
Running from Stress: Neurobiological Mechanisms of Exercise-Induced Stress Resilience
 , , , , and
, , , , and
Abstract
:1. Introduction
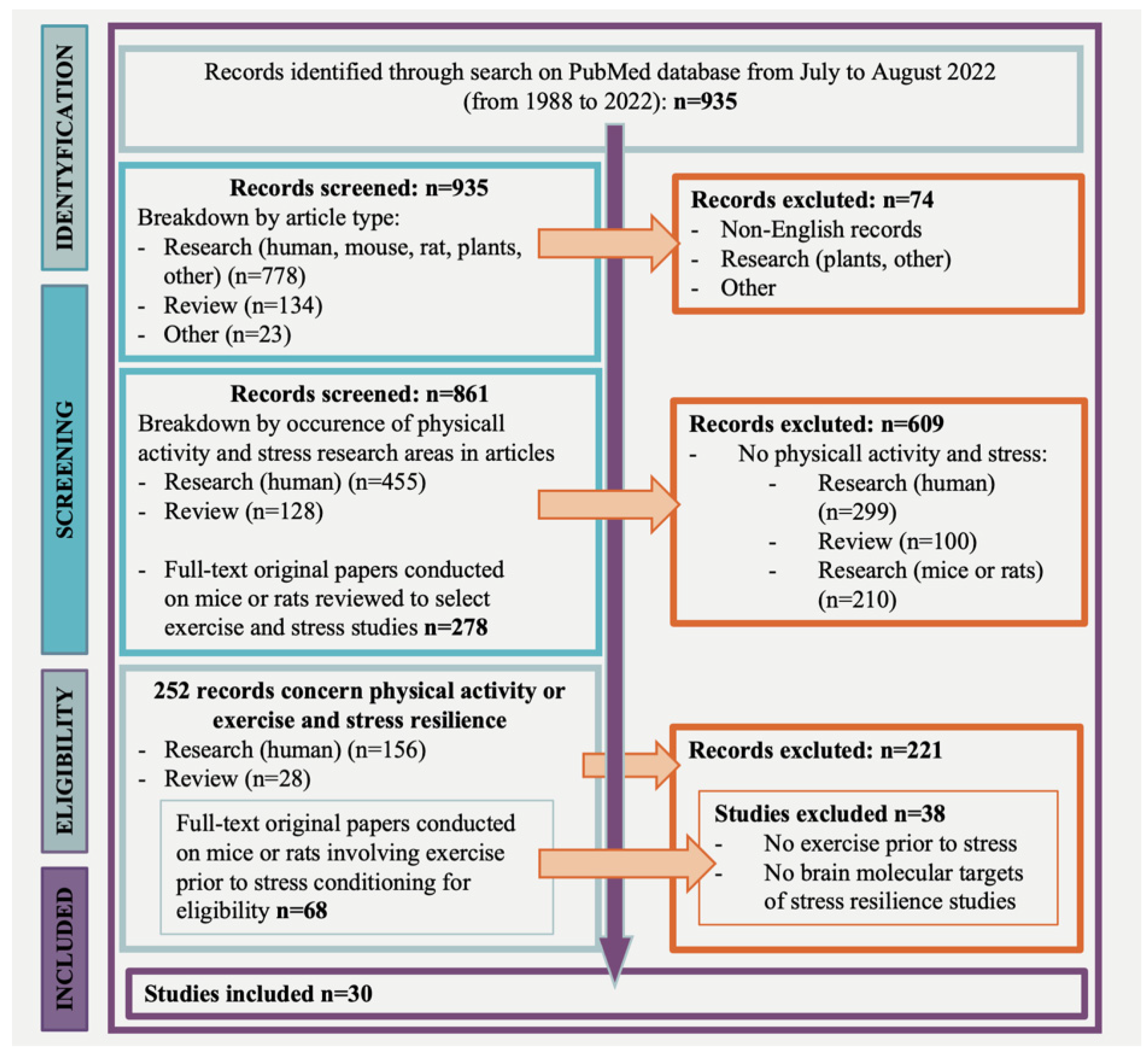
2. Literature Search Strategy
3. Results
4. Discussion
4.1. Neurogenesis, Neuroplasticity, and Cognition

{kind=link}
{kind=link}
| No. | Strain | Sex | Age | Physical Activity | Stress Model | Main Outcome | Reference |
|---|---|---|---|---|---|---|---|
| 1 | C57BL/6J | male | 5 weeks | swimming | unpredictable mild stress | modulation of AKT/GSK-3β/CRMP2 pathway; maintaining microtubule dynamics | [58] |
| 2 | C57BL/6J | male | N/A | treadmill | restraint stress | attenuated hippocampal cell death; CREB activation; decreased ROS levels | [60] |
| 3 | Long–Evans | female | 7 weeks | voluntary wheel running | traumatic stress | ameliorated stress-induced maladaptive behaviours | [57] |
| 4 | Sprague–Dawley | male | 2.5 months | treadmill | predator scent stress | improved morphological features of neurons | [59] |
4.2. Neurotransmitter Systems
| No. | Strain | Sex | Age | Physical Activity | Stress Model | Main Outcome | Reference |
|---|---|---|---|---|---|---|---|
| 1 | C57BL/6J | male | 7–8 weeks | voluntary wheel running | chronic social defeat stress | increased TH expression and number of TH-positive neurons in the VTA | [23] |
| 2 | C57BL/6 | male | 10 weeks | voluntary wheel running | chronic social defeat stress | induced ΔFosB in the subregions of the NAc | [66] |
| 3 | Sprague–Dawley | female | N/A | voluntary wheel running | uncontrollable tail shock | alleviated behavioural consequences of stress | [70] |
| 4 | Fischer F344 | male | 7–8 weeks | voluntary wheel running | uncontrollable tail shock | attenuated the dorsal raphe nucleus 5-HT activity | [71] |
| 5 | Sprague–Dawley | female | 4 weeks | voluntary wheel running | food-restriction stress | enhanced the GABAergic inhibition of pyramidal neurons in the dorsal hippocampus | [75] |
4.3. Neurotrophic Factors
| No. | Strain | Sex | Age | Physical Activity | Stress Model | Main Outcome | Reference |
|---|---|---|---|---|---|---|---|
| 1 | C57BL/6 | male | 8 weeks | voluntary wheel running | acute restraint stress | elevated expression of BDNF transcripts in the hippocampus | [94] |
| 2 | C57BL/6 | male and female | 8–9 weeks | treadmill | forced swim and tail suspension | increased IGF-1 level in the hippocampus | [99] |
| 3 | C57BL/6J | male | 2.5–3.5 months | voluntary wheel running | physical restraint | increased Adcy1 expression in the forebrain | [101] |
| 4 | Sprague–Dawley | male | 7–8 weeks | treadmill | predator scent stress | increased expression of BDNF, NPY, and DOR signalling in the hippocampus | [95] |
| 5 | Sprague–Dawley | male | 8 weeks | voluntary wheel running | depression | enhanced hippocampal neurogenesis; increased synaptic protein levels | [97] |
| 6 | Wistar | male and female | 7–8 weeks | treadmill | posttraumatic stress disorder | increased BDNF level and apoptotic markers in the hippocampus | [102] |
4.4. Hypothalamic–Pituitary–Adrenal Axis Activity
| No. | Strain | Sex | Age | Physical Activity | Stress Model | Main Outcome | Reference |
|---|---|---|---|---|---|---|---|
| 1 | C57BL/6 | male | 2 months | voluntary wheel running | restraint stress | modulation of serum CORT level, increased BDNF protein level in the hippocampus | [5] |
| 2 | C57BL/6J | male | 6 weeks | voluntary wheel running | restraint stress | reduced CORT response duration; increased adrenal sensitivity | [109] |
| 3 | C57BL/6 | N/A | 8 weeks | voluntary wheel running | social stress (housing conditions) | increased hippocampal expression of glucocorticoid receptor (Nr3c1), correlated with a down-regulation of miR-124 | [111] |
| 4 | Wistar and Sprague–Dawley | male | N/A | treadmill | social stress | decreased serum CORT levels induced by stress; alleviated behavioural consequences of stress | [108] |
| 5 | Sprague–Dawley | male | 2 months | voluntary wheel running | low-intensity stressors | reduced HPA responses to stressors | [104] |
| 6 | Sprague–Dawley | female | 40 days | treadmill | restraint stress | augmented NE decrease in the amygdala, hippocampus, and LC after which modulate the regulation of ACTH | [110] |
| 7 | Wistar | male | 2 months | voluntary wheel running | repeated noise stress | regulation of CRF and BDNF expression in several brain regions | [112] |
| 8 | Wistar | male | 3 months | swimming | chronic social isolation or immobilization or cold (4 °C) | enhanced GR and Hsp70 levels in the hippocampus and brain cortex | [113] |
| 9 | Fischer F344 | male | 8–9 weeks | voluntary wheel running | tail-shock stress | facilitated induction of HSP7 in the brain | [114] |
4.5. Oxidative Stress
| No. | Strain | Sex | Age | Physical Activity | Stress Model | Main Outcome | Reference |
|---|---|---|---|---|---|---|---|
| 1 | C57/B16J | female | 8 weeks | voluntary wheel running | restraint stress | Enhanced expression of proapoptotic Bax protein in the mouse cortex and microglia/macrophage expression of Cox-2 in the hippocampus | [116] |
| 2 | Wistar | male | 45–48 days | treadmill | sleep deprivation | increased protein expression of GLO1 and GSR1 in the amygdala, cortex, and hippocampus; ameliorated anxiety-like behaviour | [115] |
| 3 | Wistar | male | 3 weeks | treadmill | chronic stress restraint and forced swim stress | improved aversive memory and oxidative status changes in the amygdala and hippocampus, Na+, K+-ATPase activity in the hippocampus | [117] |
4.6. Galanin System in Locus Coeruleus
4.7. RNA Methylation
| No. | Strain | Sex | Age | Physical Activity | Stress Model | Main Outcome | Reference |
