
Proteomic and Biochemical Approaches Elucidate the Role of Millimeter-Wave Irradiation in Wheat Growth under Flooding Stress
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
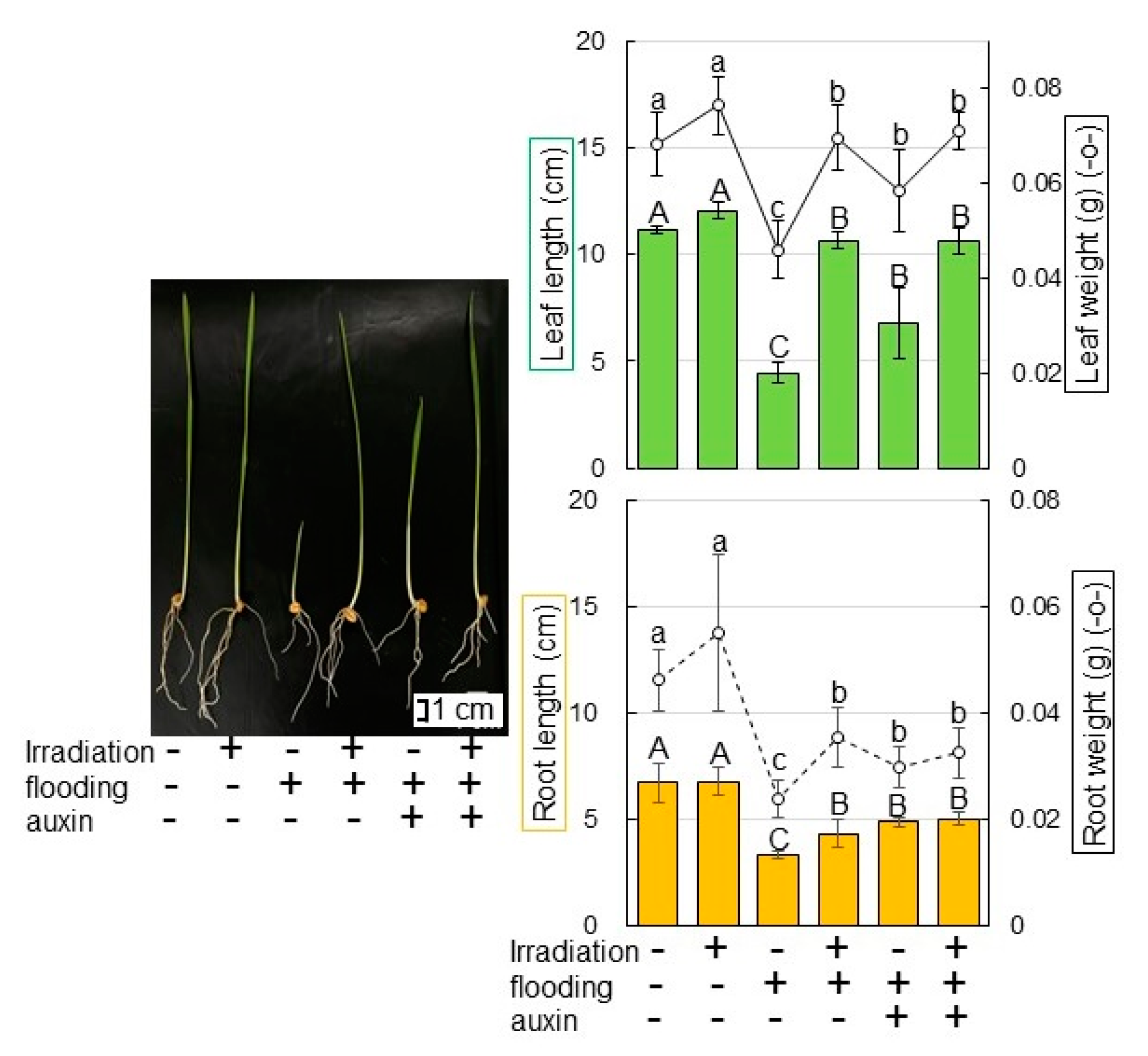
2.1. Morphological Changes of Wheat Irradiated with Millimeter Waves under Flooding Stress
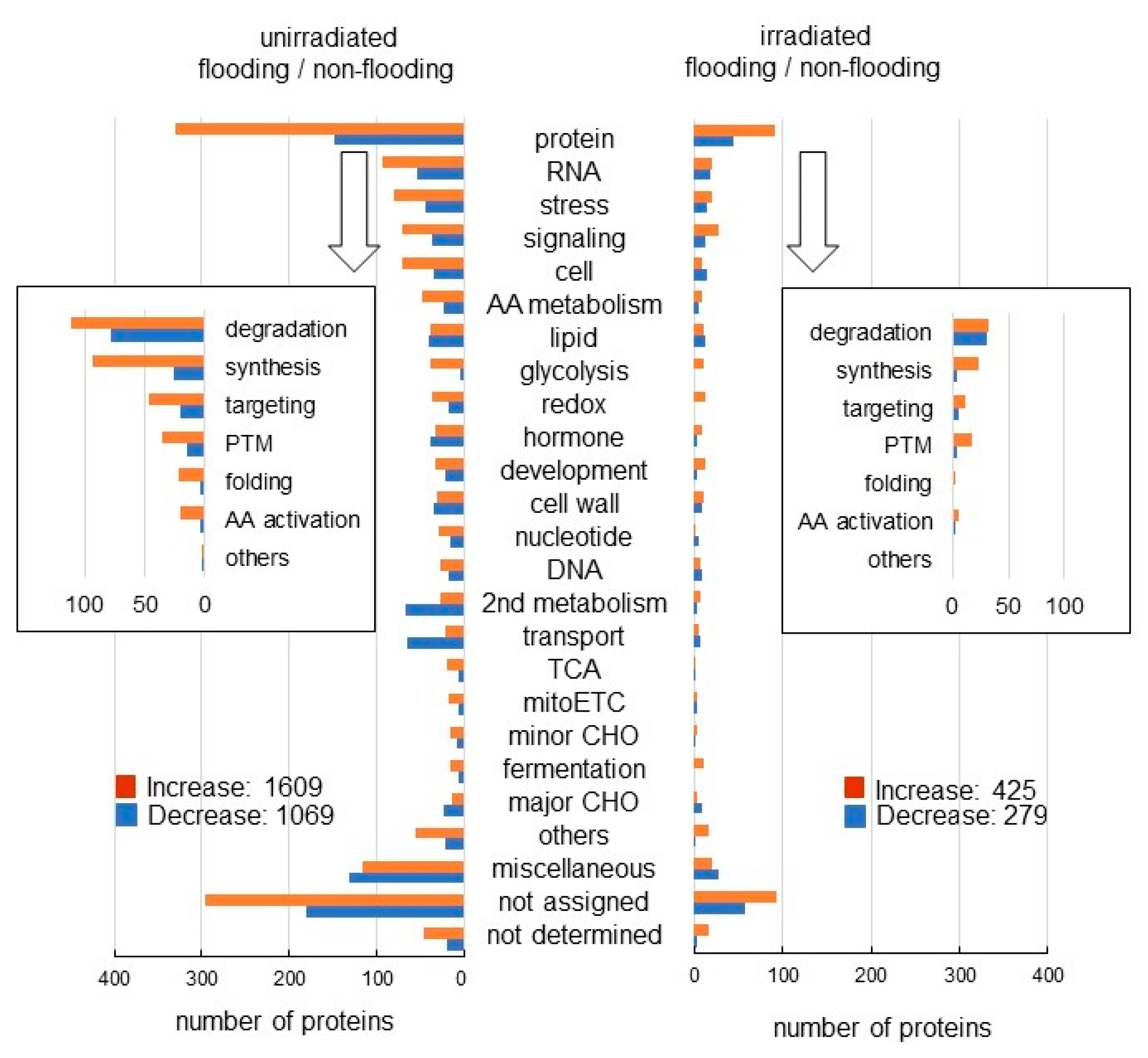
2.2. Protein Identification and Functional Categorization in Wheat Irradiated with Millimeter Waves under Flooding Stress
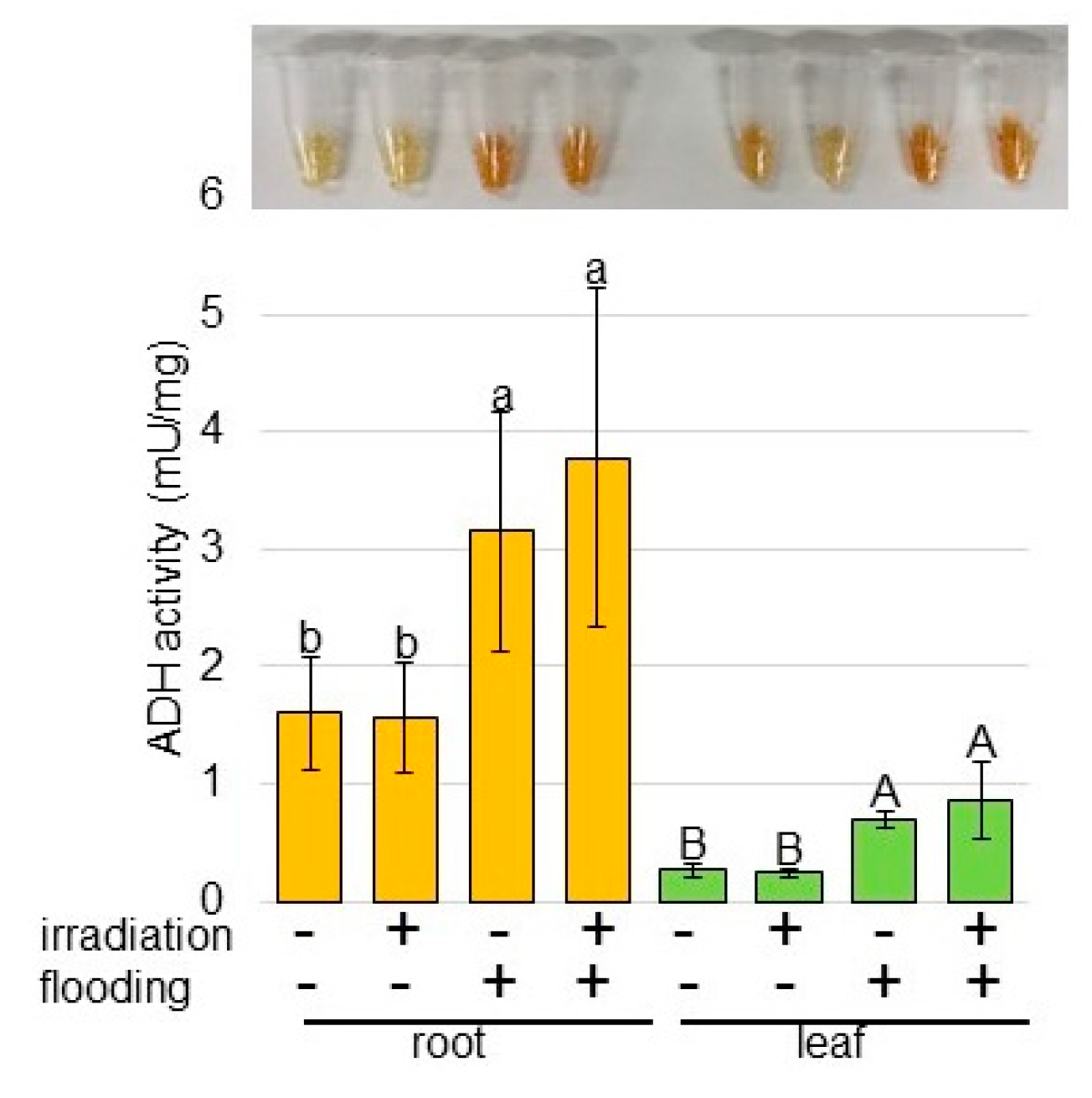
2.3. Activity of Alcohol Dehydrogenase (ADH) in Wheat Irradiated with Millimeter Waves under Flooding Stress
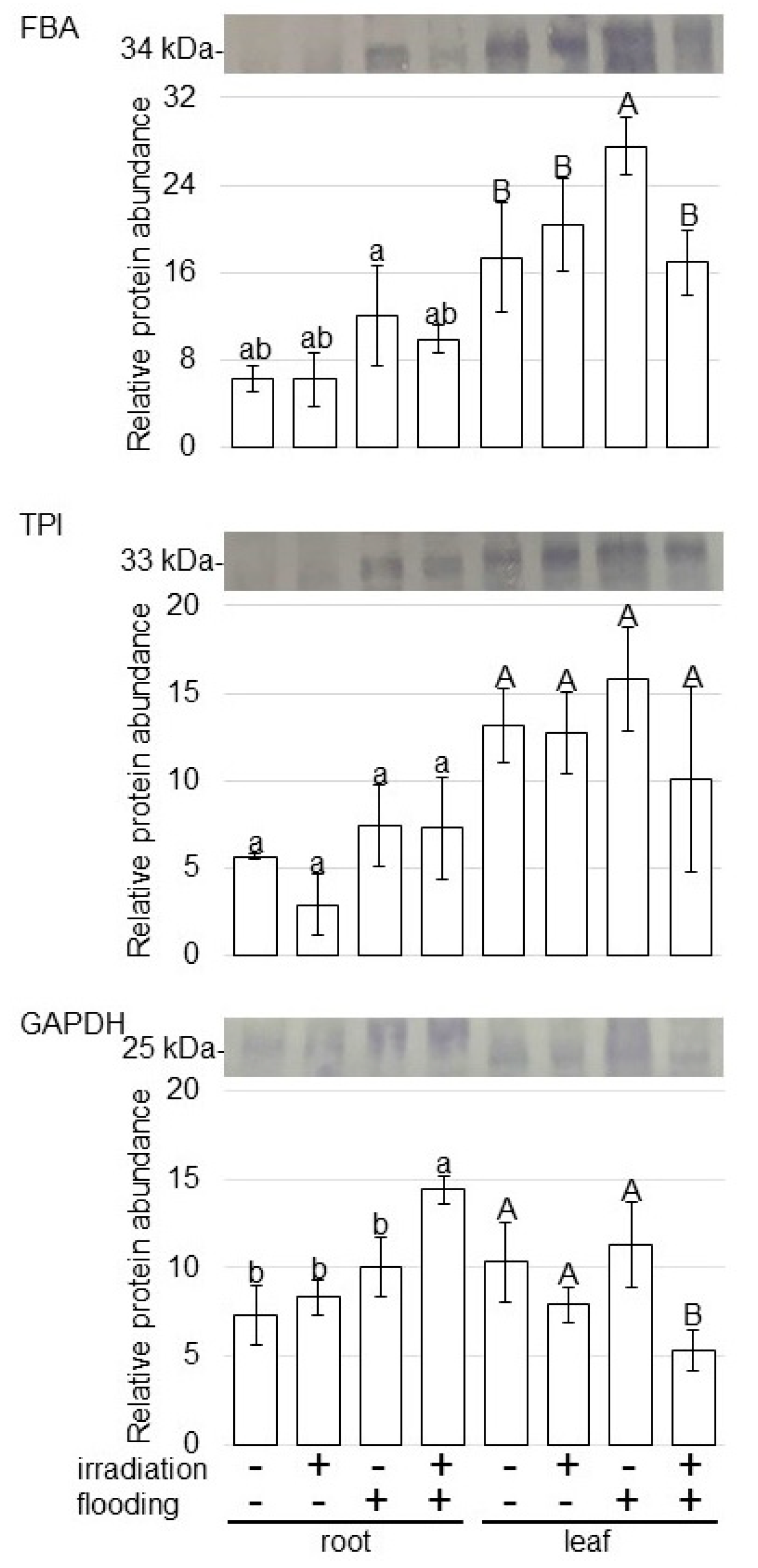
2.4. Abundance of Proteins Related to Glycolysis in Wheat Irradiated with Millimeter Waves under Flooding Stress
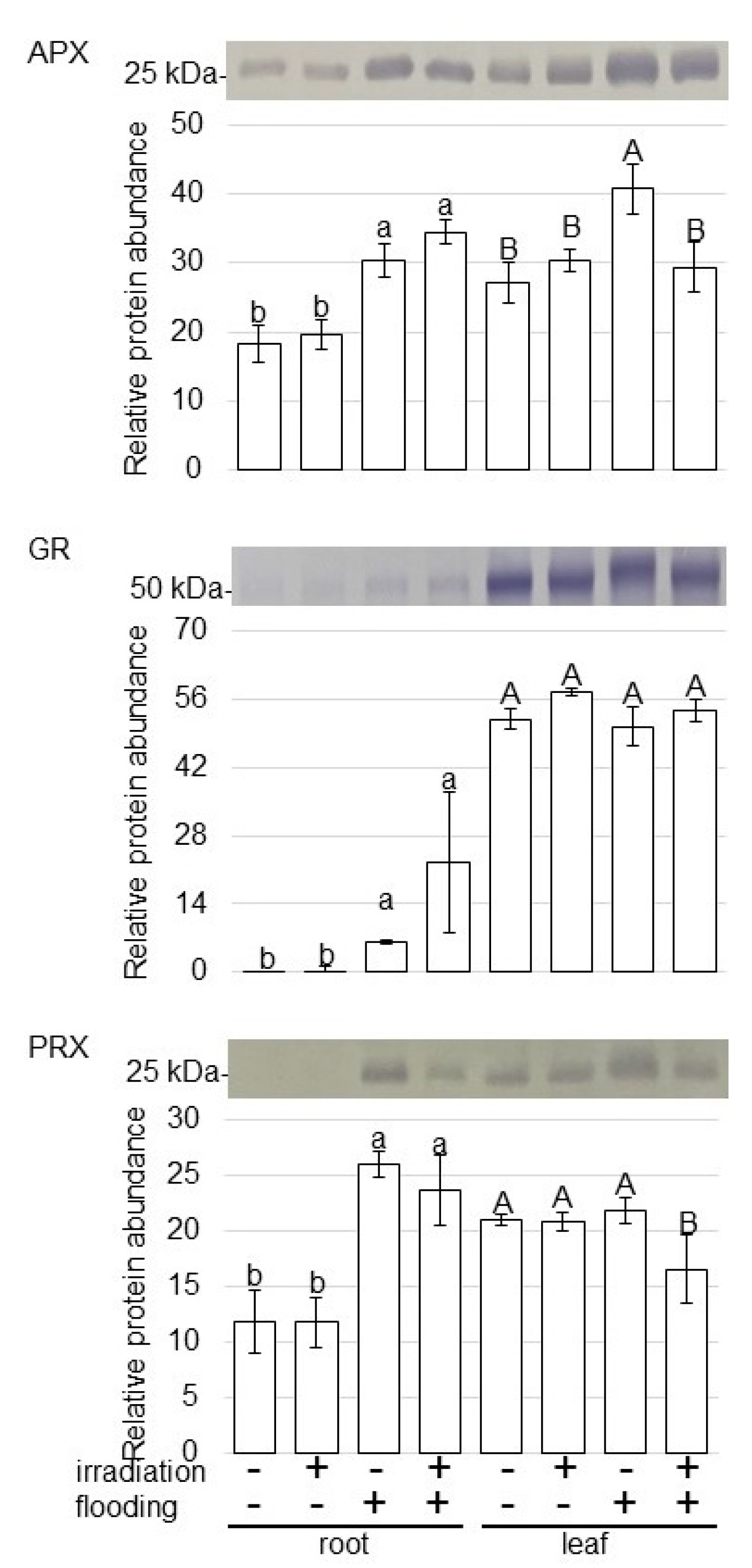
2.5. Abundance of Proteins Related to ROS Scavenging in Wheat Irradiated with Millimeter Waves under Flooding Stress
2.6. Abundance of Proteins Related to Cell Organization in Wheat Irradiated with Millimeter Waves under Flooding Stress
2.7. Morphological Changes of Wheat after Auxin Application under Flooding Stress
3. Discussion
3.1. Millimeter-Wave Irradiation Has a Positive Effect on Wheat Growth under Flooding Stress
3.2. Millimeter-Wave Irradiation Suppresses Glycolysis in Wheat under Flooding Stress
3.3. Millimeter-Wave Irradiation Suppresses ROS Scavenging and Cell Organization in Wheat under Flooding Stress
3.4. Millimeter-Wave Irradiation Regulates Auxin Metabolism in Wheat under Flooding Stress
4. Materials and Methods
4.1. Plant Material, Millimeter-Wave Irradiation, and Treatment
4.2. Protein Extraction
4.3. Protein Enrichment, Reduction, Alkylation, and Digestion
4.4. Protein Identification Using Nano-Liquid Chromatography (LC) Mass Spectrometry (MS)
4.5. MS Data Analysis
4.6. Differential Analysis of Proteins Using MS Data
4.7. Immunoblot Analysis
