
Serotonin (5-HT) 2A Receptor Involvement in Melanin Synthesis and Transfer via Activating the PKA/CREB Signaling Pathway

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Procedures
2.1. Ethical Statements
2.2. Zebrafish Lines and Maintenance
2.3. Cell Culture and Cell Viability Assay
2.4. Drug Treatment and Docking Analysis
2.5. Masson–Fontana Staining
2.6. Melanin Content and Tyrosinase Activity Assay
2.7. Western Blot
2.8. Melanocyte Dendrites’ Number and Length
2.9. Skin Tissue Culture and H&E Staining
2.10. Generation of Zebrafish Mutants with CRISPR/Cas9
2.11. Measurement of the Pigmenting Activity in the Zebrafish
2.12. Cytoskeleton Staining
2.13. Cell Scratch Assay
2.14. Whole-Mount In Situ Hybridization
2.15. Statistics
3. Results
3.1. HTR2A Mediates the Melanogenesis Induction of Serotonin in Both B16F10 Cell Line and Zebrafish Embryos
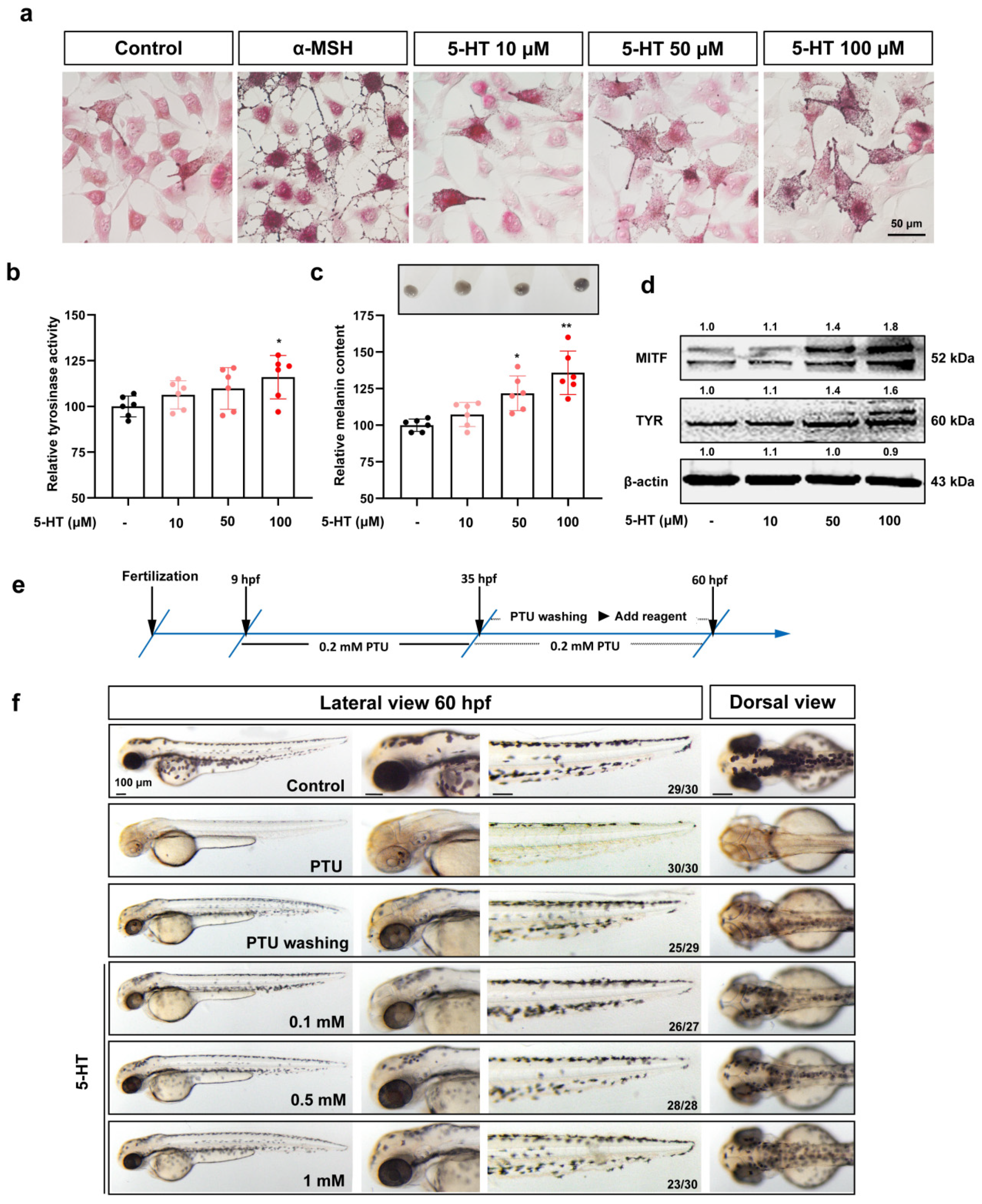
3.2. The Agonists of HTR2A Promote Melanogenesis in B16F10 Cell, Cultured Human Skin Tissue, and Zebrafish Embryos
3.3. Htr2a Deficiency Inhibits the Melanin Promotion Effect of Serotonin and HTR2A Agonist DOI in Zebrafish
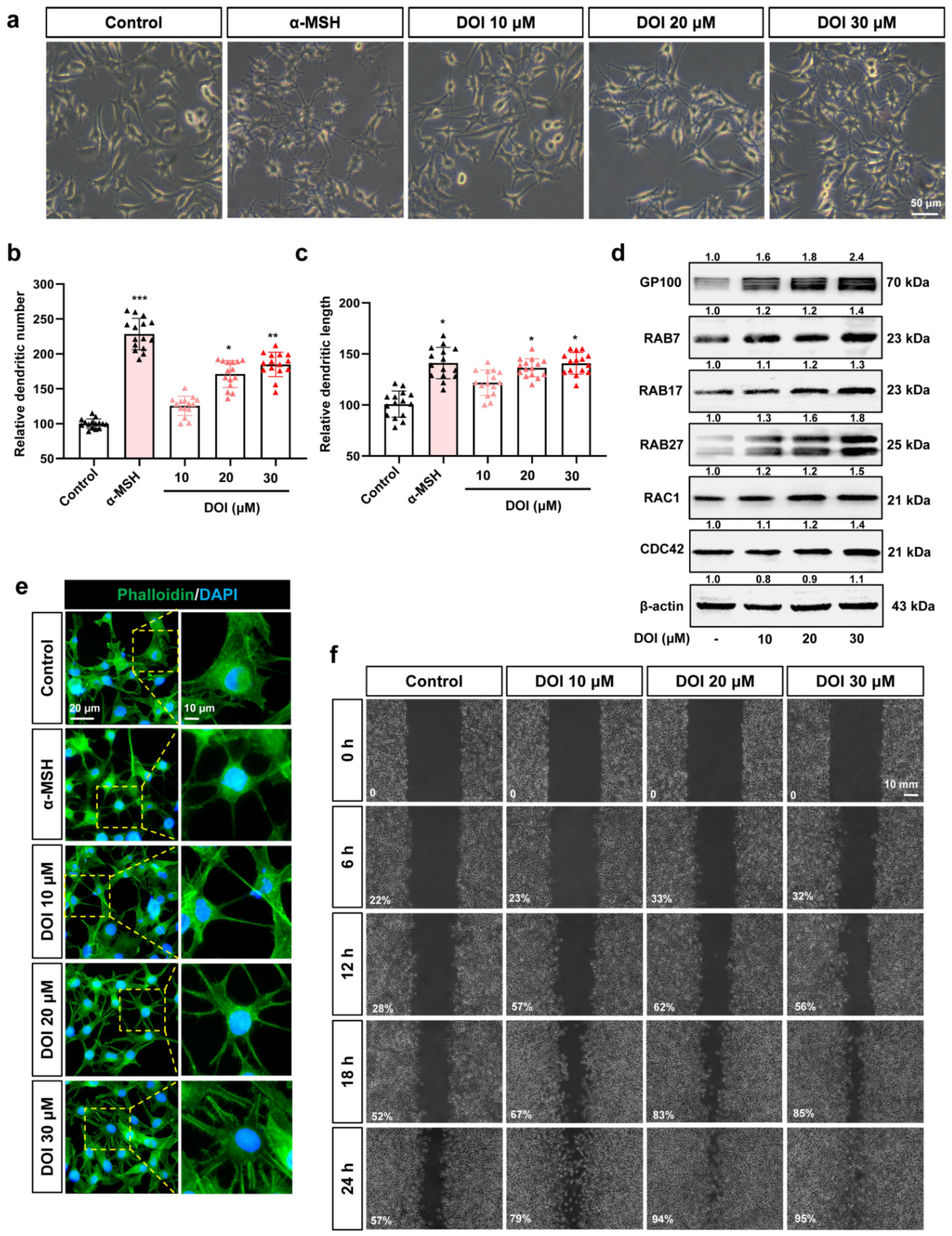
3.4. HTR2A Agonists Induce Melanosome Transport by Increasing the Melanocyte Dendrites and Migration
3.5. PKA/CREB Signaling Plays an Essential Role in the Downstream of HTR2A to Regulate Melanocyte Function
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations and Nomenclature
| 5-HT | serotonin (5-hydroxytryptamine) |
| HTR2A | serotonin receptor 2A |
| DOI | 4-iodo-2,5-dimethoxy-α-methylbenzeneethanamine hydrochloride |
| TCB-2 | 4-bromo-3,6-dimethoxybenzocyclobuten-1-yl methylamine hydrobromide |
| ketanserin tartrate | 3-[2-[4-(4-fluorobenzoyl)-1-piperidinyl]ethyl]-2,4[1H,3H]-quinazolinedione tartrate |
| AT1015 | N-[2-[4-(5H-dibenzo[a,d]cyclohepten-5-ylidene)-1-piperidinyl]ethyl]-1-formyl-4-piperidinecarboxamide hydrochloride |
| NE | Norepinephrine |
| EP | Epinephrine |
| GABA | gamma-aminobutyric acid |
| HA | histamine |
| GPCR | G protein-coupled receptors |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FBS | fetal bovine serum |
| sgRNA | single guide RNA |
