
Identification of a Putative CodY Regulon in the Gram-Negative Phylum Synergistetes
by
 , , and
, , and
Jianing Geng
1,† ,
,
Sainan Luo
1,2,†,
Hui-Ru Shieh
3,
Hsing-Yi Wang
3,
Songnian Hu
1,2,* and
Yi-Ywan M. Chen
3,4,5,* 1
State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
3
Department of Microbiology and Immunology, College of Medicine, Chang Gung University, Taoyuan 333, Taiwan
4
Graduate Institute of Biomedical Sciences, College of Medicine, Chang Gung University, Taoyuan 333, Taiwan
5
Molecular Infectious Disease Research Center, Chang Gung Memorial Hospital, Taoyuan 333, Taiwan
*
Authors to whom correspondence should be addressed.
†
These authors have contributed equally to this work.
Int. J. Mol. Sci. 2022, 23(14), 7911; https://doi.org/10.3390/ijms23147911
Submission received: 16 June 2022
/
Revised: 5 July 2022
/
Accepted: 6 July 2022
/
Published: 18 July 2022
(This article belongs to the Special Issue Gene Expression Regulation in Microorganisms: Molecular Mechanisms, Systems and Synthetic Approaches)
Abstract
:CodY is a dominant regulator in low G + C, Gram-positive Firmicutes that governs the regulation of various metabolic pathways and cellular processes. By using various bioinformatics analyses and DNA affinity precipitation assay (DAPA), this study confirmed the presence of CodY orthologues and corresponding regulons in Gram-negative Synergistetes. A novel palindromic sequence consisting of AT-rich arms separated by a spacer region of variable length and sequence was identified in the promoters of the putative codY-containing operons in Synergistetes. The consensus sequence from genera Synergistes and Cloacibacillus (5′-AATTTTCTTAAAATTTCSCTTGATATTTACAATTTT) contained three AT-rich regions, resulting in two palindromic sequences; one of which is identical to Firmicutes CodY box (5′-AATTTTCWGAAAATT). The function of the consensus sequence was tested by using a recombinant CodY protein (His-CodYDSM) of Cloacibacillus evryensis DSM19522 in DAPA. Mutations in the central AT-rich sequence reduced significantly the binding of His-CodYDSM, whereas mutations in the 5′ or 3′ end AT-rich sequence slightly reduced the binding, indicating that CodYDSM could recognize both palindromic sequences. The proposed binding sequences were found in the promoters of multiple genes involved in amino acids biosynthesis, metabolism, regulation, and stress responses in Synergistetes. Thus, a CodY-like protein from Synergistetes may function similarly to Firmicutes CodY.
1. Introduction
Synergistetes was formally recognized as a novel bacterial phylum based on the 16S rRNA gene phylogeny tree of the bacterial domain in 2009 [1]. The members of the phylum Synergistetes are characterized as Gram-negative, anaerobic bacteria with various cell shapes [1,2]. Microbes of the phylum Synergistetes can utilize amino acids as an energy source and degrade toxic environmental compounds, sludges, and glucose for the production of renewable energy and biogas [1,3,4]. For example, the representative species Synergistes jonesii, isolated from goat rumen, can degrade 3-hydroxy-4(1H)-pyridone (3,4-DHP) for growth [5]. Currently, 16 genera are classified in the phylum Synergistetes: Acetomicrobium, Aminiphilus, Aminivibrio, Aminobacterium, Aminomonas, Cloacibacillus, Dethiosulfovibrio, Fretibacterium, Jonquetella, Lactivibrio, Pacaella, Pyramidobacter, Rarimicrobium, Synergistes, Thermanaerovibrio and Thermovirga [2,6,7,8,9,10]. Microbes of the phylum Synergistetes reside in extended anaerobic environments, including animal and insect gastrointestinal tracts, animal wastes, wastewater treatment systems, soils, and oil wells. Moreover, several species of Synergistetes are present in the oral microbiome and are thought to be associated with periodontal diseases [11,12].
The genome sequences of 35 cultivable strains of Synergistetes, 286,997 pieces of metagenomes from the human gut microbiome, and 2870 pieces of metagenomes from various environmental microbiomes, including the metagenomes of samples from activated sludge, anaerobic digesters, and groundwater are currently available in the NCBI Entrez genomes database [13,14,15,16]. The availability of these sequences provides opportunities for the identification of global regulators that are present in bacterial groups at different taxonomic levels using bioinformatic analyses. Genomic analyses indicate that the genomes of Synergistetes are tightly arranged, with an average of approximately 1000 genes per megabase. While a few species are asaccharolytic, all members of the phylum Synergistetes are able to ferment amino acids [2]. The average percentage of genes encoding proteins associated with amino acid uptake and metabolism in a Synergistetes genome is 11.8 ± 0.8%, representing the highest proportion among all bacterial phyla to date [17].
CodY was first discovered in Bacillus subtilis in 1995 as a repressor of the dipeptide permease operon (dppABCDE) [18]. Later studies confirmed that CodY is a master regulator for various metabolic genes, and that regulation by CodY facilitates the adaptation of bacteria to poor nutritional conditions [19]. Subsequent studies confirmed that CodY is widely distributed and highly conserved among low G + C, Gram-positive bacteria, including pathogens such as Streptococcus pneumoniae and Listeria monocytogenes [20,21,22], in which CodY also regulates the expression of multiple virulence genes [23]. CodY is composed of an N-terminal GAF (cGMP-stimulated phosphodiesterase, adenylate cyclase and a bacterial transcription regulator FhlA) domain and a C-terminal winged helix-turn-helix (wHTH) DNA-binding domain. The GAF domain responds to stimuli and binds with branched-chain amino acids (BCAA, e.g., isoleucine, leucine, and valine) or GTP to activate the DNA-binding activity of CodY [24,25]. Upon activation, the wHTH domain of CodY binds to a palindromic DNA sequence (5′-AATTTTCWGAAAATT) to repress the expression of the downstream gene [20,26,27]. Although GAF and wHTH domains have been described in many proteins, CodY is the only known protein that harbors both a GAF domain and a wHTH domain, and, thus, it is possible to search for putative CodY in bacteria of all phyla using this characteristic, including phyla that are evolutionarily distant from Firmicutes.
In this study, a GAF-wHTH containing, CodY-like protein and its binding consensus sequence were identified in species of the phylum Synergistetes. Additionally, the interaction between the CodY-like protein and the predicted binding sequence was demonstrated using a DNA-affinity precipitation assay (DAPA), indicating that microbes of the Synergistetes phylum likely harbor a CodY-like global regulatory protein for regulation of multiple cellular activities.
2. Results
2.1. Identification and Characterization of CodY-like Proteins in the Phylum Synergistetes
As CodY is the only known protein composed of a GAF and a wHTH domain, profile hidden Markov models (HMMs) were used to investigate whether a CodY-like protein, i.e., proteins with the same domain composition, existed outside of the phylum Firmicutes. An interrogation of bacterial genome sequences in the NCBI Reference Sequence Database (RefSeq), ecological environment metagenomes in the NCBI Entrez genomes database, and human gut microbiome datasets in the European Bioinformatics Institute database (EBI), unexpectedly revealed 107 loci encoding proteins containing both domains in species of the phylum Synergistetes. Among these, 35 loci were present in the genomes of 20 cultivable species, across 12 genera (Table 1), and 23 loci were found in the human gut microbiome datasets across four genera (Table S1). The remaining 49 loci were present in different contings obtained from ecological samples of extreme environments, such as wastewater treatment lagoons, anaerobic digester sludge, activated sludge enrichment, groundwater enrichment, hot springs enrichment, sulfur mats, and the deep marine trench. Thus, a CodY-like protein is widely distributed in the phylum Synergistetes, including species occupying different ecological niches.
To analyze the evolutionary relationship of the newly identified CodY-like protein and CodY of Firmicutes, we first analyzed the phylogenic relationship of all CodY proteins identified in Firmicutes (data not shown), and randomly selected one locus from each evolution branch, resulting in a total of 17 loci. The CodY-like proteins of the 35 cultivable isolates of the phylum Synergistetes were then compared with the CodY proteins of the 17 representative species of the phylum Firmicutes (Table S2) at the deduced amino acid sequence level (Figure 1). Protein identity cluster analysis revealed two main groups: one containing the CodY proteins of Firmicutes, and the other containing the CodY-like proteins of Synergistetes. Thus, CodY and the CodY-like protein of species in the same phylum are more homologous to each other and the extent of similarity decreased across phyla. The putative CodY protein of the phylum Synergistetes displays an average of 43.32% identity (35.32% to 50.20%) with the CodY protein of Firmicutes, while an average of 70.23% and 55.48% identity are observed between species of the phyla Synergistetes and Firmicutes, respectively. The high identity between species of Synergistetes (70.23%), compared to that of Firmicutes (55.48%), is likely due to the relatively low number of species in Synergistetes.
2.2. The Origin of the CodY-like Protein of the Phylum Synergistetes
To examine whether the phylogenetic relationship of CodY and the CodY-like protein follow the same evolutionary history of the diversification of bacterial species, we constructed a 16S rRNA gene-based phylogenetic tree with 51 members of 7 phyla, including first, 5 phyla that do not possess CodY (Table S3): Proteobacteria, Fusobacteria, Bacteroidetes, Actinobacteria and Aquificae; second, the CodY-possessing Firmicutes; and third, Synergistetes. As shown in Figure 2A, species of the phyla Synergistetes and Firmicutes are located in two independent clades. An identical evolutionary relationship was found with regards to the CodY and CodY-like protein (Figure 2B), where the two proteins are also located in two independent branches. Furthermore, in both phylogenic trees, three major groups were observed under the Synergistetes clade, indicating that the evolution of the CodY-like protein follows the evolution of the bacterial species.