4.8. Measurement of Alcohol Dehydrogenase (ADH) Activity
4.9. Morphological Analysis after Auxin Application
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curtus, T.; Halford, N.G. Food security: The challenge of increasing wheat yield and the importance of not compromising food safety. Ann. Appl. Biol. 2014, 164, 354–372. [Google Scholar] [CrossRef]
- Sayre, K.; Van Ginkel, M.; Rajaram, S.; Ortiz-Monasterio, I. Tolerance to waterlogging losses in spring bread wheat: Effect of time of onset on expression. Ann. Wheat Newsl. 1994, 40, 165–171. [Google Scholar]
- Setter, T.L.; Waters, I. Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil 2013, 253, 1–34. [Google Scholar] [CrossRef]
- Herzog, M.; Striker, G.G.; Colmer, T.D.; Pedersen, O. Mechanisms of waterlogging tolerance in wheat: A review of root and shoot physiology. Plant Cell Environ. 2016, 39, 1068–1086. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, M.; Hirabayashi, Y.; Ikeuchi, H. Global-scale river flood vulnerability in the last 50 years. Sci. Rep. 2016, 6, 36021. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Barrera-Figuero, B.E.; Juntawong, P.; Pena-Castro, J.M. Submergence and waterlogging stress in plants: A review highlighting research opportunities and understudied aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef]
- Jia, W.; Ma, M.; Chen, J.; Wu, S. Plant morphological, physiological and anatomical adaption to flooding stress and the underlying molecular mechanisms. Int. J. Mol. Sci 2021, 22, 1088. [Google Scholar] [CrossRef]
- Sheen, D.M.; McMakin, D.L.; Hall, T.E. Detection of explosives by millimeter-wave imaging. Count. Detect. Tech. Explos. 2007, 237–277. [Google Scholar]
- Mignone, C.; Barnes, R. More than meets the eye: The electromagnetic spectrum. Sci. School 2011, 20, 51–59. [Google Scholar]
- Betskii, O.V.; Lebedeva, N.N.; Tambiev, A.H.; Kirikova, N.N.; Slavin, V.E. Millimeter waves in the newest agricultural biotechnologies. J. Sci. Eng. 2007, 23, 236–252. [Google Scholar]
