQ or GNA11 mutations. Mol. Cell. Oncol. 2015, 2, e970957. [Google Scholar] [CrossRef]
- Brouwer, N.J.; Konstantinou, E.K.; Gragoudas, E.S.; Marinkovic, M.; Luyten, G.P.M.; Kim, I.K.; Jager, M.J.; Vavvas, D.G. Targeting the YAP/TAZ Pathway in Uveal and Conjunctival Melanoma with Verteporfin. Investig. Opthalmol. Vis. Sci. 2021, 62, 3. [Google Scholar] [CrossRef]
- Chen, X.; Wang, J.; Shen, H.; Lu, J.; Li, C.; Hu, D.-N.; Dong, X.D.; Yan, D.; Tu, L. Epigenetics, MicroRNAs, and Carcinogenesis: Functional Role of MicroRNA-137 in Uveal Melanoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1193–1199. [Google Scholar] [CrossRef]
- Yan, D.; Dong, X.D.; Chen, X.; Yao, S.; Wang, L.; Wang, J.; Wang, C.; Hu, D.-N.; Qu, J.; Tu, L. Role of MicroRNA-182 in Posterior Uveal Melanoma: Regulation of Tumor Development through MITF, BCL2 and Cyclin D2. PLoS ONE 2012, 7, e40967. [Google Scholar] [CrossRef] [Green Version]
- Matatall, K.A.; Agapova, O.A.; Onken, M.D.; Worley, L.A.; Bowcock, A.M.; Harbour, J.W. BAP1 deficiency causes loss of melanocytic cell identity in uveal melanoma. BMC Cancer 2013, 13, 371. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.K.; Singh, L.; Pushker, N.; Saini, N.; Meel, R.; Chosdol, K.; Bakhshi, S.; Sen, S.; Venkatesh, P.; Chawla, B.; et al. Identification of canonical NFκB (C-NFκB) pathway in uveal melanoma and their relation with patient outcome. Clin. Exp. Metastasis 2019, 36, 271–290. [Google Scholar] [CrossRef] [PubMed]
- Materin, M.A.; Simões, C.; Bergeron, M.-A.; Champagne, S.; Gaudreault, M.; Deschambeault, A.; Landreville, S. Faculty of 1000 evaluation for Repression of genes involved in melanocyte differentiation in uveal melanoma. Mol. Vis. 2012, 18, 1813–1822. [Google Scholar] [CrossRef]
- Durante, M.A.; Rodriguez, D.A.; Kurtenbach, S.; Kuznetsov, J.N.; Sanchez, M.I.; Decatur, C.L.; Snyder, H.; Feun, L.G.; Livingstone, A.S.; Harbour, J.W. Single-cell analysis reveals new evolutionary complexity in uveal melanoma. Nat. Commun. 2020, 11, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandiani, C.; Strub, T.; Nottet, N.; Cheli, Y.; Gambi, G.; Bille, K.; Husser, C.; Dalmasso, M.; Béranger, G.; Lassalle, S.; et al. Single-cell RNA sequencing reveals intratumoral heterogeneity in primary uveal melanomas and identifies HES6 as a driver of the metastatic disease. Cell Death Differ. 2021, 28, 1990–2000. [Google Scholar] [CrossRef]
- Strub, T.; Martel, A.; Nahon-Esteve, S.; Baillif, S.; Ballotti, R.; Bertolotto, C. Translation of single-cell transcriptomic analysis of uveal melanomas to clinical oncology. Prog. Retin. Eye Res. 2021, 85, 100968. [Google Scholar] [CrossRef]
- Kim, E.; Zucconi, B.E.; Wu, M.; Nocco, S.E.; Meyers, D.J.; McGee, J.S.; Venkatesh, S.; Cohen, D.L.; Gonzalez, E.C.; Ryu, B.; et al. MITF Expression Predicts Therapeutic Vulnerability to p300 Inhibition in Human Melanoma. Cancer Res. 2019, 79, 2649–2661. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, S.; Feige, E.; Poling, L.L.; Levy, C.; Widlund, H.R.; Khaled, M.; Kung, A.L.; Fisher, D.E. Pharmacologic suppression of MITF expression via HDAC inhibitors in the melanocyte lineage. Pigment Cell Melanoma Res. 2008, 21, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Sundaramurthi, H.; García-Mulero, S.; Tonelotto, V.; Slater, K.; Marcone, S.; Piulats, J.M.; Watson, R.W.; Tobin, D.J.; Jensen, L.D.; Kennedy, B.N. Uveal Melanoma Cell Line Proliferation Is Inhibited by Ricolinostat, a Histone Deacetylase Inhibitor. Cancers 2022, 14, 782. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the cascade of melanin synthesis. TYR = tyrosinase, DCT = dopachrome tautomerase, TYRP-1 = tyrosinase-related protein 1, L-DOPA = dihydroxyphenylalanine, DHI = dihydroxyindole, DHICA = dihydroxindole-2-carboxylic acid, 5-s-CD/GD = 5-S-cysteinyldopa or glutathionyldopa, CD-quinones = cysteinyldopa-quinones.
