
TcMYB8, a R3-MYB Transcription Factor, Positively Regulates Pyrethrin Biosynthesis in Tanacetum cinerariifolium

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of TcMYB8 and Its Promoter
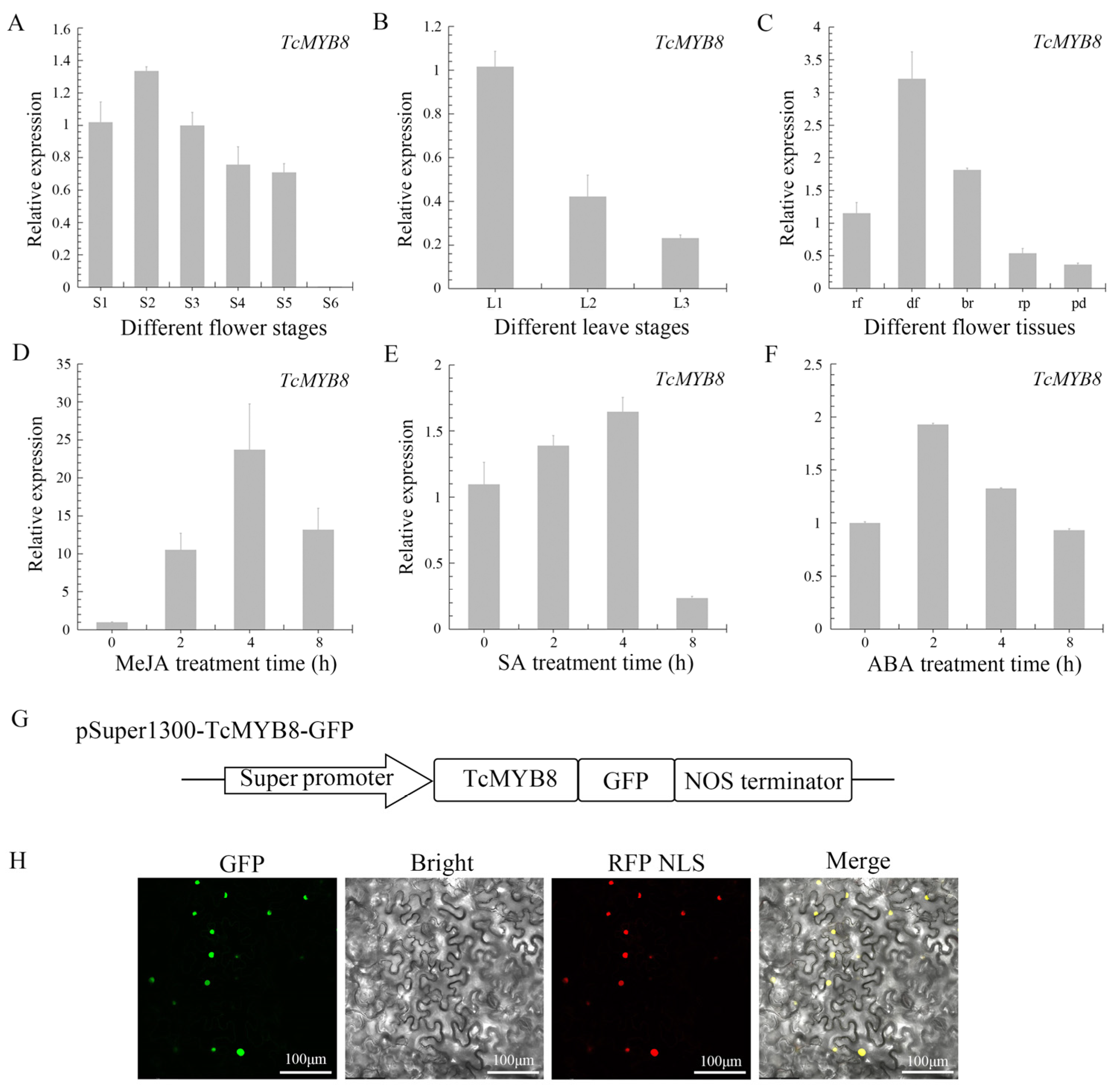
2.2. Expression Analysis and Subcellular Localization of TcMYB8
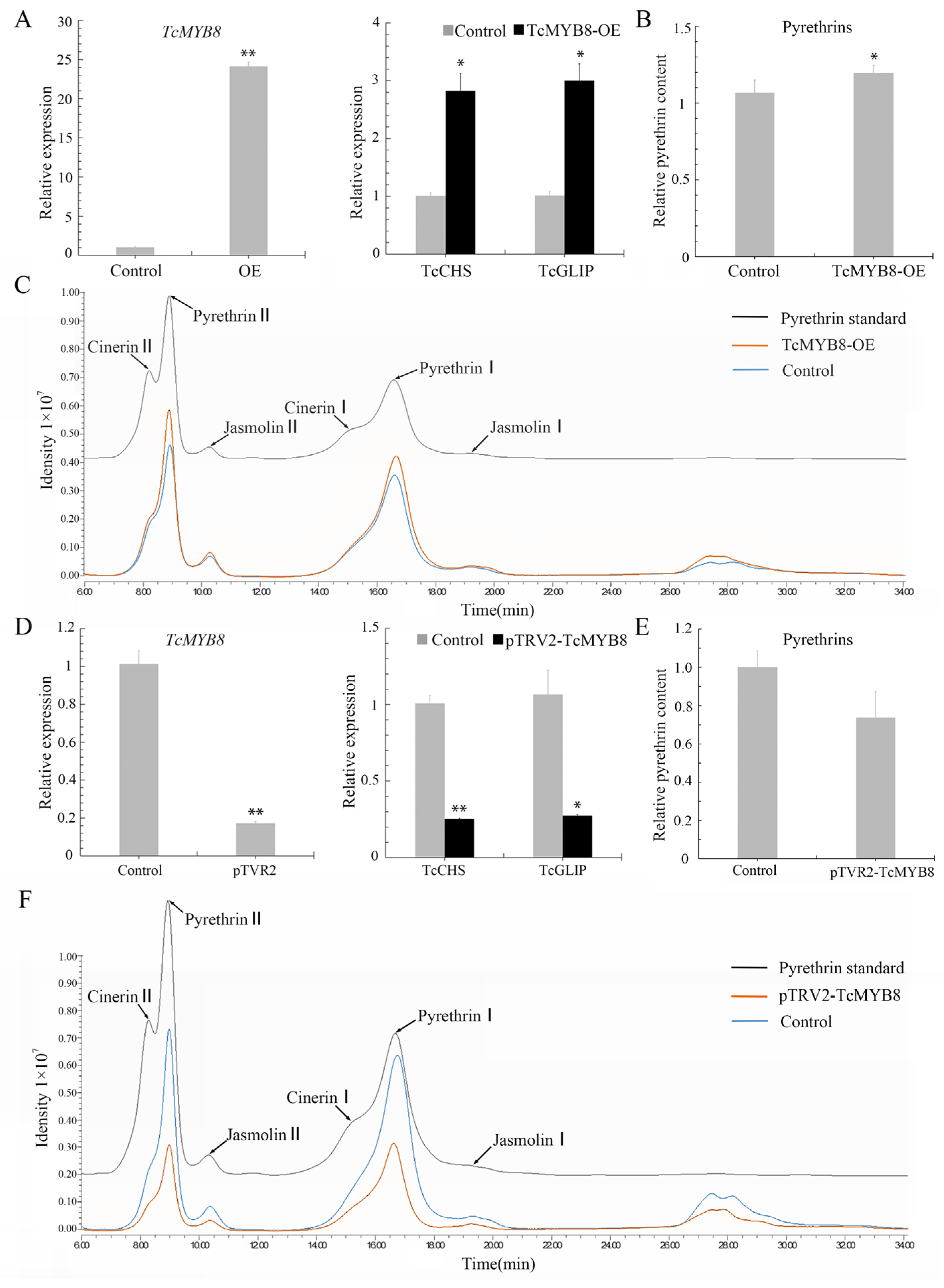
2.3. TcMYB8 Expression Promotes Pyrethrin Biosynthesis
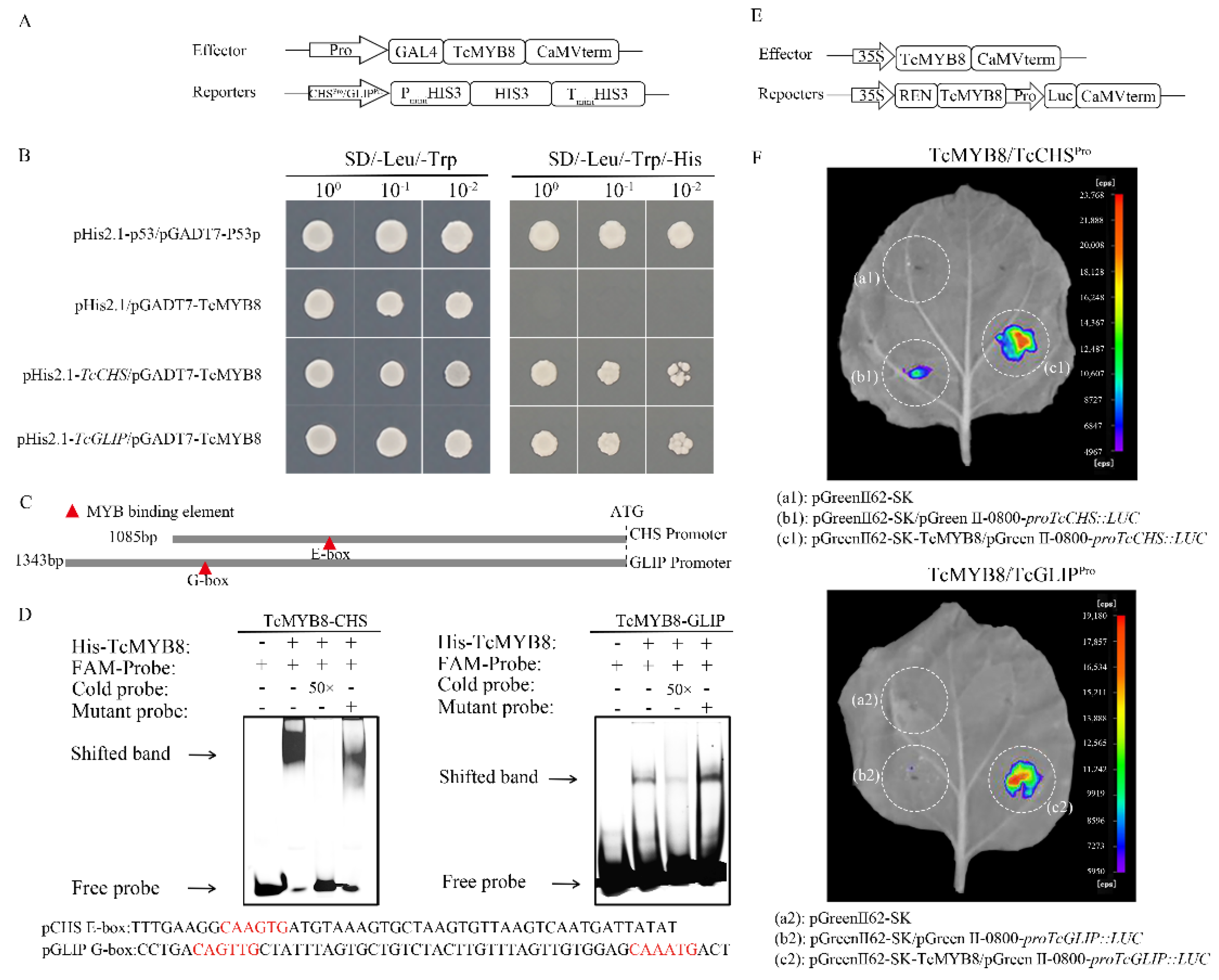
2.4. TcMYB8 Directly Binds to the Promoters of TcCHS and TcGLIP Genes
3. Discussion
3.1. TcMYB8, a 1R-MYB TF, Is a Novel MYB TF Found in T. cinerariifolium
3.2. TcMYB8 Activates the Transcription of TcCHS and TcGLIP to Modulate Pyrethrin Content in T. cinerariifolium
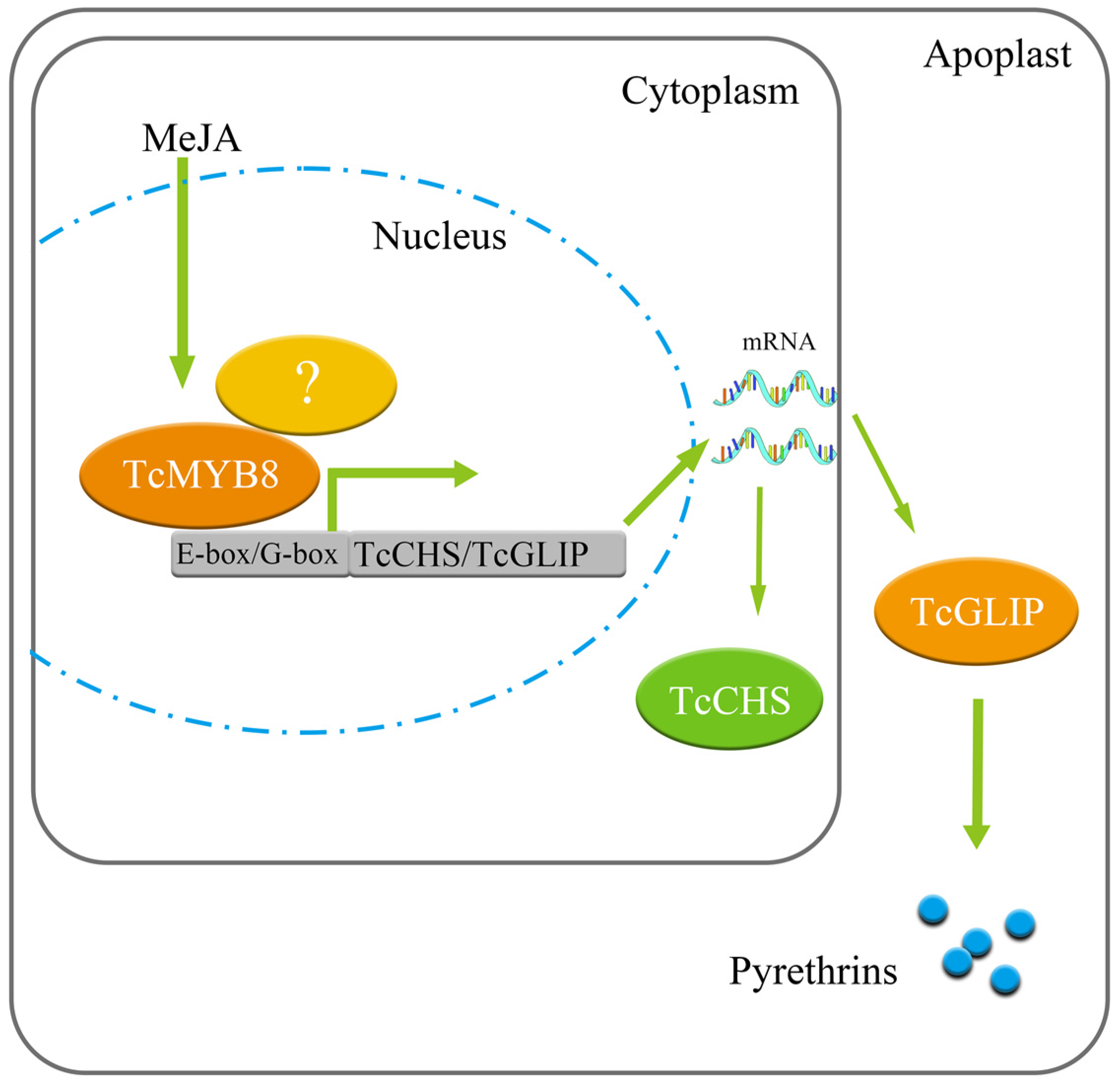
3.3. How TcMYB8 Regulates Pyrethrin Biosynthesis
4. Materials and Methods
4.1. Plant Materials, Growth Environment, and Phytohormone Treatments
4.2. RNA Extraction and Gene Expression Analysis
4.3. Gene Cloning and Sequence Analysis
4.4. Subcellular Localization Analysis
4.5. Transient Overexpression of TcMYB8 in T. cinerariifolium Leaves
4.6. VIGS Assay
4.7. Determination of Pyrethrin Content
4.8. Y1H Assay
4.9. Dual-LUC Reporter Assay
4.10. EMSA
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pavela, R. History, Presence and Perspective of Using Plant Extracts as Commercial Botanical Insecticides and Farm Products for Protection against Insects—A Review. Plant Prot. Sci. 2016, 52, 229–241. [Google Scholar]
- Huggins, C. Discipline, Governmentality and ‘Developmental Patrimonialism’: Insights from Rwanda’s Pyrethrum Sector. J. Agrar. Chang. 2017, 17, 715–732. [Google Scholar] [CrossRef]
- Gallo, M.; Formato, A.; Ianniello, D.; Andolfi, A.; Conte, E.; Ciaravolo, M.; Varchetta, V.; Naviglio, D. Supercritical Fluid Extraction of Pyrethrins from Pyrethrum Flowers (Chrysanthemum cinerariifolium) Compared to Traditional Maceration and Cyclic Pressurization Extraction. J. Supercrit. Fluids 2017, 119, 104–112. [Google Scholar] [CrossRef]