- Sukiasyan, A.; Mikaelyan, Y.; Ayrapetyan, S. Comparative study of non-ionizing and ionizing radiation effect on hydration of winter wheat seeds in metabolic active and inactive states. Environmentalist 2012, 32, 188–192. [Google Scholar] [CrossRef]
- Poghosyan, G.H.; Mukhaelyan, Z.H. The influence of low-intensity EMI treatment on seed germination and early growth of theat. Proc. Yerevan State Univ. Chem. Biol. 2018, 52, 110–115. [Google Scholar]
- Mukhaelyan, Z.H.; Shahinyan, M.A.; Poghosyan, G.H.; Vardevanyan, P.O. Wheat seedlings growth and antioxidant system activity changes as responses to extremely high frequency electromagnetic irradiation. Am. J. Plant Bio. 2016, 2, 1–10. [Google Scholar]
- Zhong, Z.; Furuya, T.; Ueno, K.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Tani, M.; Tian, J.; Komatsu, S. Proteomic analysis of irradiation with millimeter waves on soybean growth under flooding conditions. Int. J. Mol. Sci. 2020, 21, 486. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Maruyama, J.; Furuya, T.; Yin, X.; Yamaguchi, H.; Hitachi, K.; Miyashita, N.; Tsuchida, K.; Tani, M. Proteomic and biological analyses reveal the effect on growth under flooding stress of chickpea irradiated with millimeter waves. J. Proteome Res. 2021, 20, 4718–4727. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.H.; Kim, M.S.; Choi, H.W.; Sung, J.M.; Park, J.D.; Kum, J.S. Effects of millimeter wave treatment on the germination rate and antioxidant potentials and gamma-aminobutyric acid of the germinated brown rice. Food Sci. Biotechnol. 2016, 25, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Wang, X.; Yin, X.; Tian, J.; Komatsu, S. Morphophysiological and proteomic responses on plants of irradiation with electromagnetic waves. Int. J. Mol. Sci. 2021, 22, 12239. [Google Scholar] [CrossRef] [PubMed]