|---|---|---|---|---|---|---|---|
| 1 | C57BL/6J | male and female | 3–6 weeks | running wheels | foot shock | increased galanin expression in noradrenergic neurons | [129] |
| 2 | C57BL/6 | male | 5–7 weeks | treadmill | restraint stress | improved brain RNA m6A methylation | [139] |
| 3 | Sprague–Dawley | male | 7 weeks | voluntary wheel running | foot shock | increased galanin levels in the LC | [127] |
5. Limitation of the Study
6. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McEwen, B.S. Central effects of stress hormones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. Eur. J. Pharmacol. 2008, 583, 174–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar]
- Holsboer, F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000, 23, 477–501. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.P.; Ostrander, M.M.; Mueller, N.; Figueiredo, H. Limbic system mechanisms of stress regulation: Hypothalamo-pituitary-adrenocortical axis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 1201–1213. [Google Scholar] [CrossRef] [PubMed]
- Adlard, P.A.; Cotman, C.W. Voluntary exercise protects against stress-induced decreases in brain-derived neurotrophic factor protein expression. Neuroscience 2004, 124, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, P.J.; Self, R.L.; Hensley, A.K.; Little, H.J.; Littleton, J.M.; Prendergast, M.A. A 24 h corticosterone exposure exacerbates excitotoxic insult in rat hippocampal slice cultures independently of glucocorticoid receptor activation or protein synthesis. Brain Res. 2006, 1082, 165–172. [Google Scholar] [CrossRef]
- Goodman, Y.; Bruce, A.J.; Cheng, B.; Mattson, M.P. Estrogens attenuate and corticosterone exacerbates excitotoxicity, oxidative injury, and amyloid beta-peptide toxicity in hippocampal neurons. J. Neurochem. 1996, 66, 1836–1844. [Google Scholar] [CrossRef] [PubMed]
- Flügge, G.; Van Kampen, M.; Mijnster, M.J. Perturbations in brain monoamine systems during stress. Cell Tissue Res. 2004, 315, 1–14. [Google Scholar] [CrossRef]
- Watt, M.J.; Burke, A.R.; Renner, K.J.; Forster, G.L. Adolescent male rats exposed to social defeat exhibit altered anxiety behavior and limbic monoamines as adults. Behav. Neurosci. 2009, 123, 564–576. [Google Scholar] [CrossRef] [Green Version]
- Christoffel, D.J.; Golden, S.A.; Russo, S.J. Structural and synaptic plasticity in stress-related disorders. Rev. Neurosci. 2011, 22, 535–549. [Google Scholar] [CrossRef] [Green Version]
- Lex, C.; Bäzner, E.; Meyer, T.D. Does stress play a significant role in bipolar disorder? A meta-analysis. J. Affect. Disord. 2017, 208, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Jeong, J.; Kwak, Y.; Park, S.K. Depression research: Where are we now? Mol. Brain. 2010, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arida, R.M.; Teixeira-Machado, L. The Contribution of Physical Exercise to Brain Resilience. Front. Behav. Neurosci. 2021, 14, 626769. [Google Scholar] [CrossRef] [PubMed]
- Knapman, A.; Heinzmann, J.M.; Hellweg, R.; Holsboer, F.; Landgraf, R.; Touma, C. Increased stress reactivity is associated with cognitive deficits and decreased hippocampal brain-derived neurotrophic factor in a mouse model of affective disorders. J. Psychiatr. Res. 2010, 44, 566–575. [Google Scholar] [CrossRef]
- Wang, W.; Liu, W.; Duan, D.; Bai, H.; Wang, Z.; Xing, Y. Chronic social defeat stress mouse model: Current view on its behavioral deficits and modifications. Behav. Neurosci. 2021, 135, 326–335. [Google Scholar] [CrossRef]
- Parul Mishra, A.; Singh, S.; Singh, S.; Tiwari, V.; Chaturvedi, S.; Wahajuddin, M.; Palit, G.; Shukla, S. Chronic unpredictable stress negatively regulates hippocampal neurogenesis and promote anxious depression-like behavior via upregulating apoptosis and inflammatory signals in adult rats. Brain Res. Bull. 2021, 172, 164–179. [Google Scholar] [CrossRef]
- Bakhtiari-Dovvombaygi, H.; Izadi, S.; Zare Moghaddam, M.; Hashemzehi, M.; Hosseini, M.; Azhdari-Zarmehri, H.; Dinpanah, H.; Beheshti, F. Beneficial effects of vitamin D on anxiety and depression-like behaviors induced by unpredictable chronic mild stress by suppression of brain oxidative stress and neuroinflammation in rats. Naunyn Schmiedebergs Arch. Pharmacol. 2021, 394, 655–667. [Google Scholar] [CrossRef]
- Koprdova, R.; Bögi, E.; Belovičová, K.; Sedláčková, N.; Okuliarová, M.; Ujházy, E.; Mach, M. Chronic unpredictable mild stress paradigm in male Wistar rats: Effect on anxiety- and depressive-like behavior. Neuro Endocrinol. Lett. 2016, 37, 103–110. [Google Scholar]
- Krishnan, V.; Han, M.H.; Graham, D.L.; Berton, O.; Renthal, W.; Russo, S.J.; Laplant, Q.; Graham, A.; Lutter, M.; Lagace, D.C.; et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 2007, 131, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; de Boer, S.F.; Coppens, C.M.; Buwalda, B. Neuroendocrinology of coping styles: Towards understanding the biology of individual variation. Front. Neuroendocrinol. 2010, 31, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Bowen, M.T.; Dass, S.A.; Booth, J.; Suraev, A.; Vyas, A.; McGregor, I.S. Active coping toward predatory stress is associated with lower corticosterone and progesterone plasma levels and decreased methylation in the medial amygdala vasopressin system. Horm. Behav. 2014, 66, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, Z.X.; Qu, Y.S.; Li, L.F.; Wang, L.M.; Yuan, W.; Hou, W.J.; Zhu, Y.Q.; Cai, W.Q. Different baseline physical activity predicts susceptibility and resilience to chronic social defeat stress in mice: Involvement of dopamine neurons. Eur. Neuropsychopharmacol. 2021, 45, 15–28. [Google Scholar] [CrossRef]