2.3. Analysis of the GAF-wHTH Domain
Although only approximately 40% identity was detected between CodY and the CodY-like protein, it is interesting to note that the amino acid sequence of the wHTH domain region of these proteins is relatively conserved across phyla, with an average 73.40% identity (Table S4). All conserved amino acids within the key DNA-binding region (amino acids 203–222) of CodY [28] are also present in the CodY-like proteins, except for Ile212, which is a Val residue in the CodY-like proteins. The conservation of this region suggests that the CodY of Firmicutes and the CodY-like protein of Synergistetes could recognize a similar DNA sequence. On the other hand, the GAF domains of CodY and the CodY-like protein only share an average of 33.41% identity. Nevertheless, the conserved Arg8, Gln15, Glu144 and Thr148 that are essential for the dimerization of CodY in B. subtilis [29] are all present in the corresponding position in the CodY-like protein of Synergistetes, suggesting that the CodY-like protein also acts as a dimer. Although the precise binding motif of BCAA to the GAF domain of the Firmicutes CodY is not well-characterized, studies have shown that amino acid residues Phe40, Arg61, Met62, Met65, Phe71, Pro72, Tyr75, Phe98 and Pro99 are essential for BCAA binding in B. subtilis [25,30]. However, the amino acid residues found in the corresponding positions of the CodY-like proteins were dissimilar to this set of essential residues. Furthermore, the key residues Phe40 and Phe71 are absent in all CodY-like proteins, whereas Phe98 is present only in the CodY-like proteins found in the genera Jonquetella, Pyramidobacter, Dethiosulfovibrio, Aminobacterium, Aminivibrio and Fretibacterium. Thus, the effectors for activation of the DNA-binding activity of the CodY-like protein of Synergistetes may differ from that of the CodY from Firmicutes.
2.4. The Binding Consensus of the CodY-like Protein
The strategy to define the binding consensus of the CodY-like protein was made based on two observations. Firstly, the similarity in the wHTH domain between CodY and the CodY-like protein would suggest that these two proteins recognize similar sequence. Secondly, the expression of codY is autoregulated in microbes of Firmicutes. Specifically, the CodY box is present in the promoters of codY of species in the genera of Lactococcus, Streptococcus, Enterococcus, Staphylococcus and Clostridium [20]. Furthermore, the autoregulation by CodY has been demonstrated experimentally in Lactococcus lactis [26], S. pneumoniae [21], and Streptococcus pyogenes [31]. Thus, it is hypothesized that the expression of the putative codY is autoregulated in Synergistetes and it is feasible to identify the binding consensus of the CodY-like protein in the promoters of the putative codY. Based on the distance between the putative codY and its 5′ flanking loci in the genomes of Synergistetes and results from operon analysis by operon-mapper [32], the putative codY is likely to be a part of an operon, with xerC being the first gene of the operon. Thus, an unbiased search for the canonical CodY box was performed in the promoter regions of the putative codY operon of 35 cultivable strains in the phylum Synergistetes. A sequence of 5′-AATTTTCTTAAAATT, which is identical to the CodY box of Firmicutes, was found in the promoter regions of 12 isolates of the genera Synergistes and Cloacibacillus, but not in other genera. To verify whether additional consensus sequences could be recognized by the CodY-like protein, we extracted the 5′-flanking regions of xerC from genomes of cultivatable isolates, human gut metagenome-assembled genomes, and ecological metagenome assembly sequences in the phylum Synergistetes, and used the MEME motif-searching algorithm [33] to search for conserved palindromic sequences in each genus. As shown in Table S5 and Figure 3, four consensus sequences were discovered in the genera Synergistes/Cloacibacillus, Thermanaerovibrio, Pyramidobacter, and Acetomicrobium, respectively. No consensus motifs were discovered in other genera because less than three genomes were available for a specific genus. The consensus sequence identified in the genera Synergistes and Cloacibacillus, 5′-AATTTTCTTAAAATTTCSCTTGATATTTACAATTTT, contains a Firmicutes CodY box (underlined) and an AAAATT/AATTTT inverted repeat (double underlined). Upon close examination of the consensus found in the genera Thermanaerovibrio (5′-AATATT-N21-AAAATA), Pyramidobacter (5′-ATTTTY-N14-AAAAAT), and Acetomicrobium (5′-WWTTTT-N12-ATAATT), it was noted that all of the sequences contained a pair of AT-rich palindromic arms separated by a spacer region of variable length and sequence, suggesting that the binding sequence of the CodY-like protein of the phylum Synergistetes is a palindromic sequence consisting of two AT-rich arms and a variable spacer region.
2.5. The CodY-like Protein of Cloacibacillus Evryensis DSM19522 Could Interact with Both the Canonical CodY Box and an AAAATT-N15-AATTTT Palindromic Sequence
As the consensus sequence identified in the genera Synergistes and Cloacibacillus contains both a Firmicutes CodY box and an extended palindromic sequence, the binding specificity of the CodY-like protein of C. evryensis DSM19522 (CodYDSM) was analyzed using recombinant histidine-tagged CodYDSM (His-CodYDSM) in DAPA experiments, as detailed in Materials and Methods. His-CodYDSM was readily induced and purified from the recombinant E. coli strain (Figure S1), but the solubility of this protein was rather low (0.2–0.4 μg mL−1). The predicted binding sequence at the 5′ flanking region of xerC (CLOEV_RS08635) was extracted from the genome of C. evryensis DSM19522 (NZ_KK073872.1) and used to design probes for the DAPA experiment (Table S6). The DAPA results indicated that His-CodYDSM could interact specifically with the full-length probe in a dose-dependent manner (Figure 4). When the three AT-rich sequence were substituted with GGGGGG or six random bases, the signals were abolished, indicating that AT-rich palindromic sequences are critical for the binding of the CodY-like protein. While substitutions of the 5′ end AATTTT with CCCCCC reduced marginally the binding of His-CodYDSM, mutations in the 3′ AATTTT did not affect the binding of His-CodYDSM significantly, indicating that His-CodYDSM could interact with both the canonical CodY box and an AAAATT-N15-AATTTT inverted repeat sequence, and possibly with a slightly higher affinity to the canonical CodY box. On the other hand, mutations of the center AAAATT significantly reduced the binding, confirming that the CodY-like protein could interact with both targets. Thus, the obtained consensus sequence contains two CodYDSM binding sites; one of which is identical to the CodY box of Firmicutes, and the other is AATTTT-N15-AAAATT.
2.6. Identification and Functional Prediction of the CodY-like Regulon in the Phylum Synergistetes
As the first step to investigate whether the CodY-like protein also acts as a global regulator in Synergistetes, a search for the proposed binding sequence of the CodY-like protein was performed in the intergenic regions of the 21 complete genomes of Synergistetes, which includes genera Synergistetes, Cloacibacillus, Pyramidobacter and Acetomicrobium. The genus Thermanaerovibrio was not included as only two complete genomes are available (Table 1). To be noted, since the DAPA analysis revealed that His-CodYDSM could interact with both the CodY box of Firmicutes (5′-AATTTTTCWGAAAATT) and a palindromic sequence with an extended spacer region (Figure 3 and Table S5), both motifs were used to search for potential targets in the corresponding genus. It was found that the consensus sequence was widely distributed in the intergenic regions of genomes analyzed (Table S7), accounting for approximately 8–13% of the genes in a genome (Figure 5 and Table S8). Although the percentages were comparable between the four genera, genera Synergistetes and Cloacibacillus showed higher numbers of loci that harbor both motifs compared to those of genera Acetomicrobium and Pyramidobacter. The wide distribution of this novel consensus sequence in a genome suggests that the CodY-like protein is a global regulator. Among these loci, homologs of 32 loci are parts of the CodY regulon in Firmicutes (Table S9). These loci include genes encoding proteins involved in BCAA biosynthesis (ilvA, ilvN, ilvB, ilvD, ilvC, leuC, WP_051682752.1, WP_200778737.1 and WP_037975652.1), BCAA uptake (WP_037976683.1, WP_037975074.1, WP_037976889.1 and WP_009202431.1), proteolysis (clpB, clpX, clpP, fstH and resP), peptide degradation (WP_037974402.1, pepV, def, WP_051682620.1, WP_051682707.1, WP_078015053.1, WP_037973903.1, WP_111089616.1, WP_037974668.1 and WP_008708838.1), and peptide and amino acid transport (WP_009201625.1, WP_078015996.1, WP_008709275.1 and WP_120373146.1). This observation supports the hypothesis that the CodY-like global regulator of Synergistetes functions in parallel to Firmicutes CodY.