- Ramundo-Orlando, A. Effects of millimeter waves radiation on cell membrane-a brief review. J. Infrared Millim. Terahertz Waves 2010, 31, 1400–1411. [Google Scholar] [CrossRef]
- Usatii, A.; Molodoi, E.; Rotaru, A.; Moldoveanu, T. The influence of low intensity millimeter waves on the multiplication and biosynthetic activity of Saccharomyces carlsbergensis CNMN-Y-15 yeast. Ann. Oradea Univ.—Biol. Fascicle 2010, 17, 208–212. [Google Scholar]
- Ishikawa, N. Recent progress and future tasks in wheat breeding at Kanto, Tokai and Western Japan. J. Crop. Res. 2010, 55, 1–8. [Google Scholar]
- Shimizu, K.K.; Copetti, D.; Okada, M.; Wicker, T.; Tameshige, T.; Hatakeyama, M.; Shimizu-Inatsugi, R.; Aquino, C.; Nishimura, K.; Kobayashi, F.; et al. De Novo Genome Assembly of the Japanese wheat cultivar Norin 61 highlights functional variation in flowering time and fusarium-resistant genes in East Asian genotypes. Plant Cell Physiol. 2021, 62, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Sachs, M.M.; Freeling, M.; Okimoto, R. The anaerobic proteins of maize. Cell 1980, 20, 761–767. [Google Scholar] [CrossRef]
- Kennedy, R.A.; Rumpho, M.E.; Fox, T.C. Anaerobic Metabolism in Plants. Plant Physiol. 1992, 100, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Greenway, H.; Matsumura, H.; Tsutsumi, N.; Nakazono, M. Rice alcohol dehydrogenase 1 promotes survival and has a major impact on carbohydrate metabolism in the embryo and endosperm when seeds are germinated in partially oxygenated water. Ann. Bot. 2014, 113, 851–859. [Google Scholar] [CrossRef]
- Komatsu, S.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Rehman, S.U.; Ohno, T. Morphological, biochemical, and proteomic analyses to understand the promotive effects of plant-derived smoke solution on wheat growth under flooding stress. Plants 2022, 11, 1508. [Google Scholar] [CrossRef]