- Vanderheyden, W.M.; Kehoe, M.; Vanini, G.; Britton, S.L.; Koch, L.G. Rat models for low and high adaptive response to exercise differ for stress-related memory and anxiety. Physiol. Rep. 2021, 9, 14716. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.; Shivakumar, G. Effects of exercise and physical activity on anxiety. Front. Psychiatry 2013, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Wegner, M.; Helmich, I.; Machado, S.; Nardi, A.E.; Arias-Carrion, O.; Budde, H. Effects of exercise on anxiety and depression disorders: Review of meta- analyses and neurobiological mechanisms. CNS Neurol. Disord. Drug Targets 2014, 13, 1002–1014. [Google Scholar] [CrossRef]
- Erickson, K.I.; Gildengers, A.G.; Butters, M.A. Physical activity and brain plasticity in late adulthood. Dialogues Clin. Neurosci. 2013, 15, 99–108. [Google Scholar] [CrossRef]
- Mattson, M.P. Energy intake and exercise as determinants of brain health and vulnerability to injury and disease. Cell Metab. 2012, 16, 706–722. [Google Scholar] [CrossRef] [Green Version]
- Duman, C.H.; Schlesinger, L.; Russell, D.S.; Duman, R.S. Voluntary exercise produces antidepressant and anxiolytic behavioral effects in mice. Brain Res. 2008, 1199, 148–158. [Google Scholar] [CrossRef] [Green Version]
- Phillips, C.; Fahimi, A. Immune and Neuroprotective Effects of Physical Activity on the Brain in Depression. Front. Neurosci. 2018, 12, 498. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Huang, Y.; Zhang, Y.; Huang, H.; Hong, S.; Liu, T. Impacts of exercise interventions on different diseases and organ functions in mice. J. Sport Health Sci. 2020, 9, 53–73. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zheng, Y.; Wei, J.A.; Ouyang, H.; Huang, X.; Zhang, F.; Lai, C.S.W.; Ren, C.; So, K.F.; Zhang, L. Exercise training improves motor skill learning via selective activation of mTOR. Sci. Adv. 2019, 5, 1888. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, L.; Tan, M.; Lai, C.S.; Li, A.; Ren, C.; So, K.F. Treadmill exercise suppressed stress-induced dendritic spine elimination in mouse barrel cortex and improved working memory via BDNF/TrkB pathway. Transl. Psychiatry 2017, 7, 1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleshner, M.; Maier, S.F.; Lyons, D.M.; Raskind, M.A. The neurobiology of the stress-resistant brain. Stress 2011, 14, 498–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boschloo, L.; Reeuwijk, K.G.; Schoevers, R.A.; Penninx, B.W.J.H. The impact of lifestyle factors on the 2-year course of depressive and/or anxiety disorders. J. Affect. Disord. 2014, 159, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Hafez, S.; Eid, Z.; Alabasi, S.; Darwiche, Y.; Channaoui, S.; Hess, D.C. Mechanisms of Preconditioning Exercise-Induced Neurovascular Protection in Stroke. J. Stroke 2021, 23, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Deslandes, A.; Moraes, H.; Ferreira, C.; Veiga, H.; Silveira, H.; Mouta, R.; Pompeu, F.A.; Coutinho, E.S.; Laks, J. Exercise and mental health: Many reasons to move. Neuropsychobiology 2009, 59, 191–198. [Google Scholar] [CrossRef]
- Dean, J.; Keshavan, M. The neurobiology of depression: An integrated view. Asian J. Psychiatr. 2017, 27, 101–111. [Google Scholar] [CrossRef]
- Salmon, P. Effects of physical exercise on anxiety, depression, and sensitivity to stress: A unifying theory. Clin. Psychol. Rev. 2001, 21, 33–61. [Google Scholar] [CrossRef]
- Mul, J.D. Voluntary exercise and depression-like behavior in rodents: Are we running in the right direction? J. Mol. Endocrinol. 2018, 60, 77–95. [Google Scholar] [CrossRef] [Green Version]
- Matta Mello Portugal, E.; Cevada, T.; Sobral Monteiro-Junior, R.; Teixeira Guimarães, T.; da Cruz Rubini, E.; Lattari, E.; Blois, C.; Camaz Deslandes, A. Neuroscience of exercise: From neurobiology mechanisms to mental health. Neuropsychobiology 2013, 68, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartsch, T.; Wulff, P. The hippocampus in aging and disease: From plasticity to vulnerability. Neuroscience 2015, 309, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Bou Diab, Z.; Nabha, S.; Fares, Y. Neurogenesis in the adult hippocampus: History, regulation, and prospective roles. Int. J. Neurosci. 2019, 129, 598–611. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Flügge, G. Stress, glucocorticoids and structural plasticity of the hippocampus. Neurosci. Biobehav. Rev. 1998, 23, 295–300. [Google Scholar] [CrossRef]
- Woolley, C.S.; Gould, E.; McEwen, S. Exposure to excess glucocorticoids alters dendritic morphology of adult hippocampal pyramidal neurons. Brain Res. 1990, 531, 225–231. [Google Scholar] [CrossRef]
- Pavlides, C.; Nivón, L.G.; McEwen, B.S. Effects of chronic stress on hippocampal long-term potentiation. Hippocampus 2002, 12, 245–257. [Google Scholar] [CrossRef]
- Cassilhas, R.C.; Tufik, S.; de Mello, M.T. Physical exercise, neuroplasticity, spatial learning and memory. Cell. Mol. Life Sci. 2016, 73, 975–983. [Google Scholar] [CrossRef]
- van Praag, H.; Kempermann, G.; Gage, F.H. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 1999, 2, 266–270. [Google Scholar] [CrossRef]
- van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [Green Version]
- Stranahan, A.M.; Khalil, D.; Gould, E. Running induces widespread structural alterations in the hippocampus and entorhinal cortex. Hippocampus 2007, 17, 1017–1022. [Google Scholar] [CrossRef] [Green Version]