| PTU | phenylthiourea |
| siRNA | small interference RNA |
| PLC | phospholipase C |
| DAG | diacylglycerol |
| IP3 | inositol triphosphate |
| PKC | protein kinase C |
| PKA | protein kinase A |
| hpf | hours post fertilization |
References
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV Light Touches the Brain and Endocrine System Through Skin, and Why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Fistarol, S.K.; Itin, P.H. Disorders of pigmentation. JDDG 2010, 8, 187–201. [Google Scholar] [CrossRef]
- Bertolesi, G.E.; Vazhappilly, S.T.; Hehr, C.L.; McFarlane, S. Pharmacological induction of skin pigmentation unveils the neuroendocrine circuit regulated by light. Pigment Cell Melanoma Res. 2016, 29, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Brenner, M.; Hearing, V.J. The Regulation of Skin Pigmentation. J. Biol. Chem. 2007, 282, 27557–27561. [Google Scholar] [CrossRef]
- Enkhtaivan, E.; Lee, C. Role of Amine Neurotransmitters and Their Receptors in Skin Pigmentation: Therapeutic Implication. Int. J. Mol. Sci. 2021, 22, 8071. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin Pigmentation in Mammalian Skin and Its Hormonal Regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Wortsman, J.; Szczesniewski, A.; Slugocki, G.; McNulty, J.; Kauser, S.; Tobin, D.J.; et al. Serotoninergic and melatoninergic systems are fully expressed in human skin. FASEB J. 2002, 16, 896–898. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Johansson, O.; Jing, C.; Semak, I.; Slugocki, G.; Wortsman, J. Tryptophan hydroxylase expression in human skin cells. Biochim. Biophys. Acta Mol. Basis Dis. 2003, 1639, 80–86. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Tobin, D.J. The cutaneous serotoninergic/melatoninergic system: Securing a place under the sun. FASEB J. 2005, 19, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, S.M.; Akkerman, J.W.; Nijmeijer, B.; Sixma, J.J.; Witkop, C.J.; White, J. The Hermansky-Pudlak syndrome. Evidence for a lowered 5-hydroxytryptamine content in platelets of heterozygotes. Scand. J. Haematol. 