- Komatsu, S.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Kono, Y.; Nishimura, M. Proteomic and biochemical analyses of the mechanism of tolerance in mutant soybean responding to flooding stress. Int. J. Mol. Sci. 2021, 22, 9046. [Google Scholar] [CrossRef]
- Mustafa, G.; Sakata, K.; Hossain, Z.; Komatsu, S. Proteomic study on the effects of silver nanoparticles on soybean under flooding stress. J. Proteom. 2015, 122, 100–118. [Google Scholar] [CrossRef]
- Sasidharan, R.; Hartman, S.; Liu, Z.; Martopawiro, S.; Sajeev, N.; Van Veen, H.; Yeung, E.; Voesenek, L.A.C.J. Signal dynamics and interactions during flooding stress. Plant Physiol. 2018, 176, 1106–1117. [Google Scholar] [CrossRef] [Green Version]
- Parlanti, S.; Kudahettige, N.P.; Lombardi, L.; Mensuali-Sodi, A.; Alpi, A.; Perata, P.; Pucciariello, C. Distinct mechanisms for aerenchyma formation in leaf sheaths of rice genotypes displaying a quiescence or escape strategy for flooding tolerance. Ann. Bot. 2011, 107, 1335–1343. [Google Scholar] [CrossRef]
- Oliveira, R.A.C.; de Andrade, A.S.; Imparato, D.O.; de Lima, J.G.S.; de Almeida, R.V.M.; Lima, J.P.M.S.; Pasquali, M.A.B.; Dalmolin, R.J.S. Analysis of Arabidopsis thaliana redox gene network indicates evolutionary expansion of class III peroxidase in plants. Sci. Rep. 2019, 9, 15741. [Google Scholar] [CrossRef]
- Baxter-Burrell, A.; Yang, Z.; Springer, P.S.; Bailey-Serres, J. RopGAP4-dependent Rop GTPase rheostat control of Arabidopsis oxygen deprivation tolerance. Science 2002, 296, 2026–2028. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.X.; Yu, M.; Zhang, N.; Zhou, Z.Q.; Xu, Q.T.; Mei, F.Z.; Qu, L.H. Reactive oxygen species regulate programmed cell death progress of endosperm in winter wheat (Triticum aestivum L.) under waterlogging. Protoplasma 2016, 253, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Livanos, P.; Apostolakos, P.; Galatis, B. Plant cell division: ROS homeostasis is required. Plant Signal Behav. 