- Abbink, M.R.; Naninck, E.F.G.; Lucassen, P.J.; Korosi, A. Early-life stress diminishes the increase in neurogenesis after exercise in adult female mice. Hippocampus 2017, 27, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Blackmore, D.G.; Steyn, F.J.; Carlisle, A.; O’Keeffe, I.; Vien, K.Y.; Zhou, X.; Leiter, O.; Jhaveri, D.; Vukovic, J.; Waters, M.J.; et al. An exercise “sweet spot” reverses cognitive deficits of aging by growth-hormone-induced neurogenesis. iScience 2021, 24, 103275. [Google Scholar] [CrossRef] [PubMed]
- Head, D.; Singh, T.; Bugg, J.M. The moderating role of exercise on stress-related effects on the hippocampus and memory in later adulthood. Neuropsychology 2012, 26, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patki, G.; Solanki, N.; Atrooz, F.; Ansari, A.; Allam, F.; Jannise, B.; Maturi, J.; Salim, S. Novel mechanistic insights into treadmill exercise based rescue of social defeat-induced anxiety-like behavior and memory impairment in rats. Physiol. Behav. 2014, 130, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Patki, G.; Li, L.; Allam, F.; Solanki, N.; Dao, A.T.; Alkadhi, K.; Salim, S. Moderate treadmill exercise rescues anxiety and depression-like behavior as well as memory impairment in a rat model of posttraumatic stress disorder. Physiol. Behav. 2014, 130, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.; Christ, C.C.; Cahill, M.M.; Aldrich, S.J.; Taylor-Yeremeeva, E. Voluntary exercise or systemic propranolol ameliorates stress-related maladaptive behaviors in female rats. Physiol. Behav. 2019, 198, 120–133. [Google Scholar] [CrossRef]
- Holmes, P.V. Trophic Mechanisms for Exercise-Induced Stress Resilience: Potential Role of Interactions between BDNF and Galanin. Front. Psychiatry 2014, 5, 90. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, G.; Wu, Z.; Xie, Y.; Zhou, L.; Xiao, L.; Wang, H. Swimming exercise reduces the vulnerability to stress and contributes to the AKT/GSK3β/CRMP2 pathway and microtubule dynamics mediated protective effects on neuroplasticity in male C57BL/6 mice. Pharmacol. Biochem. Behav. 2021, 211, 173285. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Cohen, H.; Ostfeld, I.; Kaplan, Z.; Zohar, J.; Cohen, H. Exercise Maintains Dendritic Complexity in an Animal Model of Posttraumatic Stress Disorder. Med. Sci. Sports Exerc. 2016, 48, 2487–2494. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.S.; Kim, M.Y.; Leem, Y.H. Hippocampal neuronal death induced by kainic acid and restraint stress is suppressed by exercise. Neuroscience 2011, 194, 291–301. [Google Scholar] [CrossRef]
- Herold, S.; Jagasia, R.; Merz, K.; Wassmer, K.; Lie, D.C. CREB signalling regulates early survival, neuronal gene expression and morphological development in adult subventricular zone neurogenesis. Mol. Cell. Neurosci. 2011, 46, 79–88. [Google Scholar] [CrossRef]
- Chaudhury, D.; Walsh, J.J.; Friedman, A.K.; Juarez, B.; Ku, S.M.; Koo, J.W.; Ferguson, D.; Tsai, H.C.; Pomeranz, L.; Christoffel, D.J.; et al. Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nature 2013, 493, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.L.; Covington, H.E.; Friedman, A.K.; Wilkinson, M.B.; Walsh, J.J.; Cooper, D.C.; Nestler, E.J.; Han, M.H. Mesolimbic dopamine neurons in the brain reward circuit mediate susceptibility to social defeat and antidepressant action. J. Neurosci. 2010, 30, 16453–16458. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, B.N.; Foley, T.E.; Le, T.V.; Strong, P.V.; Loughridge, A.B.; Day, H.E.; Fleshner, M. Long-term voluntary wheel running is rewarding and produces plasticity in the mesolimbic reward pathway. Behav. Brain Res. 2011, 217, 354–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastioli, G.; Arnold, J.C.; Mancini, M.; Mar, A.C.; Gamallo-Lana, B.; Saadipour, K.; Chao, M.V.; Rice, M.E. Voluntary Exercise Boosts Striatal Dopamine Release: Evidence for the Necessary and Sufficient Role of BDNF. J. Neurosci. 2022, 42, 4725–4736. [Google Scholar] [CrossRef] [PubMed]
- Mul, J.D.; Soto, M.; Cahill, M.E.; Ryan, R.E.; Takahashi, H.; So, K.; Zheng, J.; Croote, D.E.; Hirshman, M.F.; la Fleur, S.E.; et al. Voluntary wheel running promotes resilience to chronic social defeat stress in mice: A role for nucleus accumbens ΔFosB. Neuropsychopharmacology 2018, 43, 1934–1942. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. ∆FosB: A transcriptional regulator of stress and antidepressant responses. Eur. J. Pharmacol. 2015, 753, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Werme, M.; Messer, C.; Olson, L.; Gilden, L.; Thorén, P.; Nestler, E.J.; Brené, S. Delta FosB regulates wheel running. J. Neurosci. 2002, 22, 8133–8138. [Google Scholar] [CrossRef] [Green Version]
- Hendriksen, H.; Prins, J.; Olivier, B.; Oosting, R.S. Environmental enrichment induces behavioral recovery and enhanced hippocampal cell proliferation in an antidepressant-resistant animal model for PTSD. PLoS ONE 2010, 5, 11943. [Google Scholar] [CrossRef] [Green Version]
- Tanner, M.K.; Fallon, I.P.; Baratta, M.V.; Greenwood, B.N. Voluntary exercise enables stress resistance in females. Behav. Brain Res. 2019, 369, 111923. [Google Scholar] [CrossRef]
- Greenwood, B.N.; Foley, T.E.; Day, H.E.; Campisi, J.; Hammack, S.H.; Campeau, S.; Maier, S.F.; Fleshner, M. Freewheel running prevents learned helplessness/behavioral depression: Role of dorsal raphe serotonergic neurons. J. Neurosci. 2003, 23, 2889–2898. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, B.N.; Foley, T.E.; Burhans, D.; Maier, S.F.; Fleshner, M. The consequences of uncontrollable stress are sensitive to duration of prior wheel running. Brain Res. 2005, 1033, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Zou, H.W.; Jing, X.Y.; Liu, Y.J.; Li, L.F. GABA(B) receptors within the lateral habenula modulate stress resilience and vulnerability in mice. Physiol. Behav. 2021, 230, 113311. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.W.; Li, Z.L.; Jing, X.Y.; Wang, Y.; Liu, Y.J.; Li, L.F. The GABA(B1) receptor within the infralimbic cortex is implicated in stress resilience and vulnerability in mice. Behav. Brain Res. 2021, 406, 113240. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, T.G.; Wable, G.S.; Chen, Y.W.; Tateyama, K.; Yu, I.; Wang, J.Y.; Reyes, A.D.; Aoki, C. Voluntary Wheel Running Exercise Evoked by Food-Restriction Stress Exacerbates Weight Loss of Adolescent Female Rats But Also Promotes Resilience by Enhancing GABAergic Inhibition of Pyramidal Neurons in the Dorsal Hippocampus. Cereb. Cortex. 2019, 29, 4035–4049. [Google Scholar] [CrossRef] [PubMed]