Figure 1.
Schematic representation of the cascade of melanin synthesis. TYR = tyrosinase, DCT = dopachrome tautomerase, TYRP-1 = tyrosinase-related protein 1, L-DOPA = dihydroxyphenylalanine, DHI = dihydroxyindole, DHICA = dihydroxindole-2-carboxylic acid, 5-s-CD/GD = 5-S-cysteinyldopa or glutathionyldopa, CD-quinones = cysteinyldopa-quinones.

Figure 2.
Overview of the main targets of MITF. MITF = microphthalmia-associated transcription factor, PMEL = premelanosome protein, MLANA = Melan-A, Rab27a = RAB27A, member RAS oncogene family, TBX2 = T-box transcription factor 2, CDK2 = cyclin dependent kinase 2, BCL2 = BCL2 apoptosis regulator, BIRC7 = baculoviral IAP repeat containing 7, MET = MET proto-oncogene, receptor tyrosine kinase, HIF1α = hypoxia-inducible factor 1 alpha, SNAI2 = snail family transcriptional repressor 2, OA1 = ocular albinism 1, TYR = tyrosinase, DCT = dopachrome tautomerase, TYRP-1 = tyrosinase-related protein 1.
Figure 2.
Overview of the main targets of MITF. MITF = microphthalmia-associated transcription factor, PMEL = premelanosome protein, MLANA = Melan-A, Rab27a = RAB27A, member RAS oncogene family, TBX2 = T-box transcription factor 2, CDK2 = cyclin dependent kinase 2, BCL2 = BCL2 apoptosis regulator, BIRC7 = baculoviral IAP repeat containing 7, MET = MET proto-oncogene, receptor tyrosine kinase, HIF1α = hypoxia-inducible factor 1 alpha, SNAI2 = snail family transcriptional repressor 2, OA1 = ocular albinism 1, TYR = tyrosinase, DCT = dopachrome tautomerase, TYRP-1 = tyrosinase-related protein 1.

Figure 3.
Schematic representation of the pathways that lead to MITF activation. α-MSH = α-melanocyte-stimulating hormone, MC1R = melanocortin 1 receptor, c-AMP = cyclin adenosine monophosphate, PKA = protein kinase A, CREB = cAMP response element, SOX10 = SRY-Box transcription factor 10, PAX3 = paired box 3, SCF = stem cell factor, ERK = extracellular signal-regulated kinase, LEF/TCF = lymphoid enhancer-binding factor/T cell factor, MITF = microphthalmia-associated transcription factor, TYR = tyrosinase, DCT = dopachrome tautomerase, TYRP-1 = tyrosinase-related protein 1.
Figure 3.
Schematic representation of the pathways that lead to MITF activation. α-MSH = α-melanocyte-stimulating hormone, MC1R = melanocortin 1 receptor, c-AMP = cyclin adenosine monophosphate, PKA = protein kinase A, CREB = cAMP response element, SOX10 = SRY-Box transcription factor 10, PAX3 = paired box 3, SCF = stem cell factor, ERK = extracellular signal-regulated kinase, LEF/TCF = lymphoid enhancer-binding factor/T cell factor, MITF = microphthalmia-associated transcription factor, TYR = tyrosinase, DCT = dopachrome tautomerase, TYRP-1 = tyrosinase-related protein 1.

Figure 4.
Schematic representation of the putative effect of BRAF mutation on pseudo-epithelial-mesenchymal transition and MITF in cutaneous melanoma. MITF = microphthalmia-associated transcription factor, SNAI2 = snail family transcriptional repressor 2, ZEB2 = zinc finger E-box binding homeobox 2, ZEB1 = zinc finger E-box binding homeobox 1, TWIST1 = twist family BHLH transcription factor 1, BRAF = B-Raf proto-oncogene, serine/threonine kinase. The size of the boxes represents the level of expression (higher expression = bigger box; lower expression = smaller box).
Figure 4.
Schematic representation of the putative effect of BRAF mutation on pseudo-epithelial-mesenchymal transition and MITF in cutaneous melanoma. MITF = microphthalmia-associated transcription factor, SNAI2 = snail family transcriptional repressor 2, ZEB2 = zinc finger E-box binding homeobox 2, ZEB1 = zinc finger E-box binding homeobox 1, TWIST1 = twist family BHLH transcription factor 1, BRAF = B-Raf proto-oncogene, serine/threonine kinase. The size of the boxes represents the level of expression (higher expression = bigger box; lower expression = smaller box).