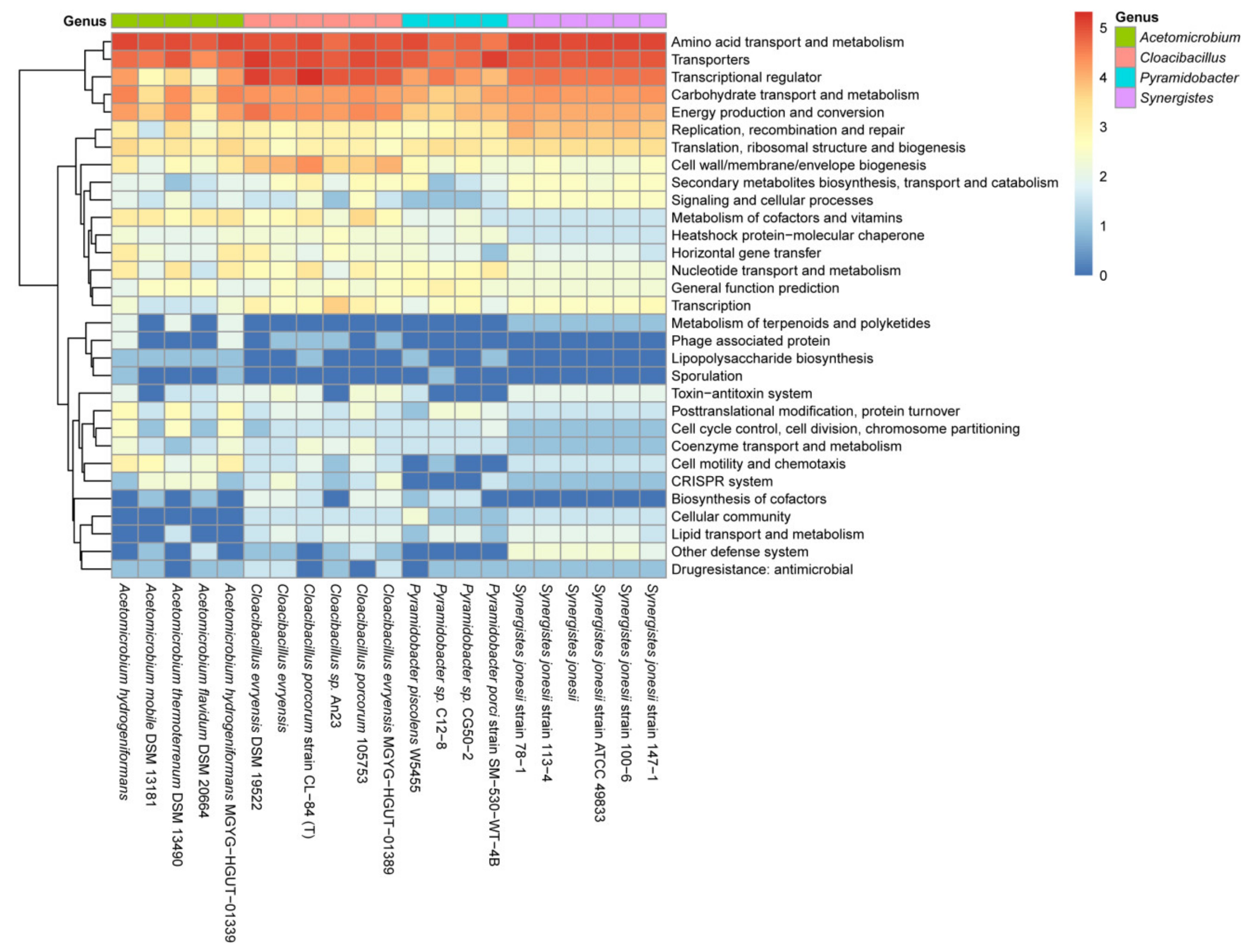
To further verify this hypothesis, we computed the functional enrichment of groups of genes listed in Table S7 in the genera Acetomicrobium, Cloacibacillus, Pyramidobacter, and Synergistes (Figure 6). Three main clusters emerged, with proteins in the categories of amino acid transport and metabolism, transporters, transcriptional regulators, carbohydrate transport and metabolism, and energy production and conversion being the most conserved in the four genera. The identification of genes encoding transcriptional regulators, including HrcA and regulators of the P-II family, GntR family, and CtsR family (Table S7), suggests that the regulon of the CodY-like protein would contain a set of genes indirectly regulated by the CodY-like protein. Specifically, regulators of both the PII family and the GntR family are closely associated with the regulation of nitrogen metabolism and the nitrogen-carbon interconversion in both Gram-positive and Gram-negative bacteria. HrcA, a heat-inducible transcription repressor, and CtsR, a transcriptional regulator of class III stress genes [34,35], could be involved in the stress responses. Additionally, transcriptional regulators of the LacI, LysR, TetR/AcrR, MurR/RpiR, MerR and MarR families were also found. These regulators are implicated in the regulation of many cellular processes, including metabolism, metal ion homeostasis, antibiotics resistance, virulence expression and stress responses, and thus, a regulon governed by the CodY-like protein is likely to indirectly control multiple cellular activities in Synergistetes.
Although the genes in the second cluster were generally less conserved, compared to the genes described above between the four genera, significant conservation within a genus was detected in genes encoding proteins involved in genetic information storage and processing, including DNA replication, recombination and repair, transcription, translation, ribosomal structure and biogenesis, nucleotide transport and metabolism. Genes encoding proteins involved in the biosynthesis of cell envelopes, secondary metabolites biosynthesis, metabolism of cofactors and vitamins, heat-shock proteins and molecular chaperones, and horizontal gene transfer were also present in the second cluster (Figure 6). Loci that were least conserved between genera were genes involved in the metabolism of terpenoids and polyketides, phage production, post-translational modification and protein turnover, the toxin-antitoxin system, cell motility and chemotaxis, CRISPR and other defense systems, and cellular community. Nevertheless, conservation among species within a genus was evident. For instance, loci associated with post-translational modification, protein turnover, cell cycle control, cell division, chromosome partition, coenzyme transport and metabolism, cell motility and chemotaxis, and the CRISPR system were enriched in the genus Acetomicrobium, and loci encoding proteins of the toxin-antitoxin systems, lipid transport and metabolism, and other defense systems were enriched in the genus Synergistes. Taken together, the loci involved in nutrient uptake, general metabolism and regulation were most conserved between the four genera, whereas loci involved in defense systems and stress responses were less conserved. The variations in the putative regulons of the CodY-like regulatory protein between genera may be associated with the natural niches of these microbes.
3. Discussion
CodY has been extensively studied in the Firmicutes for its dominant role in metabolism, stress responses and pathogenesis [23,36,37,38]. This study confirmed the presence of CodY orthologues in Synergistetes, which generally exhibits a G + C content close to, or greater than, 50% [39,40,41,42,43]. Thus, the distribution of CodY and the CodY-like protein in bacteria may be more widespread than previously known, and regulation by CodY and its homologs may have similar impacts on bacteria occupying different niches. The differences in the G + C content, the ecological niches and metabolic activities between Firmicutes and Synergistetes would suggest that CodY and the CodY-like protein evolved divergently. As the evolutionary relationship of these proteins follows the phylogenetic relationship of the genera in phyla Firmicutes and Synergistetes, genes encoding CodY and the CodY-like protein should originate from a common ancestor. The results of this study indicate that these two proteins are orthologues that are conserved in domain structures and functions, but distinct in their overall amino acid sequence.
Past transcriptomic studies of the CodY regulon in Firmicutes, either by microarray or RNA sequencing, often identified loci without a canonical CodY box in the promoter region. For instance, approximately 60% and 30% of the identified CodY targets in B. subtilis and L. lactis, respectively, are without a canonical CodY box [20]. Although some of these loci are regulated by transcriptional regulators whose activity is governed by CodY, CodY likely directly regulates the rest of the targets through binding to a modified consensus motif. This hypothesis is supported by more recent studies in B. subtilis [44], L. monocytogenes [22], and Staphylococcus aureus [45], in which a modified CodY-binding consensus generated from two overlapping canonical CodY boxes (5′-AATTTTCWGAAWWTTCWGAAAATT) was reported. Unlike the CodY box described in Firmicutes, the binding consensus in Synergistetes is more like a palindromic sequence with AT-rich arms separated by an extended spacer region, as neither the lengths nor the sequences of the identified consensus sequences fit an overlapping arrangement. Although it is possible that the targets identified in this study do not cover the full regulon, the presence of the binding consensus at the 5′ region of multiple genes strongly suggests that similar to CodY in Firmicutes, this newly identified GAF-wHTH protein is a global regulatory protein in Synergistetes.
As a system for genetic manipulation has yet to be established in microbes of the phylum Synergistetes, we selected the DAPA analysis to analyze the DNA-binding activity of the CodY-like protein. Attempts were also made to use gel electrophoresis mobility shift assays (EMSA) [46] to investigate the binding specificity of the CodY-like protein; however, the solubilities of the recombinant CodY-like proteins of Synergistetes were generally lower than those required for effective analysis by EMSA with biotin-labeled probes. Using Protein-Sol (https://protein-sol.manchester.ac.uk/, accessed on 14 January 2022), we found that the solubility of the recombinant CodY-like protein of Synergistetes was significantly lower than CodY from Streptococcus salivarius 57.I [47]. Proteins with a Protein-Sol calculated value above 0.45 are considered highly soluble [48]; and, while the Protein-Sol value of CodY from S. salivarius 57.I is 0.719, and the value for the CodY-like protein in C. evryensis DSM19522 is 0.418. Similar results were also found in CodY-like proteins of two randomly selected species: Thermanaerovibrio acidaminovorans DSM 6589, with a value of 0.455, and C. porcorum CL-84, with a value of 0.426. By using Ni-NTA magnetic beads, we were able to perform DAPA experiments in a relatively large reaction volume, compared to EMSA analysis, which circumvented any limitations of the low solubility of the CodY-like protein.
Although the difference in the GAF domain between CodY and the CodY-like protein could result in a difference in the effectors, both proteins are involved in the regulation of BCAA biosynthesis. Thus, BCAA is also likely to be the effector of the CodY-like protein. On the other hand, while both BCAA and GTP are necessary to activate CodY in B. subtilis [49], CodY of L. lactis [50], Streptococcus mutans [51], and S. pneumoniae [21] is not activated by GTP, suggesting that the choice and efficiency of effector molecules to activate CodY activity could be affected by the ecological niche and physiologic activity of the bacteria. As microbes of Synergistetes could metabolize a variety of substrates, including DHP [5], plant toxin fluoroacetate [52], and mucin, it is also possible that microbes of Synergistetes could utilize different metabolites to signal intracellular nutrient status, and perhaps to activate the CodY-like protein.
As members of phylum Synergistetes inhabit a wide range of extreme environments, it is expected that they have evolved sophisticated features to cope with environmental stresses, including extreme temperature, pH, toxic compounds and challenges from other microbes or bacteriophages. As loci in motility/chemotaxis, defense mechanisms and CRISPR systems all are less conserved among the regulons governed by the CodY-like proteins in Synergistetes, it would suggest that the regulons could vary according to the natural niches of all species in the phylum. For instance, based on our analysis, the CodY-like protein could potentially regulate the chemotaxis and motility of flagella-bearing Acetomicrobium thermoterrenum strain DSM 13490 by regulating the expression of loci encoding the methyl-accepting chemotaxis protein (Mcp) and proteins involved in flagella biosynthesis Thus, regulation by the CodY-like protein could potentially fine-tune the activity of chemotaxis and swimming activity. Such regulation seems necessary for bacteria that reside in constantly changing environments, and the regulation could provide certain advantages for these bacteria to thrive in harsh environments.