- Suraweera, D.D.; Groom, T.; Taylor, P.W.J.; Jayasinghe, C.S.; Nicolas, M.E. Dynamics of Flower, Achene and Trichome Development Governs the Accumulation of Pyrethrins in Pyrethrum (Tanacetum cinerariifolium) under Irrigated and Dryland Conditions. Ind. Crops Prod. 2017, 109, 123–133. [Google Scholar] [CrossRef]
- Zeng, T.; Li, J.W.; Zhou, L.; Xu, Z.Z.; Li, J.J.; Hu, H.; Luo, J.; Zheng, R.R.; Wang, Y.Y.; Wang, C.Y. Transcriptional Responses and GCMS Analysis for the Biosynthesis of Pyrethrins and Volatile Terpenes in Tanacetum coccineum. Int. J. Mol. Sci. 2021, 22, 13005. [Google Scholar] [CrossRef]
- Lybrand, D.B.; Xu, H.; Last, R.L.; Pichersky, E. How Plants Synthesize Pyrethrins: Safe and Biodegradable Insecticides. Trends Plant Sci. 2020, 25, 1240–1251. [Google Scholar] [CrossRef]
- Hitmi, A.; Coudret, A.; Barthomeuf, C. The production of Pyrethrins by Plant Cell and Tissue Cultures of Chrysanthemum cinerariaefolium and Tagetes Species. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 317–337. [Google Scholar] [CrossRef] [PubMed]
- Lal, R.K.; Chanotiya, C.S.; Gupta, P.; Mishra, A.; Gupta, M.M. Genetic Improvement of Pyrethrum (Tanacetum cinerariifolium sch. Bip.) Through Gamma Radiation and Selection of High Yield Stable Mutants Through Seven Post-Radiation Generations. Int. J. Radiat. Biol. 2021, 97, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jongsma, M.A.; Wang, C.-Y. Comparative Analysis of Pyrethrin Content Improvement by Mass Selection, Family Selection and Polycross in Pyrethrum Tanacetum cinerariifolium (Trevir.) Sch.Bip. Populations. Ind. Crops Prod. 2014, 53, 268–273. [Google Scholar] [CrossRef]