2012, 7, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Livanos, P.; Galatis, B.; Quader, H.; Apostolakos, P. Disturbance of reactive oxygen species homeostasis induces atypical tubulin polymer formation and affects mitosis in root-tip cells of Triticum turgidum and Arabidopsis thaliana. Cytoskeleton 2012, 69, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Livanos, P.; Galatis, B.; Gaitanaki, C.; Apostolakos, P. Phosphorylation of a p38-like MAPK is involved in sensing cellular redox state and drives atypical tubulin polymer assembly in angiosperms. Plant Cell Environ. 2014, 37, 1130–1143. [Google Scholar] [CrossRef] [PubMed]
- Nick, P. Microtubules and the tax payer. Protoplasma 2012, 249, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Kuroha, T.; Nagai, K.; Gamuyao, R.; Wang, D.R.; Furuta, T.; Nakamori, M.; Kitaoka, T.; Adachi, K.; Minami, A.; Mori, Y.; et al. Ethylene-gibberellin signaling underlies adaptation of rice to periodic flooding. Science 2018, 361, 181–186. [Google Scholar] [CrossRef]
- Du, H.; Chang, Y.; Huang, F.; Xiong, L. GID1 modulates stomatal response and submergence tolerance involving abscisic acid and gibberellic acid signaling in rice. J. Integr. Plant Biol. 2015, 57, 954–968. [Google Scholar] [CrossRef]
- Dawood, T.; Yang, X.; Visser, E.J.; Te Beek, T.A.; Kensche, P.R.; Cristescu, S.M.; Lee, S.; Floková, K.; Nguyen, D.; Mariani, C.; et al. A co-opted hormonal cascade activates dormant adventitious root primordia upon flooding in Solanum dulcamara. Plant Physiol. 2016, 170, 2351–2364. [Google Scholar] [CrossRef]
- Gao, J.; Zhuang, S.; Zhang, Y.; Qian, Z. Exogenously applied spermidine alleviates hypoxia stress in Phyllostachys praecox seedlings via changes in endogenous hormones and gene expression. BMC Plant Biol. 2022, 22, 200. [Google Scholar] [CrossRef]