- Nowacka, M.; Obuchowicz, E. BDNF and VEGF in the pathogenesis of stress-induced affective diseases: An insight from experimental studies. Pharmacol. Rep. 2013, 65, 535–546. [Google Scholar] [CrossRef]
- McAllister, A.K. Neurotrophins and neuronal differentiation in the central nervous system. Cell. Mol. Life Sci. 2001, 58, 1054–1060. [Google Scholar] [CrossRef]
- Zhang, J.; Rong, P.; Zhang, L.; He, H.; Zhou, T.; Fan, Y.; Mo, L.; Zhao, Q.; Han, Y.; Li, S.; et al. IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis. Sci. Adv. 2021, 7, 9888. [Google Scholar] [CrossRef]
- Lehmann, M.L.; Herkenham, M. Environmental enrichment confers stress resiliency to social defeat through an infralimbic cortex-dependent neuroanatomical pathway. J. Neurosci. 2011, 31, 6159–6173. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.A.; Makino, S.; Kvetnansky, R.; Post, R.M. Stress and glucocorticoids affect the expression of brain-derived neurotrophic factor and neurotrophin-3 mRNAs in the hippocampus. J. Neurosci. 1995, 15, 1768–1777. [Google Scholar] [CrossRef] [Green Version]
- Ueyamaa, T.; Kawaia, Y.; Nemotob, K.; Sekimotob, M.; Tonéa, E.; Senbaa, E. Immobilization stress reduced the expression of neurotrophins and their receptors in the rat brain. Neurosci. Res. 1997, 28, 103–110. [Google Scholar] [CrossRef]
- Nibuya, M.; Takahashi, M.; Russell, D.S.; Duman, R.S. Repeated stress increases catalytic TrkB mRNA in rat hippocampus. Neurosci. Lett. 1999, 267, 81–84. [Google Scholar] [CrossRef]
- Oliffa, H.; Berchtoldab, N.C.; Isackson, P.; Cotman, C.W. Exercise-induced regulation of brain-derived neurotrophic factor (BDNF) transcripts in the rat hippocampus. Brain Res. Mol. 1998, 68, 147–153. [Google Scholar] [CrossRef]
- Venezia, A.C.; Hyer, M.M.; Glasper, E.R.; Roth, S.M.; Quinlan, E.M. Acute forced exercise increases Bdnf IV mRNA and reduces exploratory behavior in C57BL/6J mice. Genes Brain Behav. 2020, 19, 12617. [Google Scholar] [CrossRef]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Abou Haidar, E.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. Elife 2016, 5, 15092. [Google Scholar] [CrossRef]
- Wu, Y.; Deng, F.; Wang, J.; Liu, Y.; Zhou, W.; Qu, L.; Cheng, M. Intensity-dependent effects of consecutive treadmill exercise on spatial learning and memory through the p-CREB/BDNF/NMDAR signaling in hippocampus. Behav. Brain Res. 2020, 386, 112599. [Google Scholar] [CrossRef]
- Choi, S.H.; Bylykbashi, E.; Chatila, Z.K.; Lee, S.W.; Pulli, B.; Clemenson, G.D.; Kim, E.; Rompala, A.; Oram, M.K.; Asselin, C.; et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 2018, 361, 8821. [Google Scholar] [CrossRef] [Green Version]
- Naghshvarian, M.; Zarrindast, M.R.; Mehr, S.E.; Ommati, M.M.; Sajjadi, S.F. Effect of exercise and morphine on psychological and physical dependencies, BDNF and TrkB gene expression in rat’s hippocampus. Pak. J. Med. Sci. 2017, 33, 603–609. [Google Scholar] [CrossRef]
- Walsh, J.J.; Tschakovsky, M.E. Exercise and circulating BDNF: Mechanisms of release and implications for the design of exercise interventions. Appl. Physiol. Nutr. Metab. 2018, 43, 1095–1104. [Google Scholar] [CrossRef]
- Trombetta, I.C.; DeMoura, J.R.; Alves, C.R.; Carbonari-Brito, R.; Cepeda, F.X.; Lemos, J.R. Serum Levels of BDNF in Cardiovascular Protection and in Response to Exercise. Arq. Bras. Cardiol. 2020, 115, 263–269. [Google Scholar] [CrossRef]
- Elsner, V.R.; Dorneles, G.P.; Santos, M.A.; da Silva, I.M.; Romão, P.; Peres, A. Exercise-induced BDNF production by mononuclear cells of sedentary and physically active obese men. J. Sports Med. Phys Fitness 2020, 60, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.M.; Leem, Y.H. Chronic stress-induced memory deficits are reversed by regular exercise via AMPK-mediated BDNF induction. Neuroscience 2016, 324, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xue, X.; Xia, J.; Liu, J.; Qi, Z. Swimming exercise reverses CUMS-induced changes in depression-like behaviors and hippocampal plasticity-related proteins. J. Affect. Disord. 2018, 227, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Ieraci, A.; Mallei, A.; Musazzi, L.; Popoli, M. Physical exercise and acute restraint stress differentially modulate hippocampal brain-derived neurotrophic factor transcripts and epigenetic mechanisms in mice. Hippocampus 2015, 25, 1380–1392. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.R.; Ostfeld, I.; Kaplan, Z.; Zohar, J.; Cohen, H. Exercise Enhances the Behavioral Responses to Acute Stress in an Animal Model of PTSD. Med. Sci. Sports Exerc. 2015, 47, 2043–2052. [Google Scholar] [CrossRef] [Green Version]
- Nasrallah, P.; Haidar, E.A.; Stephan, J.S.; El Hayek, L.; Karnib, N.; Khalifeh, M.; Barmo, N.; Jabre, V.; Houbeika, R.; Ghanem, A.; et al. Branched-chain amino acids mediate resilience to chronic social defeat stress by activating BDNF/TRKB signaling. Neurobiol. Stress. 2019, 11, 100170. [Google Scholar] [CrossRef]