- Rivera, S.B.; Swedlund, B.D.; King, G.J.; Bell, R.N.; Hussey, C.E.; Shattuck-Eidens, D.M.; Wrobel, W.M.; Peiser, G.D.; Poulter, C.D. Chrysanthemyl Diphosphate Synthase: Isolation of the Gene and Characterization of the Recombinant Non-Head-to-Tail Monoterpene Synthase from Chrysanthemum cinerariaefolium. Proc. Natl. Acad. Sci. USA 2001, 98, 4373–4378. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Gao, L.; Hu, H.; Stoopen, G.; Wang, C.; Jongsma, M.A. Chrysanthemyl Diphosphate Synthase Operates in Planta as a Bifunctional Enzyme with Chrysanthemol Synthase Activity. J. Biol. Chem. 2014, 289, 36325–36335. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Moghe, G.D.; Wiegert-Rininger, K.; Schilmiller, A.L.; Barry, C.S.; Last, R.L.; Pichersky, E. Coexpression Analysis Identifies Two Oxidoreductases Involved in the Biosynthesis of the Monoterpene Acid Moiety of Natural Pyrethrin Insecticides in Tanacetum cinerariifolium. Plant Physiol. 2018, 176, 524–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, A.M.; Yang, T.; Bouwmeester, H.J.; Jongsma, M.A. A Trichome-Specific Linoleate Lipoxygenase Expressed During Pyrethrin Biosynthesis in Pyrethrum. Lipids 2013, 48, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Song, W.C.; Funk, C.D.; Brash, A.R. Molecular Cloning of an Allene Oxide Synthase: A Cytochrome-p450 Specialized for the Metabolism of Fatty–Acid Hydroperoxides. Proc. Natl. Acad. Sci. USA 1993, 90, 8519–8523. [Google Scholar] [CrossRef] [Green Version]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and Action of Jasmonates in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 355–381. [Google Scholar] [CrossRef] [Green Version]
- Tijet, N.; Brash, A.R. Allene Oxide Synthases and Allene Oxides. Prostaglandins Other Lipid Mediat. 2002, 68–69, 423–431. [Google Scholar] [CrossRef]
- Li, W.; Lybrand, D.B.; Zhou, F.; Last, R.L.; Pichersky, E. Pyrethrin Biosynthesis: The Cytochrome P450 Oxidoreductase CYP82Q3 Converts Jasmolone To Pyrethrolone. Plant Physiol. 2019, 181, 934–944. [Google Scholar] [CrossRef] [Green Version]