Taken together, although Synergistetes and Firmicutes are phylogenetically distinct, both CodY and the CodY-like protein are conserved with respect to their regulatory function. Based on the potential targets of the CodY-like protein, the regulation spectrum of the CodY-like protein overlaps with that of the Firmicutes CodY regulon. Thus, similar to Firmicutes, regulation by the CodY-like protein is likely to be essential for the microbes of Synergistetes to cope with stresses in their natural niches.
4. Materials and Methods
4.1. Bacterial Genome and Metagenome Database
The cultivable bacterial genomes were obtained from the NCBI Genbank database of sequenced bacteria (last updated August 2020) (http://ftp.ncbi.nlm.nih.gov/refseq/release/, accessed on 24 August 2020). Metagenomic data used in this study included genomes obtained from samples from various ecological sources (https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?mode=Undef&id=408169&lvl=3&keep=1&srchmode=1&unlock/, accessed on 26 Octomber 2020), and human gut metagenomic data (http://ftp.ebi.ac.uk/pub/databases/metagenomics/mgnify_genomes/human-gut/v1.0/uhgp_catalogue/uhgp-100.tar.gz/, accessed on 6 November 2020).
4.2. Identification of CodY Homologous Proteins
HMMER, version 3.1b2 [53], was used to search the GAF (PF06018) and HTH (PF08222) domains of CodY in the deduced amino acid sequence derived from the bacterial genome and metagenome databases. The significance was set at an e-value < 10−3. A putative CodY was identified in proteins that contained both the GAF and HTH domains. BlastP analysis was used to calculate the average amino acid identity of the CodY-like proteins of Synergistetes (Table 1) and the CodY proteins of Firmicutes (Table S2).
4.3. Phylogenetic Analysis of 16S rRNA and the CodY-like/CodY Proteins from Synergistetes and Other Phyla
The 16S rRNA genes from 35 strains of the phylum Synergistetes (Table 1), 17 species of the Firmicutes (Table S2) and 12 species from the phyla Proteobacteria, Fusobacteria, Bacteroidetes, Actinobacteria and Aquificae (Table S3) were retrieved from the database. An alignment of these 16S genes was performed using ClustalW integrated in MEGA version 7 [54] with standard parameters. The sequences of the codY genes from species of Synergistetes and Firmicutes were retrieved from the database, and an alignment of the deduced protein sequence was prepared using MUSCLE [55] integrated into MEGA version 7 with standard parameters. The maximum likelihood algorithm (ML) with 1000 bootstrap replications was used for phylogenetic tree reconstruction. The phylogenetic tree was managed and visualized by iTOL, version 5 [56].
4.4. Purification of the Recombinant CodY-like Protein
A preparation of the total cellular DNA of C. evryensis DSM19522 was purchased from the Leibniz Institute DSMZ (Braunschweig, Germany). The coding region of the putative codY (CLOEV_RS08630) was PCR amplified from chromosomal DNA by using the Blend Taq™-Plus (Osaka, Toyobo) with primers DSM19522_codY-SphI_S and DSM19522_codY-HindIII_AS. The estimated melting temperatures of primers DSM19522_codY-SphI_S and DSM19522_codY-HindIII_AS are 60.7 °C and 61.4 °C, respectively. The annealing temperature of the PCR was set at 56 °C (30 s), and reaction was performed for 35 cycles. Restriction recognition sequences were included in the primers to facilitate cloning. The PCR product was restriction digested and cloned into pQE30 (Qiagen, Hilden, Germany) in E. coli M15. The identity of the recombinant plasmid was verified by sequencing analysis. The recombinant histidine-tagged CodY-like protein (His-CodYDSM) was induced and purified from the recombinant E. coli strain by HIS-Select Nickel Affinity Gel (Sigma, St. Louis, MO, USA) under native conditions, according to the manufacturer’s recommendation. The concentration of the purified protein was determined by Bio-Rad Protein Assay based on the method of Bradford [57]. All primers used in this study are listed in Table S6.
4.5. DAPA
All oligonucleotides used in DAPA were end-labeled using the Pierce biotin 3′ end DNA labeling kit (Thermo, Waltham, MA, USA). Two complementary biotin-labeled oligonucleotides containing the desired sequences were annealed and then used in DAPA reactions. Various amounts of purified His-CodYDMS was mixed with 10 μL Mag-Beads- Ni-NTA (Tools, Taiwan) in separate reactions, with constant rotation for 30 min at room temperature. At the end of the reaction, the beads were washed with 1 mL reaction buffer (10 mM MgCl2, 50 mM KCl, 10 mM Tris, [pH 7.5], 1 mM EDTA, 50 µg ml−1 BSA) in a magnetic field to remove unbound proteins. The protein-DNA binding reaction was carried out in 50 μL of the reaction buffer containing 0.1 pmol biotin-labeled probe and poly[dI-dC] at 20 µg mL−1, and the reaction was incubated with tilt rotation for 30 min at room temperature. The final concentrations of His-CodYDMS in the reactions were 0.02, 0.1, 0.5 and 2.5 μM, respectively. At the end of incubation, the beads were washed with 200 μL of reaction buffer three times, and then resuspended in 20 µL TE buffer (50 mM Tris, 10 mM EDTA [pH 8]) containing proteinase K at 2 mg mL−1. The digestion was carried out at 37 °C for 30 min to release the DNA. The final product was resolved on 6% non-denaturing polyacrylamide gels, electro-transferred onto nylon membranes, and detected using a chemiluminescent nucleic acid detection module kit (Pierce). The DAPA experiments were repeated six times with three batches of purified His-CodYDMS.
4.6. Binding Site Analysis and Target Gene Prediction
The sequence of a 400-bp region 5′ to the xerC gene, the first gene of the codY-containing operon of species of the phylum Synergistetes were extracted from the genomes of 35 cultivatable isolates, 17 human gut metagenome-assembled genomes and 22 ecological metagenome assembly sequences. All these sequences were then used for common motif discovery using MEME motif-searching algorithm [58]. Genera with less than three genomes available were excluded from the analysis. The default settings were used with the following exceptions: the minimum length was set to 15 bases and interrogated sequences with palindromes only.
To define the possible regulon of the CodY-like protein, all promoter regions of the 21 complete genomes in the phylum Synergistetes were scanned for 5′-AATTTTCWGAAAATT and the newly identified consensus binding sequence of the CodY-like protein by FIMO [59].
4.7. Functional Enrichment of the Identified Target Genes Regulated by the CodY-like Protein
The functional clustering of the target genes regulated by the CodY-like proteins was initially analyzed using Roary [60]. The identified target genes were further enriched based on predicted functions using EggNog [61], Clusters of Orthologous Groups (COGs) [62] and KEGG orthology [63]. The enrichment score was calculated using log2(N+1) (where N is the number of the identified genes). R script was used to generate the heat map showing the enrichment scores.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms23147911/s1.