- Yau, S.Y.; Li, A.; Zhang, E.D.; Christie, B.R.; Xu, A.; Lee, T.; So, K.F. Sustained running in rats administered corticosterone prevents the development of depressive behaviors and enhances hippocampal neurogenesis and synaptic plasticity without increasing neurotrophic factor levels. Cell Transplant. 2014, 23, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Grunbaum-Novak, N.; Taler, M.; Gil-Ad, I.; Weizman, A.; Cohen, H.; Weizman, R. Relationship between antidepressants and IGF-1 system in the brain: Possible role in cognition. Eur. Neuropsychopharmacol. 2008, 18, 431–438. [Google Scholar] [CrossRef]
- Munive, V.; Santi, A.; Torres-Aleman, I. A Concerted Action Of Estradiol And Insulin Like Growth Factor I Underlies Sex Differences In Mood Regulation By Exercise. Sci. Rep. 2016, 6, 25969. [Google Scholar] [CrossRef] [Green Version]
- Kandel, E.R. The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain 2012, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Ding, Q.; Zhang, M.; Moon, C.; Wang, H. Forebrain overexpression of type 1 adenylyl cyclase promotes molecular stability and behavioral resilience to physical stress. Neurobiol. Stress 2020, 13, 100237. [Google Scholar] [CrossRef] [PubMed]
- Mirjalili, R.; Shokouh, E.; Dehkordi, N.S.; Afsari, R.; Shafia, S.; Rashidy-Pour, A. Prior short-term exercise prevents behavioral and biochemical abnormalities induced by single prolonged stress in a rat model of posttraumatic stress disorder. Behav. Brain Res. 2022, 428, 113864. [Google Scholar] [CrossRef] [PubMed]
- Droste, S.K.; Gesing, A.; Ulbricht, S.; Müller, M.B.; Linthorst, A.C.; Reul, J.M. Effects of long-term voluntary exercise on the mouse hypothalamic-pituitary-adrenocortical axis. Endocrinology 2003, 144, 3012–3023. [Google Scholar] [CrossRef] [PubMed]
- Campeau, S.; Nyhuis, T.J.; Sasse, S.K.; Kryskow, E.M.; Herlihy, L.; Masini, C.V.; Babb, J.A.; Greenwood, B.N.; Fleshner, M.; Day, H.E.W. Hypothalamic pituitary adrenal axis responses to low-intensity stressors are reduced after voluntary wheel running in rats. J. Neuroendocrinol. 2010, 22, 872–888. [Google Scholar]
- Sasse, S.K.; Greenwood, B.N.; Masini, C.V.; Nyhuis, T.J.; Fleshner, M.; Day, H.E.; Campeau, S. Chronic voluntary wheel running facilitates corticosterone response habituation to repeated audiogenic stress exposure in male rats. Stress 2008, 11, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Yanagita, S.; Amemiya, S.; Suzuki, S.; Kita, I. Effects of spontaneous and forced running on activation of hypothalamic corticotropin-releasing hormone neurons in rats. Life Sci. 2007, 80, 356–363. [Google Scholar] [CrossRef]
- Leasure, J.L.; Jones, M. Forced and voluntary exercise differentially affect brain and behavior. Neuroscience 2008, 156, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Kochi, C.; Liu, H.; Zaidi, S.; Atrooz, F.; Dantoin, P.; Salim, S. Prior treadmill exercise promotes resilience to vicarious trauma in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 77, 216–221. [Google Scholar] [CrossRef]
- Hare, B.D.; Beierle, J.A.; Toufexis, D.J.; Hammack, S.E.; Falls, W.A. Exercise-associated changes in the corticosterone response to acute restraint stress: Evidence for increased adrenal sensitivity and reduced corticosterone response duration. Neuropsychopharmacology 2014, 39, 1262–1269. [Google Scholar] [CrossRef] [Green Version]
- Dishman, R.K.; Renner, K.J.; White-Welkley, J.E.; Burke, K.A.; Bunnell, B.N. Treadmill exercise training augments brain norepinephrine response to familiar and novel stress. Brain Res. Bull. 2000, 52, 337–342. [Google Scholar] [CrossRef]
- Pan-Vazquez, A.; Rye, N.; Ameri, M.; McSparron, B.; Smallwood, G.; Bickerdyke, J.; Rathbone, A.; Dajas-Bailador, F.; Toledo-Rodriguez, M. Impact of voluntary exercise and housing conditions on hippocampal glucocorticoid receptor, miR-124 and anxiety. Mol. Brain 2015, 8, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasse, S.K.; Nyhuis, T.J.; Masini, C.V.; Day, H.E.; Campeau, S. Central gene expression changes associated with enhanced neuroendocrine and autonomic response habituation to repeated noise stress after voluntary wheel running in rats. Front. Physiol. 2013, 4, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipović, D.; Gavrilović, L.; Dronjak, S.; Radojcić, M.B. The effect of repeated physical exercise on hippocampus and brain cortex in stressed rats. Ann. N. Y. Acad. Sci. 2007, 1096, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; Leem, T.H.; Greenwood, B.N.; Hansen, M.K.; Moraska, A.; Higgins, K.; Smith, T.P.; Fleshner, M. Habitual physical activity facilitates stress-induced HSP72 induction in brain, peripheral, and immune tissues. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, 520–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollert, C.; Zagaar, M.; Hovatta, I.; Taneja, M.; Vu, A.; Dao, A.; Levine, A.; Alkadhi, K.; Salim, S. Exercise prevents sleep deprivation-associated anxiety-like behavior in rats: Potential role of oxidative stress mechanisms. Behav. Brain Res. 2011, 224, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Gerecke, K.M.; Kolobova, A.; Allen, S.; Fawer, J.L. Exercise protects against chronic restraint stress-induced oxidative stress in the cortex and hippocampus. Brain Res. 2013, 1509, 66–78. [Google Scholar] [CrossRef]
- Dos Santos, T.M.; Kolling, J.; Siebert, C.; Biasibetti, H.; Bertó, C.G.; Grun, L.K.; Dalmaz, C.; Barbé-Tuana, F.M.; Wyse, A.T. Effects of previous physical exercise to chronic stress on long-term aversive memory and oxidative stress in amygdala and hippocampus of rats. Int. J. Dev. Neurosci. 2017, 56, 58–67. [Google Scholar] [CrossRef]