- Kikuta, Y.; Ueda, H.; Takahashi, M.; Mitsumori, T.; Yamada, G.; Sakamori, K.; Takeda, K.; Furutani, S.; Nakayama, K.; Katsuda, Y.; et al. Identification and Characterization of a GDSL Lipase–Like Protein that Catalyzes the Ester–Forming Reaction for Pyrethrin Biosynthesis in Tanacetum Cinerariifolium—A New Target for Plant Protection. Plant J. 2012, 71, 183–193. [Google Scholar] [CrossRef]
- Li, J.; Xu, Z.; Zeng, T.; Zhou, L.; Li, J.; Hu, H.; Luo, J.; Wang, C. Overexpression of TcCHS Increases Pyrethrin Content When Using a Genotype-Independent Transformation System in Pyrethrum (Tanacetum cinerariifolium). Plants 2022, 11, 1575. [Google Scholar] [CrossRef]
- Sugisaka, Y.; Aoyama, S.; Kumagai, K.; Ihara, M.; Matsuda, K. TcGLIP GDSL Lipase Substrate Specificity Co-determines the Pyrethrin Composition in Tanacetum cinerariifolium. J Agric. Food Chem. 2022, 70, 8645–8652. [Google Scholar] [CrossRef]
- Simon, M.; Tamara, G. Regulation of Glucosinolate Biosynthesis. J. Exp. Bot. 2020, 72, 70–91. [Google Scholar]
- Xiao, R.; Zhang, C.; Guo, X.; Li, H.; Lu, H. MYB Transcription Factors and Its Regulation in Secondary Cell Wall Formation and Lignin Biosynthesis during Xylem Development. Int. J. Mol. Sci. 2021, 22, 3560. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, L.-J.; Wang, Y.; Geng, Z.; Liu, S.; Chen, C.; Chen, S.; Jiang, J.; Chen, F. CmMYB9a Activates Floral Coloration by Positively Regulating Anthocyanin Biosynthesis in Chrysanthemum. Plant Mol. Biol. 2021, 108, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, X.; Xu, X.; Wang, F.; Zhao, X. The MYB Transcription Factor CiMYB42 Regulates Limonoids Biosynthesis in Citrus. BMC Plant Biol. 2020, 20, 254. [Google Scholar] [CrossRef]