1977, 18, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Wu, H.; Wang, Q.; Cai, M.; Shi, W.; Shang, J. Chronic Stress Suppresses the Expression of Cutaneous Hypothalamic–Pituitary–Adrenocortical Axis Elements and Melanogenesis. PLoS ONE 2014, 9, e98283. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-L.; Pang, S.-L.; Liu, Q.-Z.; Wang, Q.; Cai, M.-X.; Shang, J. 5-HT1A/1B Receptors as Targets for Optimizing Pigmentary Responses in C57BL/6 Mouse Skin to Stress. PLoS ONE 2014, 9, e89663. [Google Scholar] [CrossRef]
- Zhou, L.; Cai, M.; Ren, Y.; Wu, H.; Liu, M.; Chen, H.; Shang, J. The different roles of 5-HT1A/2A receptors in fluoxetine ameliorated pigmentation of C57BL/6 mouse skin in response to stress. J. Dermatol. Sci. 2018, 92, 222–229. [Google Scholar] [CrossRef]
- Liu, L.; Fu, M.; Pei, S.; Zhou, L.; Shang, J. R-Fluoxetine Increases Melanin Synthesis Through a 5-HT1A/2A Receptor and p38 MAPK Signaling Pathways. Int. J. Mol. Sci. 2018, 20, 80. [Google Scholar] [CrossRef]
- Cheret, J.; Gherardini, J.; Bertolini, M.; Paus, R. Fluoxetine promotes human hair follicle pigmentation ex vivo: Serotonin reuptake inhibition as a new antigreying strategy? Br. J. Dermatol. 2020, 182, 1492–1494. [Google Scholar] [CrossRef]
- Liao, S.; Shang, J.; Tian, X.; Fan, X.; Shi, X.; Pei, S.; Wang, Q.; Yu, B. Up-regulation of melanin synthesis by the antidepressant fluoxetine. Exp. Dermatol. 2012, 21, 635–637. [Google Scholar] [CrossRef]
- Lee, H.; Park, M.; Kim, S.; Choo, H.P.; Lee, A.; Lee, C. Serotonin induces melanogenesis via serotonin receptor 2A. Br. J. Dermatol. 2011, 165, 1344–1348. [Google Scholar] [CrossRef]
- Kim, K.; Che, T.; Panova, O.; DiBerto, J.F.; Lyu, J.; Krumm, B.E.; Wacker, D.; Robertson, M.J.; Seven, A.B.; Nichols, D.E.; et al. Structure of a Hallucinogen-Activated Gq-Coupled 5-HT2A Serotonin Receptor. Cell 2020, 182, 1574–1588. [Google Scholar] [CrossRef]