Author Contributions
Conceptualization, J.G., S.H. and Y.-Y.M.C.; methodology, S.L., H.-R.S. and H.-Y.W.; software, J.G. and S.L.; validation, S.H. and Y.-Y.M.C.; formal analysis, J.G. and S.L.; investigation, J.G, H.-R.S. and H.-Y.W.; resources, S.H.; data curation, J.G., S.L. and Y.-Y.M.C.; writing—original draft preparation, J.G., S.L. and Y.-Y.M.C.; writing—review and editing, S.H. and Y.-Y.M.C.; visualization, J.G. and S.L.; supervision, S.H. and Y.-Y.M.C.; project administration, S.H.; funding acquisition, J.G., S.H. and Y.-Y.M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by the National Key Research and Development Program of China (2021YFA0909500 to SH), the National Natural Science Foundation of China (31200968 to JG) and the Ministry of Science and Technology (MOST) of Taiwan, (108-2320-B-182-020-YM3 to YMC).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Y. Luo, S. Gao and X. Ge for technical assistance and discussion of the results. We thank R. Faustoferri for review of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jumas-Bilak, E.; Roudiere, L.; Marchandin, H. Description of ‘Synergistetes’ phyl. nov. and emended description of the phylum ‘Deferribacteres’ and of the family Syntrophomonadaceae, phylum ‘Firmicutes’. Int. J. Syst. Evol. Microbiol. 2009, 59, 1028–1035. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, V.; Gupta, R.S. Molecular signatures for the phylum Synergistetes and some of its subclades. Antonie Van Leeuwenhoek 2012, 102, 517–540. [Google Scholar] [CrossRef] [PubMed]
- Maune, M.W.; Tanner, R.S. Description of Anaerobaculum hydrogeniformans sp. nov., an anaerobe that produces hydrogen from glucose, and emended description of the genus Anaerobaculum. Int. J. Syst. Evol. Microbiol. 2012, 62, 832–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahle, H.; Birkeland, N.K. Thermovirga lienii gen. nov., sp. nov., a novel moderately thermophilic, anaerobic, amino-acid-degrading bacterium isolated from a North Sea oil well. Int. J. Syst. Evol. Microbiol. 2006, 56, 1539–1545. [Google Scholar] [CrossRef] [Green Version]
- Allison, M.J.; Mayberry, W.R.; McSweeney, C.S.; Stahl, D.A. Synergistes jonesii, gen. nov., sp.nov.: A rumen bacterium that degrades toxic pyridinediols. Syst. Appl. Microbiol. 1992, 15, 522–529. [Google Scholar] [CrossRef]
- Honda, T.; Fujita, T.; Tonouchi, A. Aminivibrio pyruvatiphilus gen. nov., sp. nov., an anaerobic, amino-acid-degrading bacterium from soil of a Japanese rice field. Int. J. Syst. Evol. Microbiol. 2013, 63, 3679–3686. [Google Scholar] [CrossRef]
- Jumas-Bilak, E.; Bouvet, P.; Allen-Vercoe, E.; Aujoulat, F.; Lawson, P.A.; Jean-Pierre, H.; Marchandin, H. Rarimicrobium hominis gen. nov., sp. nov., representing the fifth genus in the phylum Synergistetes that includes human clinical isolates. Int. J. Syst. Evol. Microbiol. 2015, 65, 3965–3970. [Google Scholar] [CrossRef]
- Ndongo, S.; Dubourg, G.; Fournier, P.E.; Raoult, D. Pacaella massiliensis gen. nov., sp. nov., a new bacterial species isolated from the human gut. New Microbes New Infect. 2017, 16, 32–33. [Google Scholar] [CrossRef]
- Qiu, Y.L.; Hanada, S.; Kamagata, Y.; Guo, R.B.; Sekiguchi, Y. Lactivibrio alcoholicus gen. nov., sp. nov., an anaerobic, mesophilic, lactate-, alcohol-, carbohydrate- and amino-acid-degrading bacterium in the phylum Synergistetes. Int. J. Syst. Evol. Microbiol. 2014, 64, 2137–2145. [Google Scholar] [CrossRef]
- Vartoukian, S.R.; Downes, J.; Palmer, R.M.; Wade, W.G. Fretibacterium fastidiosum gen. nov., sp. nov., isolated from the human oral cavity. Int. J. Syst. Evol. Microbiol. 2013, 63, 458–463. [Google Scholar] [CrossRef] [PubMed]
- McCracken, B.A.; Nathalia Garcia, M. Phylum Synergistetes in the oral cavity: A possible contributor to periodontal disease. Anaerobe 2021, 68, 102250. [Google Scholar] [CrossRef] [PubMed]
- Wade, W.G. The oral microbiome in health and disease. Pharmacol. Res. 2013, 69, 137–143. [Google Scholar] [CrossRef]
- Chovatia, M.; Sikorski, J.; Schroder, M.; Lapidus, A.; Nolan, M.; Tice, H.; Glavina Del Rio, T.; Copeland, A.; Cheng, J.F.; Lucas, S.; et al. Complete genome sequence of Thermanaerovibrio acidaminovorans type strain (Su883). Stand. Genom. Sci. 2009, 1, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, C.K.; Webb, R.I.; Sly, L.I.; Denman, S.E.; McSweeney, C.S. Isolation and survey of novel fluoroacetate-degrading bacteria belonging to the phylum Synergistetes. FEMS Microbiol. Ecol. 2012, 80, 671–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; McVeigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef] [PubMed]
- Plichta, D.R.; Juncker, A.S.; Bertalan, M.; Rettedal, E.; Gautier, L.; Varela, E.; Manichanh, C.; Fouqueray, C.; Levenez, F.; Nielsen, T.; et al. Transcriptional interactions suggest niche segregation among microorganisms in the human gut. Nat. Microbiol. 2016, 1, 16152. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Hooper, S.D.; Kyrpides, N.C. Focus: Synergistetes. Environ. Microbiol. 2009, 11, 1327–1329. [Google Scholar] [CrossRef]
- Slack, F.J.; Serror, P.; Joyce, E.; Sonenshein, A.L. A gene required for nutritional repression of the Bacillus subtilis dipeptide permease operon. Mol. Microbiol. 1995, 15, 689–702. [Google Scholar] [CrossRef]
- Sonenshein, A.L. Control of key metabolic intersections in Bacillus subtilis. Nat. Rev. Microbiol. 2007, 5, 917–927. [Google Scholar] [CrossRef]
- Guedon, E.; Sperandio, B.; Pons, N.; Ehrlich, S.D.; Renault, P. Overall control of nitrogen metabolism in Lactococcus lactis by CodY, and possible models for CodY regulation in Firmicutes. Microbiology 2005, 151, 3895–3909. [Google Scholar] [CrossRef] [Green Version]
- Hendriksen, W.T.; Bootsma, H.J.; Estevao, S.; Hoogenboezem, T.; de Jong, A.; de Groot, R.; Kuipers, O.P.; Hermans, P.W. CodY of Streptococcus pneumoniae: Link between nutritional gene regulation and colonization. J. Bacteriol. 2008, 190, 590–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, R.; Sonenshein, A.L.; Belitsky, B.R. Genome-wide identification of Listeria monocytogenes CodY-binding sites. Mol. Microbiol. 2020, 113, 841–858. [Google Scholar] [CrossRef] [PubMed]
- Stenz, L.; Francois, P.; Whiteson, K.; Wolz, C.; Linder, P.; Schrenzel, J. The CodY pleiotropic repressor controls virulence in gram-positive pathogens. FEMS Immunol. Med. Microbiol. 2011, 62, 123–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, A.R.; Kang, H.R.; Son, J.; Kwon, D.H.; Kim, S.; Lee, W.C.; Song, H.K.; Song, M.J.; Hwang, K.Y. The structure of the pleiotropic transcription regulator CodY provides insight into its GTP-sensing mechanism. Nucleic Acids Res. 2016, 44, 9483–9493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levdikov, V.M.; Blagova, E.; Colledge, V.L.; Lebedev, A.A.; Williamson, D.C.; Sonenshein, A.L.; Wilkinson, A.J. Structural rearrangement accompanying ligand binding in the GAF domain of CodY from Bacillus subtilis. J. Mol. Biol. 2009, 390, 1007–1018. [Google Scholar] [CrossRef] [Green Version]
- den Hengst, C.D.; van Hijum, S.A.; Geurts, J.M.; Nauta, A.; Kok, J.; Kuipers, O.P. The Lactococcus lactis CodY regulon: Identification of a conserved cis-regulatory element. J. Biol. Chem. 2005, 280, 34332–34342. [Google Scholar] [CrossRef] [Green Version]
- Belitsky, B.R.; Sonenshein, A.L. Genetic and biochemical analysis of CodY-binding sites in Bacillus subtilis. J. Bacteriol. 2008, 190, 1224–1236. [Google Scholar] [CrossRef] [Green Version]
- Joseph, P.; Ratnayake-Lecamwasam, M.; Sonenshein, A.L. A region of Bacillus subtilis CodY protein required for interaction with DNA. J. Bacteriol. 2005, 187, 4127–4139. [Google Scholar] [CrossRef] [Green Version]
- Levdikov, V.M.; Blagova, E.; Joseph, P.; Sonenshein, A.L.; Wilkinson, A.J. The structure of CodY, a GTP- and isoleucine-responsive regulator of stationary phase and virulence in Gram-positive bacteria. J. Biol. Chem. 2006, 281, 11366–11373. [Google Scholar] [CrossRef] [Green Version]
- Villapakkam, A.C.; Handke, L.D.; Belitsky, B.R.; Levdikov, V.M.; Wilkinson, A.J.; Sonenshein, A.L. Genetic and biochemical analysis of the interaction of Bacillus subtilis CodY with branched-chain amino acids. J. Bacteriol. 2009, 191, 6865–6876. [Google Scholar] [CrossRef] [Green Version]