- Reddy, V.A.; Wang, Q.; Dhar, N.; Kumar, N.; Venkatesh, P.N.; Rajan, C.; Panicker, D.; Sridhar, V.; Mao, H.-Z.; Sarojam, R. Spearmint R2R3-MYB Transcription Factor MsMYB Negatively Regulates Monoterpene Production and Suppresses the Expression of Geranyl Diphosphate Synthase Large Subunit (MsGPPS.LSU). Plant Biotechnol. J. 2017, 15, 1105–1119. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Luo, X.; Zhang, C.; Xu, X.; Huang, J.; Chen, Y.; Feng, S.; Zhan, X.; Zhang, L.; Yuan, H.; et al. Tissue–Specific Study Across the Stem of Taxus Media Identifies a Phloem–Specific TmMYB3 Involved in the Transcriptional Regulation of Paclitaxel Biosynthesis. Plant J. 2020, 103, 95–110. [Google Scholar] [CrossRef]
- Yin, J.; Sun, L.; Li, Y.; Xiao, J.; Zhan, Y. Functional Identification of BpMYB21 and BpMYB61 Transcription Factors Responding to MeJA and SA in birch Triterpenoid Synthesis. BMC Plant Biol. 2020, 20, 374. [Google Scholar] [CrossRef]
- Xie, L.; Yan, T.; Li, L.; Chen, M.; Hassani, D.; Li, Y.; Qin, W.; Liu, H.; Chen, T.; Fu, X.; et al. An HD-ZIP-MYB Complex Regulates Glandular Secretory Trichome Initiation in Artemisia annua. New Phytol. 2021, 231, 2050–2064. [Google Scholar] [CrossRef]
- Wu, Z.; Li, L.; Liu, H.; Yan, X.; Tang, K. AaMYB15, an R2R3-MYB TF in Artemisia annua, Acts as a Negative Regulator of Artemisinin Biosynthesis. Plant Sci. 2021, 308, 110920. [Google Scholar] [CrossRef]
- Olsson, M.E.; Olofsson, L.M.; Lindahl, A.L.; Lundgren, A.; Brodelius, M.; Brodelius, P.E. Localization of Enzymes of Artemisinin Biosynthesis to the Apical Cells of Glandular Secretory Trichomes of Artemisia annua L. Phytochemistry 2009, 70, 1123–1128. [Google Scholar] [CrossRef]