- Liu, L.; Zhong, M.; Dong, J.; Chen, M.; Shang, J.; Yue, Y. 5-Hydroxytryptamine (5-HT) Positively Regulates Pig-mentation via Inducing Melanoblast Specification and Melanin Synthesis in Zebrafish Embryos. Biomolecules 2020, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Kurfurst, R.; Nizard, C.; Schnebert, S.; Perrier, E.; Tobin, D.J. Melanin transfer in human skin cells is mediated by filopodia—A model for homotypic and heterotypic lysosome-related organelle transfer. FASEB J. 2010, 24, 3756–3769. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Hammer, J.A. Melanosome transfer: It is best to give and receive. Curr. Opin. Cell Biol. 2014, 29, 1–7. [Google Scholar] [CrossRef]
- Fukuda, M. Rab GTPases: Key players in melanosome biogenesis, transport, and transfer. Pigment Cell Melanoma Res. 2021, 34, 222–235. [Google Scholar] [CrossRef]
- Pytliak, M.; Vargova, V.; Mechirova, V.; Felsoci, M. Serotonin receptors-from molecular biology to clinical applications. Physiol. Res. 2011, 60, 15–25. [Google Scholar] [CrossRef]
- Yohn, C.N.; Gergues, M.M.; Samuels, B.A. The role of 5-HT receptors in depression. Mol. Brain 2017, 10, 28. [Google Scholar] [CrossRef]
- Barbanti, P.; Aurilia, C.; Egeo, G.; Fofi, L.; Palmirotta, R. Serotonin receptor targeted therapy for migraine treatment: An overview of drugs in phase I and II clinical development. Expert Opin. Investig. Drugs 2017, 26, 269–277. [Google Scholar] [CrossRef]
- Patel, R.; Dickenson, A.H. Modality selective roles of pro-nociceptive spinal 5-HT2A and 5-HT3 receptors in normal and neuropathic states. Neuropharmacology 2018, 143, 29–37. [Google Scholar] [CrossRef]
- Nascimento, E.B.; Seniuk, J.G.T.; Godin, A.M.; Ferreira, W.; Dutra, M.B.; Oliveira, A.C.P.; Bastos, L.; Fiebich, B.L.; Coelho, M.M. Peripheral 5-HT1B and 5-HT2A receptors mediate the nociceptive response induced by 5-hydroxytryptamine in mice. Pharmacol. Biochem. Behav. 2011, 99, 598–603. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Zbytek, B.; Tobin, D.; Wortsman, J. Functional activity of serotoninergic and melatoninergic systems expressed in the skin. J. Cell. Physiol. 2003, 196, 144–153. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Salem, M.A.; Gibbons, N.C.; Maitland, D.J.; Marsch, E.; Elwary, S.M.; Healey, A.R. Blunted epidermal L-tryptophan metabolism in vitiligo affects immune response and ROS scavenging by Fenton chemistry, part 2: Epidermal H2O2/ONOO(-)-mediated stress in vitiligo hampers indoleamine 2,3-dioxygenase and aryl hydrocarbon receptor-mediated immune response signaling. FASEB J. 2012, 26, 2471–2485. [Google Scholar] [PubMed]