- Kreth, J.; Chen, Z.; Ferretti, J.; Malke, H. Counteractive balancing of transcriptome expression involving CodY and CovRS in Streptococcus pyogenes. J. Bacteriol. 2011, 193, 4153–4165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taboada, B.; Estrada, K.; Ciria, R.; Merino, E. Operon-mapper: A web server for precise operon identification in bacterial and archaeal genomes. Bioinformatics 2018, 34, 4118–4120. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Elsholz, A.K.; Michalik, S.; Zuhlke, D.; Hecker, M.; Gerth, U. CtsR, the Gram-positive master regulator of protein quality control, feels the heat. EMBO J. 2010, 29, 3621–3629. [Google Scholar] [CrossRef] [Green Version]
- Kruger, E.; Hecker, M. The first gene of the Bacillus subtilis clpC operon, ctsR, encodes a negative regulator of its own operon and other class III heat shock genes. J. Bacteriol. 1998, 180, 6681–6688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinsmade, S.R. CodY, a master integrator of metabolism and virulence in Gram-positive bacteria. Curr. Genet. 2017, 63, 417–425. [Google Scholar] [CrossRef]
- Geng, J.; Huang, S.C.; Chen, Y.Y.; Chiu, C.H.; Hu, S.; Chen, Y.M. Impact of growth pH and glucose concentrations on the CodY regulatory network in Streptococcus salivarius. BMC Genom. 2018, 19, 386. [Google Scholar] [CrossRef]
- Richardson, A.R.; Somerville, G.A.; Sonenshein, A.L. Regulating the intersection of metabolism and pathogenesis in Gram-positive bacteria. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Menes, R.J.; Muxi, L. Anaerobaculum mobile sp. nov., a novel anaerobic, moderately thermophilic, peptide-fermenting bacterium that uses crotonate as an electron acceptor, and emended description of the genus Anaerobaculum. Int. J. Syst. Evol. Microbiol. 2002, 52, 157–164. [Google Scholar] [CrossRef]
- Baena, S.; Fardeau, M.L.; Ollivier, B.; Labat, M.; Thomas, P.; Garcia, J.L.; Patel, B.K. Aminomonas paucivorans gen. nov., sp. nov., a mesophilic, anaerobic, amino-acid-utilizing bacterium. Int. J. Syst. Bacteriol. 1999, 49 Pt 3, 975–982. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, Y.; Kamagata, Y.; Nakamura, K.; Ohashi, A.; Harada, H. Syntrophothermus lipocalidus gen. nov., sp. nov., a novel thermophilic, syntrophic, fatty-acid-oxidizing anaerobe which utilizes isobutyrate. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 2, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, C.; Baena, S.; Fardeau, M.L.; Patel, B.K.C. Aminiphilus circumscriptus gen. nov., sp. nov., an anaerobic amino-acid-degrading bacterium from an upflow anaerobic sludge reactor. Int. J. Syst. Evol. Microbiol. 2007, 57, 1914–1918. [Google Scholar] [CrossRef] [PubMed]
- Surkov, A.V.; Dubinina, G.A.; Lysenko, A.M.; Glockner, F.O.; Kuever, J. Dethiosulfovibrio russensis sp. nov., Dethosulfovibrio marinus sp. nov. and Dethosulfovibrio acidaminovorans sp. nov., novel anaerobic, thiosulfate- and sulfur-reducing bacteria isolated from ‘Thiodendron’ sulfur mats in different saline environments. Int. J. Syst. Evol. Microbiol. 2001, 51, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belitsky, B.R.; Sonenshein, A.L. Genome-wide identification of Bacillus subtilis CodY-binding sites at single-nucleotide resolution. Proc. Natl. Acad. Sci. USA 2013, 110, 7026–7031. [Google Scholar] [CrossRef] [Green Version]
- Majerczyk, C.D.; Dunman, P.M.; Luong, T.T.; Lee, C.Y.; Sadykov, M.R.; Somerville, G.A.; Bodi, K.; Sonenshein, A.L. Direct targets of CodY in Staphylococcus aureus. J. Bacteriol. 2010, 192, 2861–2877. [Google Scholar] [CrossRef] [Green Version]
- Hellman, L.M.; Fried, M.G. Electrophoretic mobility shift assay (EMSA) for detecting protein-nucleic acid interactions. Nat. Protoc. 2007, 2, 1849–1861. [Google Scholar] [CrossRef]
- Huang, S.C.; Burne, R.A.; Chen, Y.Y. The pH-dependent expression of the urease operon in Streptococcus salivarius is mediated by CodY. Appl. Environ. Microbiol. 2014, 80, 5386–5393. [Google Scholar] [CrossRef] [Green Version]
- Hebditch, M.; Carballo-Amador, M.A.; Charonis, S.; Curtis, R.; Warwicker, J. Protein-Sol: A web tool for predicting protein solubility from sequence. Bioinformatics 2017, 33, 3098–3100. [Google Scholar] [CrossRef] [Green Version]
- Shivers, R.P.; Sonenshein, A.L. Activation of the Bacillus subtilis global regulator CodY by direct interaction with branched-chain amino acids. Mol. Microbiol. 2004, 53, 599–611. [Google Scholar] [CrossRef]
- den Hengst, C.D.; Curley, P.; Larsen, R.; Buist, G.; Nauta, A.; van Sinderen, D.; Kuipers, O.P.; Kok, J. Probing direct interactions between CodY and the oppD promoter of Lactococcus lactis. J. Bacteriol. 2005, 187, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Lemos, J.A.; Nascimento, M.M.; Lin, V.K.; Abranches, J.; Burne, R.A. Global regulation by (p)ppGpp and CodY in Streptococcus mutans. J. Bacteriol. 2008, 190, 5291–5299. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Khan, S.; Webb, R.; Denman, S.; McSweeney, C. Characterization and survey in cattle of a rumen Pyrimadobacter sp. which degrades the plant toxin fluoroacetate. FEMS Microbiol. Ecol. 2020, 96, fiaa077. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for occurrences of a given motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernandez-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galperin, M.Y.; Wolf, Y.I.; Makarova, K.S.; Vera Alvarez, R.; Landsman, D.; Koonin, E.V. COG database update: Focus on microbial diversity, model organisms, and widespread pathogens. Nucleic Acids Res. 2021, 49, D274–D281. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
A heat map showing the identity between the CodY/CodY-like proteins from species of Firmicutes and Synergistetes. The score is the % identity of the deduced amino acid sequence of codY and its homologs.
Figure 1.
A heat map showing the identity between the CodY/CodY-like proteins from species of Firmicutes and Synergistetes. The score is the % identity of the deduced amino acid sequence of codY and its homologs.
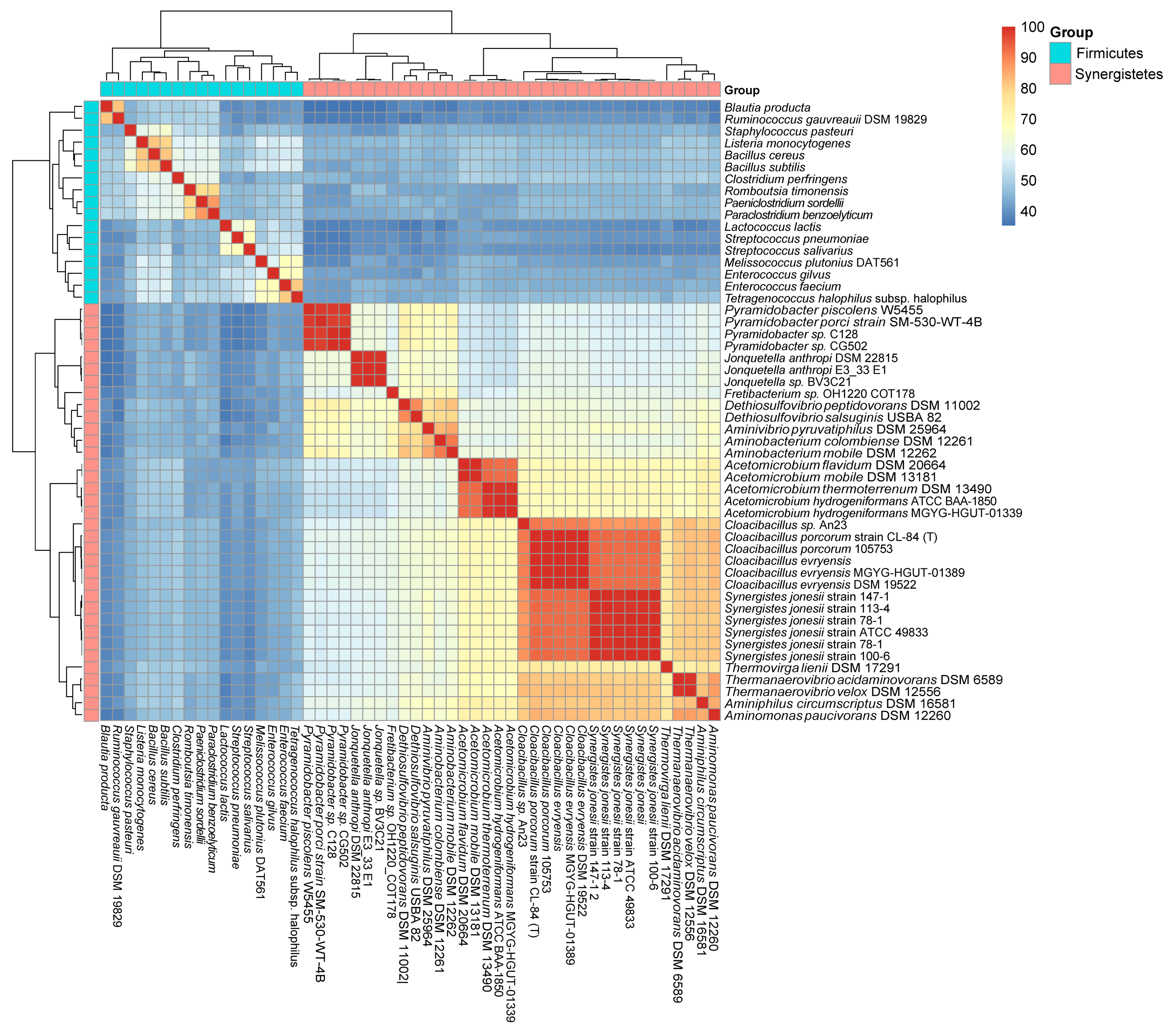
Figure 2.
The phylogenetic relationship of species and the CodY/CodY-like proteins of Firmicutes and Synergistetes. (A) Phylogenetic tree of the 16S rRNA of representative species from various bacterial phyla. The presence (grey) and absence (yellow) of a codY gene in the chromosome of a species is shown on the outer circle. Species of different phyla are color-coded and shown in the inner circle. The phylum information is shown on the right. An ML distance tree based on the 16S rRNA sequences of 51 species from different phyla is shown inside the circle. Gram-negative bacteria are indicated by purple branches and Gram-positive bacteria are indicated by blue branches. (B) An ML distance tree based on 52 CodY homologous protein sequences of different species in the phyla Firmicutes and Synergistetes. The phylum Synergistetes is indicated by the red branch and the phylum Firmicutes is indicated by the blue branch. The numbers on the branches indicate the percentage of calculations performed by bootstrap.
Figure 2.
The phylogenetic relationship of species and the CodY/CodY-like proteins of Firmicutes and Synergistetes. (A) Phylogenetic tree of the 16S rRNA of representative species from various bacterial phyla. The presence (grey) and absence (yellow) of a codY gene in the chromosome of a species is shown on the outer circle. Species of different phyla are color-coded and shown in the inner circle. The phylum information is shown on the right. An ML distance tree based on the 16S rRNA sequences of 51 species from different phyla is shown inside the circle. Gram-negative bacteria are indicated by purple branches and Gram-positive bacteria are indicated by blue branches. (B) An ML distance tree based on 52 CodY homologous protein sequences of different species in the phyla Firmicutes and Synergistetes. The phylum Synergistetes is indicated by the red branch and the phylum Firmicutes is indicated by the blue branch. The numbers on the branches indicate the percentage of calculations performed by bootstrap.