- Yang, R.; Wang, S.; Zou, H.; Li, L.; Li, Y.; Wang, D.; Xu, H.; Cao, X. R2R3-MYB Transcription Factor SmMYB52 Positively Regulates Biosynthesis of Salvianolic Acid B and Inhibits Root Growth in Salvia miltiorrhiza. Int. J. Mol. Sci. 2021, 22, 9538. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB Transcription Factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yan, T.; Shen, Q.; Lu, X.; Pan, Q.; Huang, Y.; Tang, Y.; Fu, X.; Liu, M.; Jiang, W.; et al. Glandular Trichome–Specific WRKY1 Promotes Artemisinin Biosynthesis in Artemisia annua. New Phytol. 2017, 214, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Lee, M.M.; Lin, Y.; Gish, L.; Schiefelbein, J. Distinct and Overlapping Roles of Single–Repeat MYB Genes in Root Epidermal Patterning. Dev. Biol. 2007, 311, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, R.; Bulley, S.M.; Zhong, C.; Li, D. Kiwifruit MYBS1–Like and GBF3 Transcription Factors Influence L-Ascorbic Acid Biosynthesis by Activating Transcription of GDP-L-Galactose Phosphorylase 3. New Phytol 2022, 234, 1782–1800. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Yamagishi, M.; Matsuyama, K. Repression of Anthocyanin Biosynthesis by R3-MYB Transcription Factors in Lily (Lilium spp.). Plant Cell Rep. 2019, 38, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Jeran, N.; Grdiša, M.; Varga, F.; Šatović, Z.; Liber, Z.; Dabić, D.; Biošić, M. Correction to: Pyrethrin from Dalmatian Pyrethrum (Tanacetum cinerariifolium (Trevir.) Sch. Bip.): Biosynthesis, Biological Activity, Methods of Extraction and Determination. Phytochem. Rev. 2021, 20, 907. [Google Scholar] [CrossRef]
- Sakamori, K.; Ono, N.; Ihara, M.; Suzuki, H.; Matsuura, H.; Tanaka, K.; Ohta, D.; Kanaya, S.; Matsuda, K. Selective Regulation of Pyrethrin Biosynthesis by the Specific Blend of Wound Induced Volatiles in Tanacetum cinerariifolium. Plant Signal. Behav. 2016, 11, e1149675. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhou, F.; Pichersky, E. Jasmone Hydroxylase, a Key Enzyme in the Synthesis of the Alcohol Moiety of Pyrethrin Insecticides. Plant Physiol. 2018, 177, 1498–1509. [Google Scholar] [CrossRef] [Green Version]