- Ogren, S.O.; Razani, H.; Elvander-Tottie, E.; Kehr, J. The neuropeptide galanin as an in vivo modulator of brain 5-HT1A receptors: Possible relevance for affective disorders. Physiol. Behav. 2007, 92, 172–179. [Google Scholar] [CrossRef]
- Mills, E.G.; Izzi-Engbeaya, C.; Abbara, A.; Comninos, A.N.; Dhillo, W.S. Functions of galanin, spexin and kisspeptin in metabolism, mood and behaviour. Nat. Rev. Endocrinol. 2021, 17, 97–113. [Google Scholar] [CrossRef]
- Holmes, P.V.; Yoo, H.S.; Dishman, R.K. Voluntary exercise and clomipramine treatment elevate prepro-galanin mRNA levels in the locus coeruleus in rats. Neurosci. Lett. 2006, 408, 1–4. [Google Scholar] [CrossRef]
- Sciolino, N.R.; Dishman, R.K.; Holmes, P.V. Voluntary exercise offers anxiolytic potential and amplifies galanin gene expression in the locus coeruleus of the rat. Behav. Brain Res. 2012, 233, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Ross, B.; Sanchez-Alavez, M.; Zorrilla, E.P.; Bartfai, T. Phenotypic analysis of GalR2 knockout mice in anxiety- and depression-related behavioral tests. Neuropeptides 2008, 42, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Kramár, E.A.; Chen, L.Y.; Babayan, A.H.; Andres, A.L.; Gall, C.M.; Lynch, G.; Baram, T.Z. Impairment of synaptic plasticity by the stress mediator CRH involves selective destruction of thin dendritic spines via RhoA signaling. Mol. Psychiatry 2013, 18, 485–496. [Google Scholar] [CrossRef]
- Hobson, S.A.; Vanderplank, P.A.; Pope, R.J.; Kerr, N.C.; Wynick, D. Galanin stimulates neurite outgrowth from sensory neurons by inhibition of Cdc42 and Rho GTPases and activation of cofilin. J. Neurochem. 2013, 127, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Y.; Yang, C.; Ren, Q.; Ma, M.; Dong, C.; Hashimoto, K. Regional differences in dendritic spine density confer resilience to chronic social defeat stress. Acta Neuropsychiatr. 2018, 30, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shirayama, Y.; Zhang, J.C.; Ren, Q.; Hashimoto, K. Regional differences in brain-derived neurotrophic factor levels and dendritic spine density confer resilience to inescapable stress. Int. J. Neuropsychopharmacol. 2015, 18, pyu121. [Google Scholar] [CrossRef] [Green Version]
- Sciolino, N.R.; Smith, J.M.; Stranahan, A.M.; Freeman, K.G.; Edwards, G.L.; Weinshenker, D.; Holmes, P.V. Galanin mediates features of neural and behavioral stress resilience afforded by exercise. Neuropharmacology 2015, 89, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.S.; Groves, J.L.; Pettett, B.J.; Britton, S.L.; Koch, L.G.; Dishman, R.K.; Holmes, P.V. Locus coeruleus galanin expression is enhanced after exercise in rats selectively bred for high capacity for aerobic activity. Peptides 2010, 31, 2264–2268. [Google Scholar] [CrossRef] [Green Version]
- Tillage, R.P.; Wilson, G.E.; Liles, L.C.; Holmes, P.V.; Weinshenker, D. Chronic Environmental or Genetic Elevation of Galanin in Noradrenergic Neurons Confers Stress Resilience in Mice. J. Neurosci. 2020, 40, 7464–7474. [Google Scholar] [CrossRef]
- Chokkalla, A.K.; Mehta, S.L.; Vemuganti, R. Epitranscriptomic regulation by m6A RNA methylation in brain development and diseases. J. Cereb. Blood Flow Metab. 2020, 40, 2331–2349. [Google Scholar] [CrossRef]
- Chang, M.; Lv, H.; Zhang, W.; Ma, C.; He, X.; Zhao, S.; Zhang, Z.W.; Zeng, Y.X.; Song, S.; Niu, Y.; et al. Region-specific RNA m6A methylation represents a new layer of control in the gene regulatory network in the mouse brain. Open Biol. 2017, 7, 170166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.; Tian, G.G.; Zhang, L.; Lee, T.; Zhang, Z.; Li, J.; Sun, T. Identifying cortical specific long noncoding RNAs modified by m6A RNA methylation in mouse brains. Epigenetics 2021, 16, 1260–1276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Du, K.; Wang, J.; Nie, Y.; Lee, T.; Sun, T. Unique and Specific m6A RNA Methylation in Mouse Embryonic and Postnatal Cerebral Cortices. Genes 2020, 11, 1139. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.J.; Ringeling, F.R.; Vissers, C.; Jacob, F.; Pokrass, M.; Jimenez-Cyrus, D.; Su, Y.; Kim, N.S.; Zhu, Y.; Zheng, L.; et al. Temporal Control of Mammalian Cortical Neurogenesis by m6A Methylation. Cell 2017, 171, 877–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widagdo, J.; Zhao, Q.Y.; Kempen, M.J.; Tan, M.C.; Ratnu, V.S.; Wei, W.; Leighton, L.; Spadaro, P.A.; Edson, J.; Anggono, V.; et al. Experience-Dependent Accumulation of N6-Methyladenosine in the Prefrontal Cortex Is Associated with Memory Processes in Mice. J. Neurosci. 2016, 36, 6771–6777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathoux, J.; Henshall, D.C.; Brennan, G.P. Regulatory Mechanisms of the RNA Modification m6A and Significance in Brain Function in Health and Disease. Front. Cell. Neurosci. 2021, 15, 671932. [Google Scholar] [CrossRef]
- Engel, M.; Eggert, C.; Kaplick, P.M.; Eder, M.; Röh, S.; Tietze, L.; Namendorf, C.; Arloth, J.; Weber, P.; Rex-Haffner, M.; et al. The Role of m6A/m-RNA Methylation in Stress Response Regulation. Neuron 2018, 99, 389–403.e9. [Google Scholar] [CrossRef]
- Song, N.; Du, J.; Gao, Y.; Yang, S. Epitranscriptome of the ventral tegmental area in a deep brain-stimulated chronic unpredictable mild stress mouse model. Transl. Neurosci. 2020, 11, 402–418. [Google Scholar] [CrossRef]
- Yan, L.; Wei, J.A.; Yang, F.; Wang, M.; Wang, S.; Cheng, T.; Liu, X.; Jia, Y.; So, K.F.; Zhang, L. Physical Exercise Prevented Stress-Induced Anxiety via Improving Brain RNA Methylation. Adv. Sci. 2022, 9, 2105731. [Google Scholar] [CrossRef]