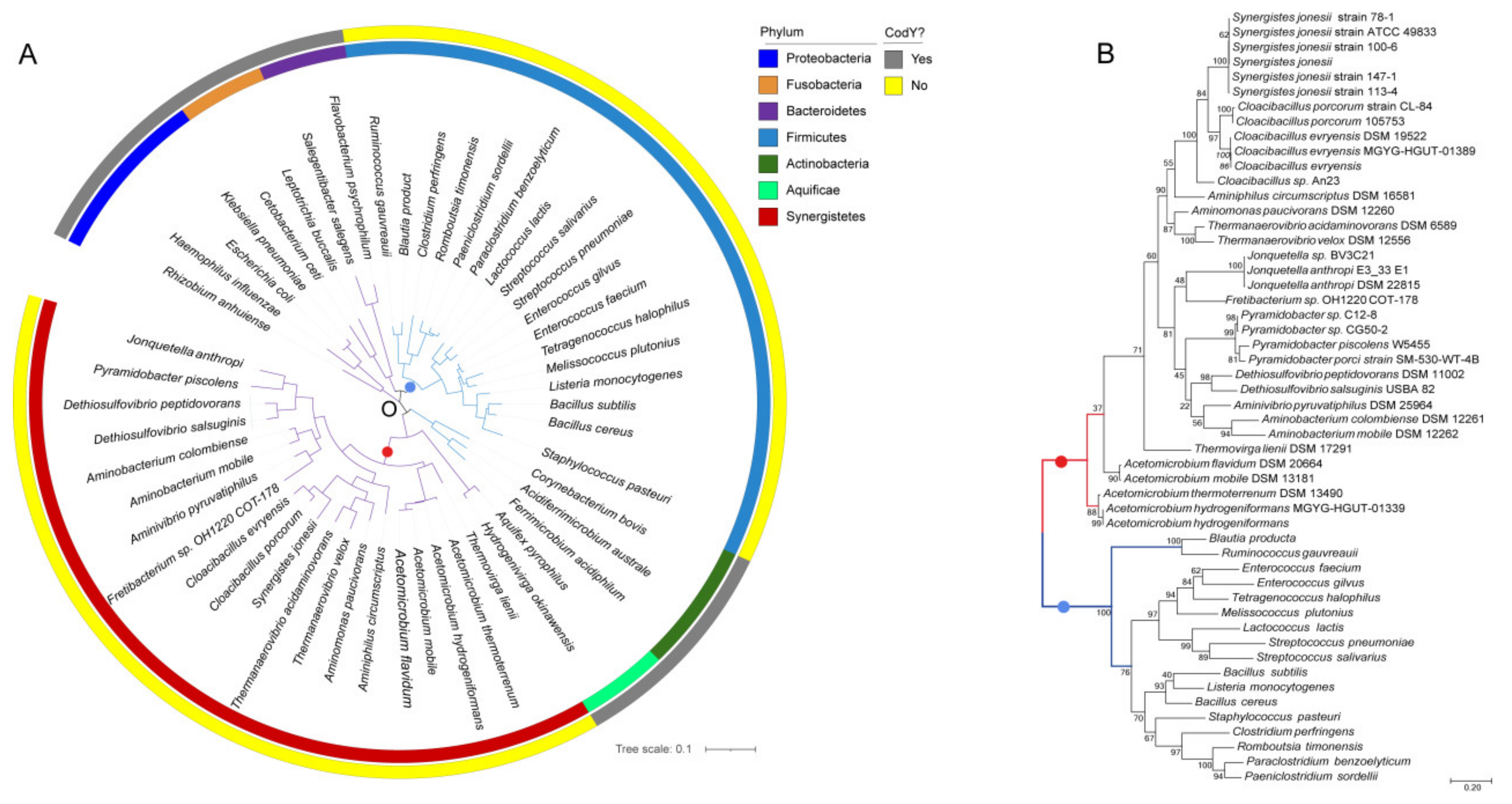
Figure 3.
Motif logos representing the binding sites of the CodY-like proteins for five genera in the phylum Synergistetes. The logos were generated by the MEME function of the MEME suite. All 40 core binding sequences of the CodY-like protein listed in Table S5 were used to analyze the common motifs in Synergistes and Cloacibacillus (A), Thermanaerovibrio (B), Pyramidobacter (C) and Acetomicrobium (D).
Figure 3.
Motif logos representing the binding sites of the CodY-like proteins for five genera in the phylum Synergistetes. The logos were generated by the MEME function of the MEME suite. All 40 core binding sequences of the CodY-like protein listed in Table S5 were used to analyze the common motifs in Synergistes and Cloacibacillus (A), Thermanaerovibrio (B), Pyramidobacter (C) and Acetomicrobium (D).
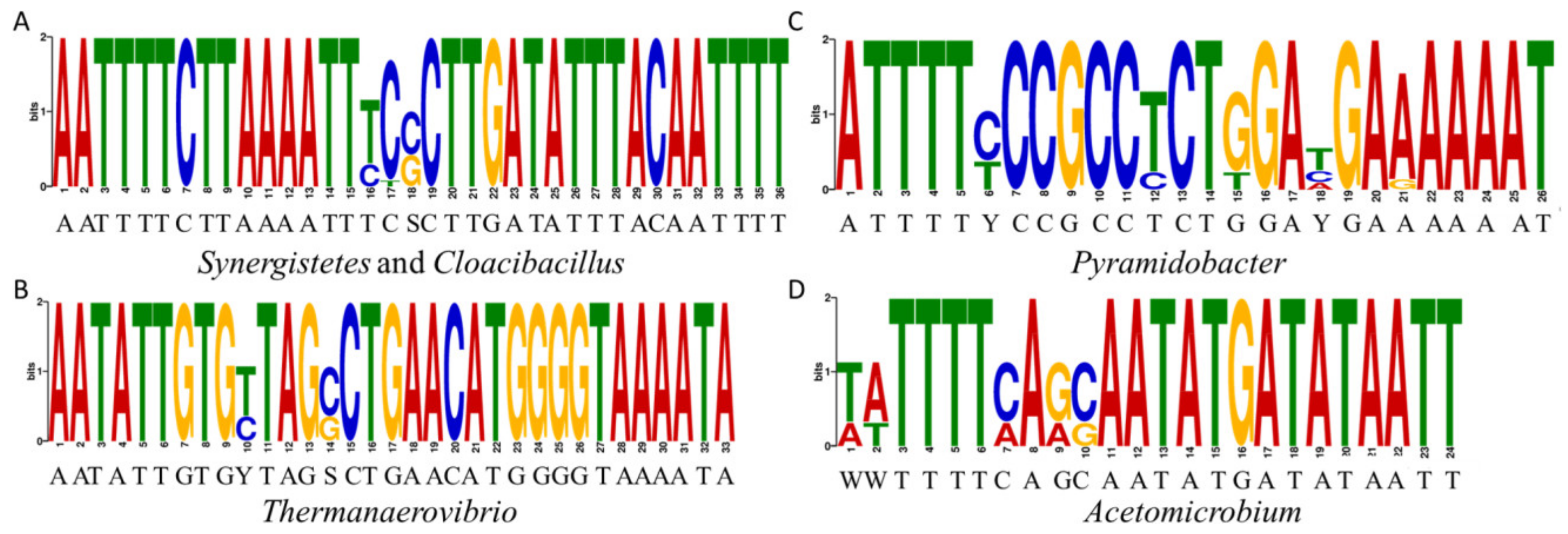
Figure 4.
DAPA experiments showing the binding specificity of the CodY-like protein of C. evryensis DSM19522. Lanes 1 to 4, 2.5 μM, 0.5 μM, 0.1 μM, and 0.02 μM of the purified His-CodYDSM were used in the binding reactions, respectively. Lane 5, 1.25% of the probe used in the reaction is shown to ensure a comparable number of labeled probes was used in each of the reactions. (I), the wild-type probe containing the predicted binding sequence 5′ to xerC of C. evryensis DSM1952 was used. (II), the 5′ AT-rich sequence of the probe is mutated. (III), the central AT-rich sequence of the probe is mutated. (IV), the 3′ AT-rich sequence of the probe is mutated. (V) and (VI), all three AT-rich sequences were mutated. The key features of the probes used in each reaction are listed right to the images. For clarity, only the sense strand of the probe is shown.
Figure 4.
DAPA experiments showing the binding specificity of the CodY-like protein of C. evryensis DSM19522. Lanes 1 to 4, 2.5 μM, 0.5 μM, 0.1 μM, and 0.02 μM of the purified His-CodYDSM were used in the binding reactions, respectively. Lane 5, 1.25% of the probe used in the reaction is shown to ensure a comparable number of labeled probes was used in each of the reactions. (I), the wild-type probe containing the predicted binding sequence 5′ to xerC of C. evryensis DSM1952 was used. (II), the 5′ AT-rich sequence of the probe is mutated. (III), the central AT-rich sequence of the probe is mutated. (IV), the 3′ AT-rich sequence of the probe is mutated. (V) and (VI), all three AT-rich sequences were mutated. The key features of the probes used in each reaction are listed right to the images. For clarity, only the sense strand of the probe is shown.