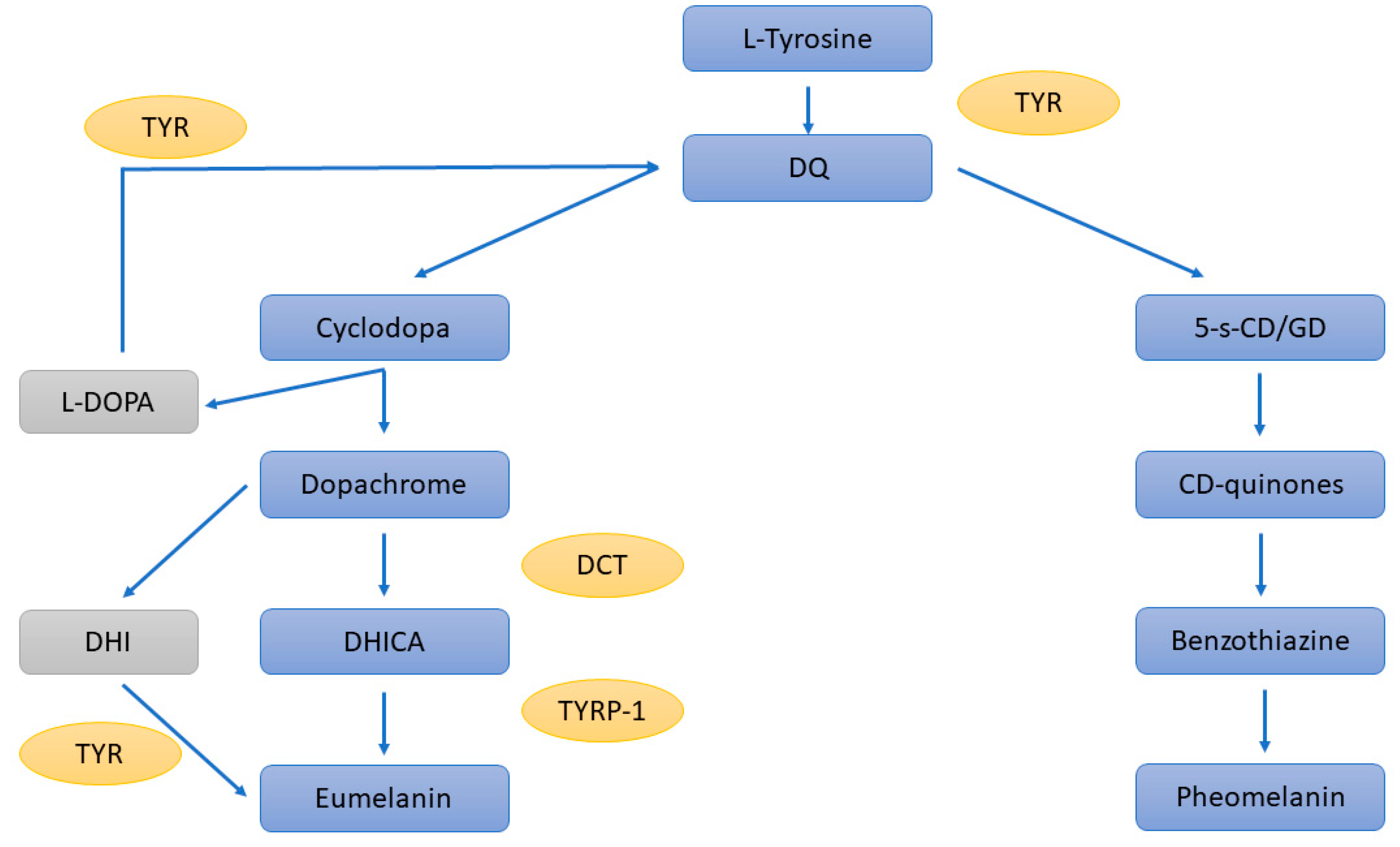{kind=link}
{kind=link}
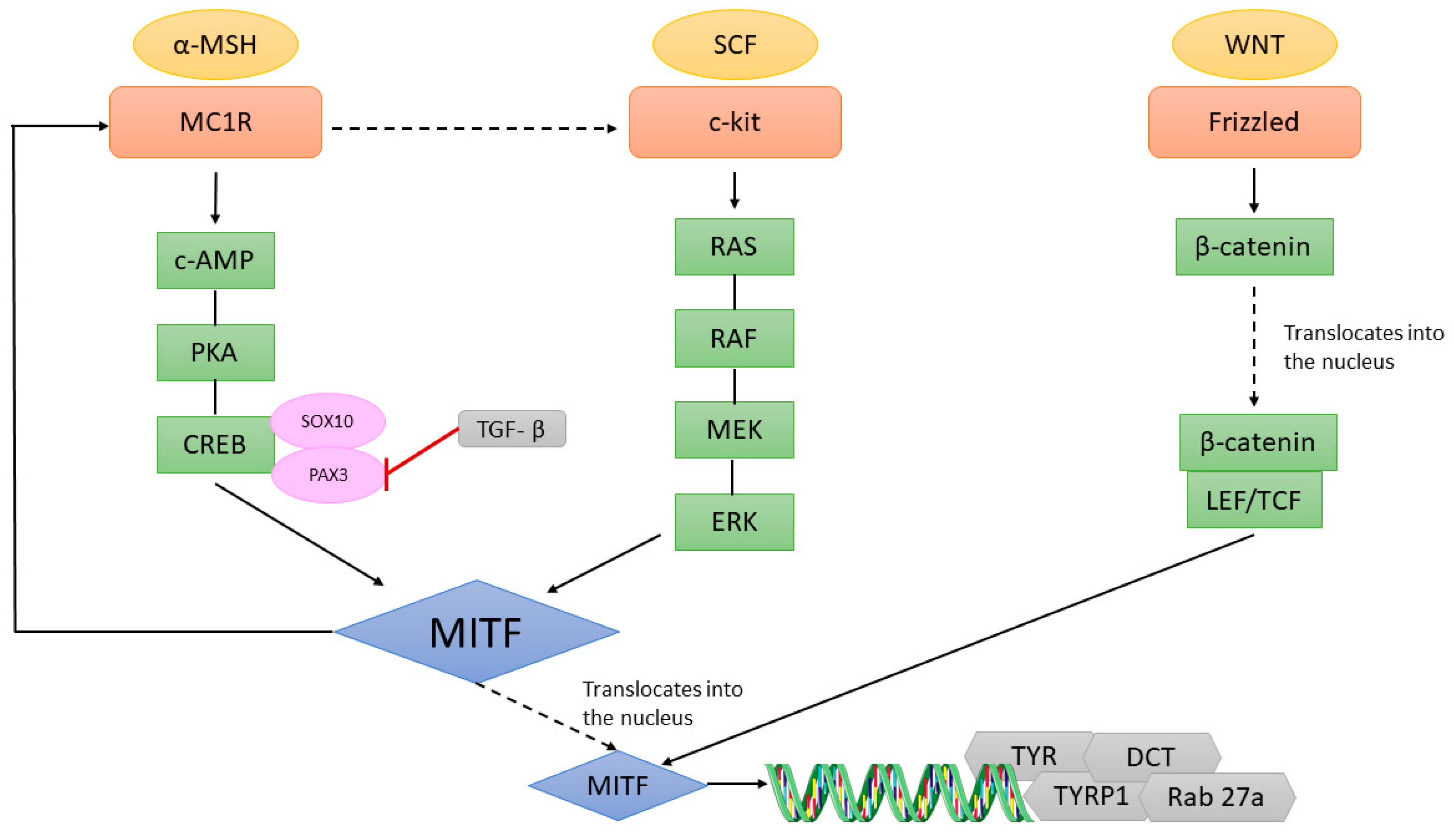{kind=link}
{kind=link}
Table 1.
Distribution of MITF isoforms in eye tissues.
| Isoform | Location in the Eye |
|---|---|
| MITF-A | Retina, RPE, choroid |
| MITF-D | RPE |
| MITF-H | RPE |
| MITF-J | Retina, RPE, choroid |
| MITF-M | Choroid and iris, variable results in RPE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gelmi, M.C.; Houtzagers, L.E.; Strub, T.; Krossa, I.; Jager, M.J. MITF in Normal Melanocytes, Cutaneous and Uveal Melanoma: A Delicate Balance. Int. J. Mol. Sci. 2022, 23, 6001. https://doi.org/10.3390/ijms23116001
AMA Style
Gelmi MC, Houtzagers LE, Strub T, Krossa I, Jager MJ. MITF in Normal Melanocytes, Cutaneous and Uveal Melanoma: A Delicate Balance. International Journal of Molecular Sciences. 2022; 23(11):6001. https://doi.org/10.3390/ijms23116001
Chicago/Turabian StyleGelmi, Maria Chiara, Laurien E. Houtzagers, Thomas Strub, Imène Krossa, and Martine J. Jager. 2022. "MITF in Normal Melanocytes, Cutaneous and Uveal Melanoma: A Delicate Balance" International Journal of Molecular Sciences 23, no. 11: 6001. https://doi.org/10.3390/ijms23116001
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.