- Zeng, T.; Li, J.-W.; Xu, Z.-Z.; Zhou, L.; Li, J.-J.; Yu, Q.; Luo, J.; Chan, Z.-L.; Jongsma, M.A.; Hu, H.; et al. TcMYC2 Regulates Pyrethrin Biosynthesis in Tanacetum cinerariifolium. Hortic. Res. 2022, 9, uhac178. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Li, X.P.; Zhang, T.Q.; Wang, Y.Y.; Wang, C.; Gao, C.Q. Overexpression of ThMYB8 Mediates Salt Stress Tolerance by Directly Activating Stress–Responsive Gene Expression. Plant Sci. 2021, 302, 110668. [Google Scholar] [CrossRef] [PubMed]
- Judd, R.; Bagley, M.C.; Li, M.; Zhu, Y.; Lei, C.; Yuzuak, S.; Ekelöf, M.; Pu, G.; Zhao, X.; Muddiman, D.C.; et al. Artemisinin Biosynthesis in Non-glandular Trichome Cells of Artemisia annua. Mol. Plant 2019, 12, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.T.; Voinnet, O.; Baulcombe, D.C. Initiation and Maintenance of Virus–Induced Gene Silencing. Plant Cell 1998, 10, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Zhang, L.; Tang, K. New Insights into Artemisinin Regulation. Plant Signal. Behav. 2017, 12, e1366398. [Google Scholar] [CrossRef]
- Zhou, L.; Huang, Y.; Wang, Q.; Guo, D. AaHY5 ChIP–seq Based on Transient Expression System Reveals the Role of AaWRKY14 in Artemisinin Biosynthetic Gene Regulation. Plant Physiol. Biochem. 2021, 168, 321–328. [Google Scholar] [CrossRef]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-Domain Proteins Interact with the WD-Repeat/bHLH/MYB Complexes to Regulate Jasmonate–Mediated Anthocyanin Accumulation and Trichome Initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Wang, Z.; Ye, S.; Li, J.; Zheng, B.; Bao, M.; Ning, G. Fusion Primer and Nested Integrated PCR (FPNI-PCR): A New High-Efficiency Strategy for Rapid Chromosome Walking or Flanking Sequence Cloning. BMC Biotechnol. 2011, 11, 109. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Wang, H.; Chen, S.; Ren, S.; Fu, H.; Li, R.; Wang, C. CmNAC73 Mediates the Formation of Green Color in Chrysanthemum Flowers by Directly Activating the Expression of Chlorophyll Biosynthesis Genes HEMA1 and CRD1. Genes 2021, 12, 704. [Google Scholar] [CrossRef]