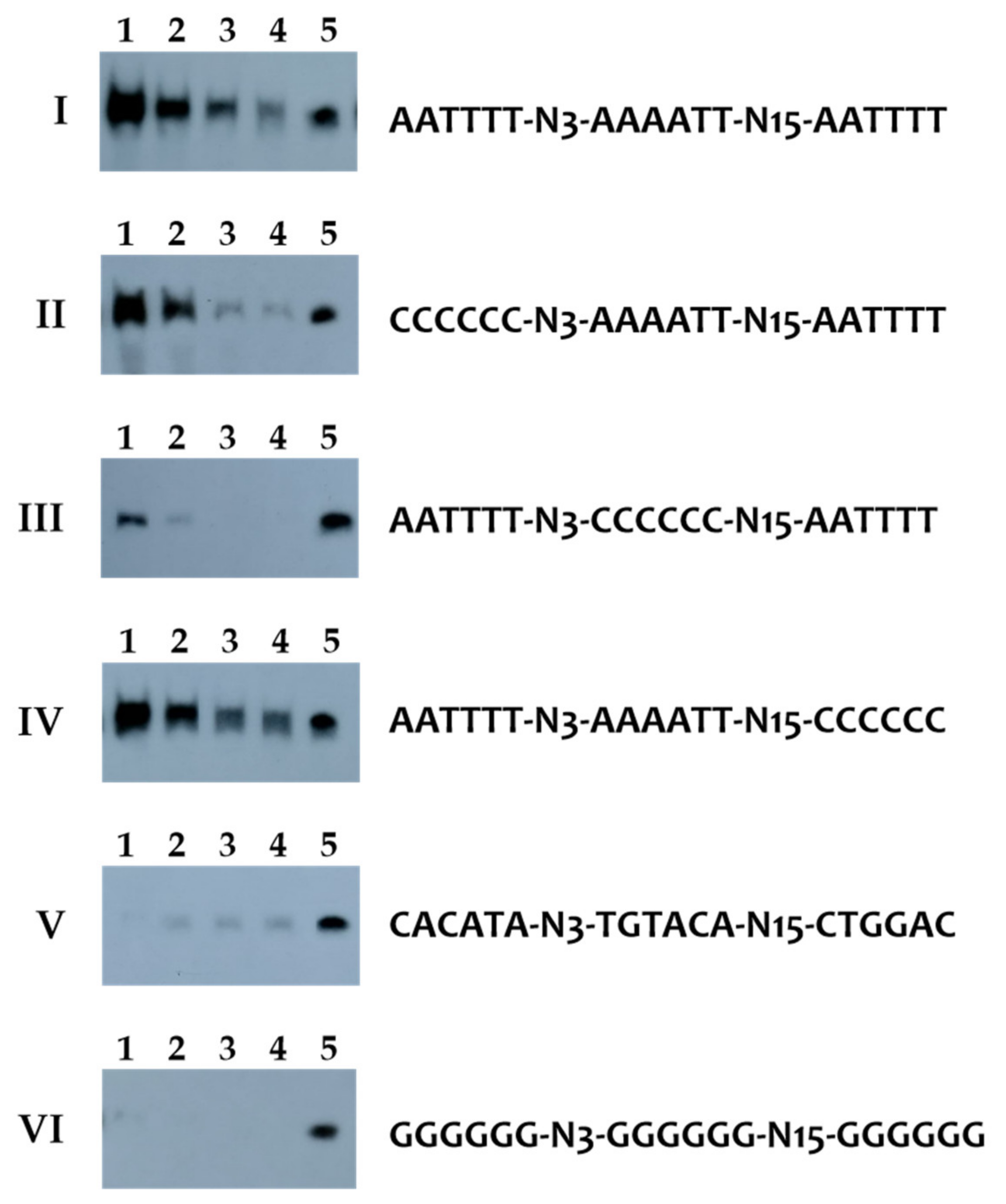
Figure 5.
The distribution of genes regulated by the CodY-like protein in the available genomes of the phylum Synergistetes. The number of loci that harbor the canonical CodY box, the newly defined extended palindromic sequence, and both motifs in the promoter regions are shown in orange, green and light orange, respectively. The percentages of total targets in the genomes are indicated.
Figure 5.
The distribution of genes regulated by the CodY-like protein in the available genomes of the phylum Synergistetes. The number of loci that harbor the canonical CodY box, the newly defined extended palindromic sequence, and both motifs in the promoter regions are shown in orange, green and light orange, respectively. The percentages of total targets in the genomes are indicated.

Figure 6.
A pathway-enrichment heat map showing the clustering of loci regulated by the CodY-like protein. The positions of the four genera are color-coded and listed on the top of the figure. The number of loci regulated by the CodY-like protein was analyzed based on the predicted function. The number was converted using log2 (N + 1), where the N is the number of the identified target genes.
Figure 6.
A pathway-enrichment heat map showing the clustering of loci regulated by the CodY-like protein. The positions of the four genera are color-coded and listed on the top of the figure. The number of loci regulated by the CodY-like protein was analyzed based on the predicted function. The number was converted using log2 (N + 1), where the N is the number of the identified target genes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The distribution of the putative CodY-like protein among the cultivable species of the phylum Synergistetes.
Table 1.
The distribution of the putative CodY-like protein among the cultivable species of the phylum Synergistetes.
| Locus_Tag | RefSeq | Assembly | GC (%) | Species |
|---|---|---|---|---|
| BUQ78_RS04225 | NZ_FSQZ01000001 | GCF_900129645 | 48.29 | Acetomicrobium flavidum DSM 20664 |
| HMPREF1705_RS06035 | NZ_ACJX03000001 | GCF_000160455 | 46.53 | Acetomicrobium hydrogeniformans |
| FY315_RS06110 | NZ_CABKOO010000001 | GCF_902374045 | 46.53 | Acetomicrobium hydrogeniformans MGYG-HGUT-01339 |
| ANAMO_RS01275 | NC_018024 | GCF_000266925 | 47.94 | Acetomicrobium mobile DSM 13181 |
| BLU12_RS09355 | NZ_FNPD01000014 | GCF_900107215 | 48.46 | Acetomicrobium thermoterrenum DSM 13490 |
| K349_RS0111180 | NZ_JAFY01000007 | GCF_000526375 | 59.16 | Aminiphilus circumscriptus DSM 16581 |
| C8D99_RS00920 | NZ_SORI01000001 | GCF_000165795 | 67.58 | Aminivibrio pyruvatiphilus DSM 25964 |
| AMICO_RS07740 | NC_014011 | GCF_000025885 | 45.31 | Aminobacterium colombiense DSM 12261 |
| K360_RS0102980 | NZ_JAFZ01000001 | GCF_000526395 | 43.88 | Aminobacterium mobile DSM 12262 |
| APAU_RS04160 | NZ_CM001022 | GCF_000165795 | 67.58 | Aminomonas paucivorans DSM 12260 |
| FXY42_RS11030 | NZ_CABKQM010000008 | GCF_902374565 | 56.26 | Cloacibacillus evryensis MGYG-HGUT-01389 |
| CLOEV_RS08630 | NZ_KK073872 | GCF_000585335 | 55.95 | Cloacibacillus evryensis DSM 19522 |
| HMPREF1006_RS10935 | NZ_JH414697 | GCF_000238615 | 56.26 | Cloacibacillus evryensis |
| BED41_RS09070 | NZ_CP016757 | GCF_001701045 | 54.84 | Cloacibacillus porcorum strain CL-84 (T) |
| HF883_RS06600 | NZ_JABAGT010000007 | GCF_012844265 | 56.42 | Cloacibacillus porcorum 105753 |
| B5F39_RS01840 | NZ_NFJQ01000002 | GCF_002159945 | 55.63 | Cloacibacillus sp. An23 |
| DPEP_RS01550 | NZ_ABTR02000001 | GCF_000172975 | 54.41 | Dethiosulfovibrio peptidovorans DSM 11002 |
| B9Y55_RS09045 | NZ_FXBB01000024 | GCF_900177735 | 55.24 | Dethiosulfovibrio salsuginis USBA 82 |
| EII26_RS10880 | NZ_RQYL01000028 | GCF_003860125 | 62.36 | Fretibacterium sp. OH1220_COT-178 |
| JONANDRAFT_RS04180 | NZ_CM001376 | GCF_000237805 | 59.50 | Jonquetella anthropic DSM 22815 |
| GCWU000246_RS04765 | NZ_GG697147 | GCF_000161995 | 59.09 | Jonquetella anthropic E3_33 E1 |
| HMPREF1249_RS06160 | NZ_AWWC01000024 | GCF_000468895 | 59.31 | Jonquetella sp. BV3C21 |
| HMPREF7215_RS03875 | NZ_ADFP01000047 | GCF_000177335 | 58.15 | Pyramidobacter piscolens W5455 |
| B0D78_RS09560 | NZ_MUHX01000026 | GCF_002007215 | 57.20 | Pyramidobacter sp. C12-8 |
| D7D26_RS08945 | NZ_RAWT01000042 | GCF_003612005 | 58.62 | Pyramidobacter sp. CG50-2 |
| FYJ74_RS00750 | NZ_VUNH01000001 | GCF_009695745 | 58.90 | Pyramidobacter porci strain SM-530-WT-4B |
| EH55_RS06015 | NZ_JMKI01000026 | GCF_000712295 | 55.86 | Synergistes jonesii strain 78-1 |
| JS79_RS06185 | NZ_JPZR01000022 | GCF_001757565 | 55.84 | Synergistes jonesii strain 147-1 |
| JS78_RS05700 | NZ_JPZS01000024 | GCF_001757415 | 55.84 | Synergistes jonesii strain 113-4 |
| JS77_RS05735 | NZ_JPZQ01000030 | GCF_001757495 | 55.85 | Synergistes jonesii strain 100-6 |
| JS73_RS05710 | NZ_JQEL01000027 | GCF_001757485 | 55.84 | Synergistes jonesii strain ATCC 49833 |
| JS72_RS09655 | NZ_JQEK01000046 | GCF_001757465 | 55.83 | Synergistes jonesii |
| TACI_RS06905 | NC_013522 | GCF_000024905 | 63.79 | Thermanaerovibrio acidaminovorans DSM 6589 |
| THEVEDRAFT_RS02110 | NZ_CM001377 | GCF_000237825 | 58.78 | Thermanaerovibrio velox DSM 12556 |
| TLIE_RS02745 | NC_016148 | GCF_000233775 | 47.10 | Thermovirga lienii DSM 17291 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Geng, J.; Luo, S.; Shieh, H.-R.; Wang, H.-Y.; Hu, S.; Chen, Y.-Y.M. Identification of a Putative CodY Regulon in the Gram-Negative Phylum Synergistetes. Int. J. Mol. Sci. 2022, 23, 7911. https://doi.org/10.3390/ijms23147911
AMA Style
Geng J, Luo S, Shieh H-R, Wang H-Y, Hu S, Chen Y-YM. Identification of a Putative CodY Regulon in the Gram-Negative Phylum Synergistetes. International Journal of Molecular Sciences. 2022; 23(14):7911. https://doi.org/10.3390/ijms23147911
Chicago/Turabian StyleGeng, Jianing, Sainan Luo, Hui-Ru Shieh, Hsing-Yi Wang, Songnian Hu, and Yi-Ywan M. Chen. 2022. "Identification of a Putative CodY Regulon in the Gram-Negative Phylum Synergistetes" International Journal of Molecular Sciences 23, no. 14: 7911. https://doi.org/10.3390/ijms23147911
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.