- Tamang, B.G.; Li, S.; Rajasundaram, D.; Lamichhane, S.; Fukao, T. Overlapping and stress-specific transcriptomic and hormonal responses to flooding and drought in soybean. Plant J. 2021, 107, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hartman, S.; van Veen, H.; Zhang, H.; Leeggangers, H.A.C.F.; Martopawiro, S.; Bosman, F.; de Deugd, F.; Su, P.; Hummel, M.; et al. Ethylene augments root hypoxia tolerance via growth cessation and reactive oxygen species amelioration. Plant Physiol. 2022, kiac245. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Ni, R.; Wang, Y.; Sun, J.; Ma, M.; Bi, H. Effects of different growth regulators on the rooting of Catalpa bignonioides softwood cuttings. Life 2022, 12, 1231. [Google Scholar] [CrossRef] [PubMed]
- Thorat, S.A.; Kaniyassery, A.; Poojari, P.; Rangel, M.; Tantry, S.; Kiran, K.R.; Joshi, M.B.; Rai, P.S.; Botha, A.M.; Muthusamy, A. Differential gene expression and withanolides biosynthesis during in vitro and ex vitro growth of Withania somnifera (L.) Dunal. Front. Plant Sci. 2022, 13, 917770. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Komatsu, S.; Han, C.; Nanjo, Y.; Altaf-Un-Nahar, M.; Wang, K.; He, D.; Yang, P. Label-free quantitative proteomic analysis of abscisic acid effect in early-stage soybean under flooding. J. Proteome Res. 2013, 12, 4769–4784. [Google Scholar] [CrossRef]
- Li, X.; Rehman, S.U.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Yamaguchi, T.; Sunohara, Y.; Matsumoto, H.; Komatsu, S. Proteomic analysis of the effect of plant-derived smoke on soybean during recovery from flooding stress. J. Proteom. 2018, 181, 238–248. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Komatsu, S.; Yamamoto, A.; Nakamura, T.; Nouri, M.Z.; Nanjo, Y.; Nishizawa, K.; Furukawa, K. Comprehensive analysis of mitochondria in roots and hypocotyls of soybean under flooding stress using proteomics and metabolomics techniques. J. Proteome Res. 2011, 10, 3993–4004. [Google Scholar] [CrossRef]