- Jung, S.K.; McDonald, K.A.; Dandekar, A.M. Effect of Leaf Incubation Temperature Profiles on Agrobacterium Tumefaciens-Mediated Transient Expression. Biotechnol. Prog. 2015, 31, 783–790. [Google Scholar] [CrossRef]
- Hu, H.; Li, J.; Delatte, T.; Vervoort, J.; Gao, L.; Verstappen, F.; Xiong, W.; Gan, J.; Jongsma, M.A.; Wang, C. Modification of Chrysanthemum Odour and Taste with Chrysanthemol Synthase Induces Strong Dual Resistance Against Cotton aphids. Plant Biotechnol. J. 2018, 16, 1434–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Luo, D.; Zhang, F. DcWRKY75 Promotes Ethylene Induced Petal Senescence in Carnation (Dianthus caryophyllus L.). Plant J. 2021, 108, 1473–1492. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Li, J.; Zeng, T.; Xu, Z.; Luo, J.; Zheng, R.; Wang, Y.; Wang, C. TcMYB8, a R3-MYB Transcription Factor, Positively Regulates Pyrethrin Biosynthesis in Tanacetum cinerariifolium. Int. J. Mol. Sci. 2022, 23, 12186. https://doi.org/10.3390/ijms232012186
Zhou L, Li J, Zeng T, Xu Z, Luo J, Zheng R, Wang Y, Wang C. TcMYB8, a R3-MYB Transcription Factor, Positively Regulates Pyrethrin Biosynthesis in Tanacetum cinerariifolium. International Journal of Molecular Sciences. 2022; 23(20):12186. https://doi.org/10.3390/ijms232012186
Chicago/Turabian StyleZhou, Li, Jiawen Li, Tuo Zeng, Zhizhuo Xu, Jing Luo, Riru Zheng, Yuanyuan Wang, and Caiyun Wang. 2022. "TcMYB8, a R3-MYB Transcription Factor, Positively Regulates Pyrethrin Biosynthesis in Tanacetum cinerariifolium" International Journal of Molecular Sciences 23, no. 20: 12186. https://doi.org/10.3390/ijms232012186