- Schallreuter, K.U.; Salem, M.A.; Gibbons, N.C.; Martinez, A.; Slominski, R.; Ludemann, J.; Rokos, H. Blunted epidermal L-tryptophan metabolism in vitiligo affects immune response and ROS scavenging by Fenton chemistry, part 1: Epidermal H2O2/ONOO(-)-mediated stress abrogates tryptophan hydroxylase and dopa decarboxylase activities, leading to low serotonin and melatonin levels. FASEB J. 2012, 26, 2457–2470. [Google Scholar] [PubMed]
- Kurbanov, K.; Berezov, T.T. Tryptophan metabolism in vitiligo. Vopr. Meditsinskoi Khimii 1976, 22, 683–687. [Google Scholar]
- Liao, S.; Lv, J.; Zhou, J.; Kalavagunta, P.K.; Shang, J. Effects of two chronic stresses on mental state and hair follicle melanogenesis in mice. Exp. Dermatol. 2017, 26, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Berg, K.A.; Harvey, J.A.; Spampinato, U.; Clarke, W.P. Physiological relevance of constitutive activity of 5-HT2A and 5-HT2C receptors. Trends Pharmacol. Sci. 2005, 26, 625–630. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, Y.; Zhong, M.; An, X.; Feng, Q.; Lai, Y.; Yu, M.; Zhang, X.; Liao, Z.; Chen, M.; Dong, J.; et al. Serotonin (5-HT) 2A Receptor Involvement in Melanin Synthesis and Transfer via Activating the PKA/CREB Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 6111. https://doi.org/10.3390/ijms23116111
Yue Y, Zhong M, An X, Feng Q, Lai Y, Yu M, Zhang X, Liao Z, Chen M, Dong J, et al. Serotonin (5-HT) 2A Receptor Involvement in Melanin Synthesis and Transfer via Activating the PKA/CREB Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(11):6111. https://doi.org/10.3390/ijms23116111
Chicago/Turabian StyleYue, Yunyun, Min Zhong, Xiaohong An, Qingyuan Feng, Yifan Lai, Meng Yu, Xiaofeng Zhang, Zixian Liao, Minghan Chen, Jing Dong, and et al. 2022. "Serotonin (5-HT) 2A Receptor Involvement in Melanin Synthesis and Transfer via Activating the PKA/CREB Signaling Pathway" International Journal of Molecular Sciences 23, no. 11: 6111. https://doi.org/10.3390/ijms23116111