- Svensson, M.; Rosvall, P.; Boza-Serrano, A.; Andersson, E.; Lexell, J.; Deierborg, T. Forced treadmill exercise can induce stress and increase neuronal damage in a mouse model of global cerebral ischemia. Neurobiol. Stress 2016, 5, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Aas, M.; Ueland, T.; Mørch, R.H.; Laskemoen, J.F.; Lunding, S.H.; Reponen, E.J.; Cattaneo, A.; Agartz, I.; Melle, I.; Steen, N.E.; et al. Physical activity and childhood trauma experiences in patients with schizophrenia or bipolar disorders. World J. Biol. Psychiatry 2021, 22, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Voorendonk, E.M.; Sanches, S.A.; Tollenaar, M.S.; De Jongh, A.; Van Minnen, A. Augmenting PTSD treatment with physical activity: Study protocol of the APPART study (Augmentation for PTSD with Physical Activity in a Randomized Trial). Eur. J. Psychotraumatol. 2022, 13, 2016219. [Google Scholar] [CrossRef] [PubMed]
- Pérez-de la Cruz, S. Influence of an Aquatic Therapy Program on Perceived Pain, Stress, and Quality of Life in Chronic Stroke Patients: A Randomized Trial. Int. J. Environ. Res. Public Health 2020, 17, 4796. [Google Scholar] [CrossRef] [PubMed]
- Wermelinger Ávila, M.P.; Corrêa, J.C.; Lucchetti, A.L.G.; Lucchetti, G. Relationship Between Mental Health, Resilience, and Physical Activity in Older Adults: A 2-Year Longitudinal Study. J. Aging Phys. Act. 2022, 30, 73–81. [Google Scholar] [CrossRef]
- Radino, A.; Tarantino, V. Impact of physical activity on response to stress in people aged 65 and over during COVID-19 pandemic lockdown. Psychogeriatrics 2022, 22, 227–235. [Google Scholar] [CrossRef]
- To, Q.G.; Vandelanotte, C.; Cope, K.; Khalesi, S.; Williams, S.L.; Alley, S.J.; Thwaite, T.L.; Fenning, A.S.; Stanton, R. The association of resilience with depression, anxiety, stress and physical activity during the COVID-19 pandemic. BMC Public Health 2022, 22, 491. [Google Scholar] [CrossRef]
- Dobler, C.L.; Krüger, B.; Strahler, J.; Weyh, C.; Gebhardt, K.; Tello, K.; Ghofrani, H.A.; Sommer, N.; Gall, H.; Richter, M.J.; et al. Physical Activity and Mental Health of Patients with Pulmonary Hypertension during the COVID-19 Pandemic. J. Clin. Med. 2020, 9, 4023. [Google Scholar] [CrossRef]
- Neumann, R.J.; Ahrens, K.F.; Kollmann, B.; Goldbach, N.; Chmitorz, A.; Weichert, D.; Fiebach, C.J.; Wessa, M.; Kalisch, R.; Lieb, K.; et al. The impact of physical fitness on resilience to modern life stress and the mediating role of general self-efficacy. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 679–692. [Google Scholar] [CrossRef]
- Bernstein, E.E.; McNally, R.J. Exercise as a buffer against difficulties with emotion regulation: A pathway to emotional wellbeing. Behav Res. Ther. 2018, 109, 29–36. [Google Scholar] [CrossRef]
- Mata, J.; Thompson, R.J.; Gotlib, I.H. BDNF genotype moderates the relation between physical activity and depressive symptoms. Health Psychol. 2010, 29, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Greene, C.; Hanley, N.; Campbell, M. Claudin-5: Gatekeeper of neurological function. Fluids Barriers CNS 2019, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ittner, C.; Burek, M.; Störk, S.; Nagai, M.; Förster, C.Y. Increased Catecholamine Levels and Inflammatory Mediators Alter Barrier Properties of Brain Microvascular Endothelial Cells in vitro. Front. Cardiovasc. Med. 2020, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Małkiewicz, M.A.; Małecki, A.; Toborek, M.; Szarmach, A.; Winklewski, P.J. Substances of abuse and the blood brain barrier: Interactions with physical exercise. Neurosci. Biobehav. Rev. 2020, 119, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Burek, M.; Steinberg, K.; Förster, C.Y. Mechanisms of transcriptional activation of the mouse claudin-5 promoter by estrogen receptor alpha and beta. Mol. Cell. Endocrinol. 2014, 392, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Burek, M.; Arias-Loza, P.A.; Roewer, N.; Förster, C.Y. Claudin-5 as a novel estrogen target in vascular endothelium. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Burek, M.; Förster, C.Y. Cloning and characterization of the murine claudin-5 promoter. Mol. Cell. Endocrinol. 2009, 298, 19–24. [Google Scholar] [CrossRef]
- Menard, C.; Pfau, M.L.; Hodes, G.E.; Kana, V.; Wang, V.X.; Bouchard, S.; Takahashi, A.; Flanigan, M.E.; Aleyasin, H.; LeClair, K.B.; et al. Social stress induces neurovascular pathology promoting depression. Nat. Neurosci. 2017, 20, 1752–1760. [Google Scholar] [CrossRef] [Green Version]
- Dion-Albert, L.; Cadoret, A.; Doney, E.; Kaufmann, F.N.; Dudek, K.A.; Daigle, B.; Parise, L.F.; Cathomas, F.; Samba, N.; Hudson, N.; et al. Vascular and blood-brain barrier-related changes underlie stress responses and resilience in female mice and depression in human tissue. Nat. Commun. 2022, 13, 164. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowacka-Chmielewska, M.; Grabowska, K.; Grabowski, M.; Meybohm, P.; Burek, M.; Małecki, A. Running from Stress: Neurobiological Mechanisms of Exercise-Induced Stress Resilience. Int. J. Mol. Sci. 2022, 23, 13348. https://doi.org/10.3390/ijms232113348
Nowacka-Chmielewska M, Grabowska K, Grabowski M, Meybohm P, Burek M, Małecki A. Running from Stress: Neurobiological Mechanisms of Exercise-Induced Stress Resilience. International Journal of Molecular Sciences. 2022; 23(21):13348. https://doi.org/10.3390/ijms232113348
Chicago/Turabian StyleNowacka-Chmielewska, Marta, Konstancja Grabowska, Mateusz Grabowski, Patrick Meybohm, Malgorzata Burek, and Andrzej Małecki. 2022. "Running from Stress: Neurobiological Mechanisms of Exercise-Induced Stress Resilience" International Journal of Molecular Sciences 23, no. 21: 13348. https://doi.org/10.3390/ijms232113348