- Yin, X.; Komatsu, S. Comprehensive analysis of response and tolerant mechanisms in early-stage soybean at initial-flooding stress. J. Proteom. 2017, 169, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Konishi, H.; Yamane, H.; Maeshima, M.; Komatsu, S. Characterization of fructose-bisphosphate aldolase regulated by gibberellin in roots of rice seedling. Plant Mol. Biol. 2004, 56, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.A.; Côté, R.G.; Csordas, A.; Dianes, J.A.; Fabregat, A.; Foster, J.M.; Griss, J.; Alpi, E.; Birim, M.; Contell, J.; et al. The PRoteomics IDEntifications (PRIDE) database and associated tools: Status in 2013. Nucleic Acids Res. 2013, 41, D1063–D1069. [Google Scholar] [CrossRef]
- Okuda, S.; Watanabe, Y.; Moriya, Y.; Kawano, S.; Yamamoto, T.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Araki, N.; Yoshizawa, A.C.; et al. jPOSTTrepo: An international standard data repository for proteomes. Nucleic Acids Res. 2017, 45, D1107–D1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komatsu, S.; Tsutsui, Y.; Furuya, T.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Tani, M. Proteomic and Biochemical Approaches Elucidate the Role of Millimeter-Wave Irradiation in Wheat Growth under Flooding Stress. Int. J. Mol. Sci. 2022, 23, 10360. https://doi.org/10.3390/ijms231810360
Komatsu S, Tsutsui Y, Furuya T, Yamaguchi H, Hitachi K, Tsuchida K, Tani M. Proteomic and Biochemical Approaches Elucidate the Role of Millimeter-Wave Irradiation in Wheat Growth under Flooding Stress. International Journal of Molecular Sciences. 2022; 23(18):10360. https://doi.org/10.3390/ijms231810360
Chicago/Turabian StyleKomatsu, Setsuko, Yoshie Tsutsui, Takashi Furuya, Hisateru Yamaguchi, Keisuke Hitachi, Kunihiro Tsuchida, and Masahiko Tani. 2022. "Proteomic and Biochemical Approaches Elucidate the Role of Millimeter-Wave Irradiation in Wheat Growth under Flooding Stress" International Journal of Molecular Sciences 23, no. 18: 10360. https://doi.org/10.3390/ijms231810360