
Systematic Identification and Analysis of OSC Gene Family of Rosa rugosa Thunb

College of Horticulture and Landscape Architecture, Yangzhou University, Yangzhou 225009, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2022, 23(22), 13884; https://doi.org/10.3390/ijms232213884
Submission received: 17 September 2022
/
Revised: 7 November 2022
/
Accepted: 9 November 2022
/
Published: 11 November 2022
(This article belongs to the Section Molecular Genetics and Genomics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The oxidosqualene cyclase family of Rosa rugosa (RrOSC) provides a starting point for the triterpenoid pathway, which contributes to the medicinal value of the extraction of tissues of Rosa rugosa. However, the structure and function of key RrOSCs of active triterpenoids remain ambiguous. In this study, a total of 18 RrOSC members with conservative gene structures and motifs were identified based on the genome of Rosa rugosa. The RrOSCs were located on three chromosomes including two gene clusters that derived from gene replication. The phylogenetic relationship divided RrOSCs into six groups, and the RrOSCs of GI and GIV that were represented by lupeol or α-amyrin were identified as likely to include candidate genes for producing active triterpenoids. Considering the high expression or specific-tissue expression of the candidates, RrOSC1, RrOSC10, RrOSC12, and RrOSC18 were considered the key genes. RrOSC12 was identified in vitro as lupeol synthase. The results provided fundamental information and candidate genes for further illustration of the triterpenoid pathway involved in the pharmacological activities of Rosa rugosa.
1. Introduction
Sterols and triterpenes are important components of plant metabolites, and nearly 200 different skeletons of these compounds have been identified [1]. In addition to components of cell membranes and roles in plant growth and development, triterpenes perform key roles in commercial applications by biotic or abiotic function, such as preservatives, flavor modifiers, and cholesterol-lowering agents. For example, the sweetness of licorice roots is attributable to the presence of the triterpenoid saponin glycyrrhizin. Evidence of the health benefits of saponins is increasing, and their expanding application in food, cosmetics, and pharmaceutical industries is attracting commercial attention [2]. In the aspect of biochemical reaction, 2,3-Oxidosqualene is the common precursor of all the sterols and triterpenoids. The cyclization processes of 2,3-Oxidosqualene including protonation, cyclization, rearrangement, and deprotonation produce the diversiform skeletons of sterols and triterpenes [3,4,5]. During the cyclization processes, “chair-chair-chair (C-C-C)” and “chair-boat-chair (C-B-C)” configurations are present prior to cyclization into diverse triterpenes. Sterols are mainly formed through the “C-B-C” configuration, and triterpenoids are mainly formed through the “C-C-C” configuration [1].
Oxidosqualene cyclases (OSCs) are the pivotal enzyme in promoting the diversification of triterpenes, which can catalyze cyclization processes. Phillips et al., (2006) [6] divided plant OSCs into two groups based on the nature of their presumed catalytic intermediates, that is, protosteryl and dammarenyl cations, which possess distinct stereochemistry and ring configurations [6,7]. The protosteryl cation adopts the C-B-C configuration and is an intermediate leading to the formation of cycloartenol, lanosterol, parkeol, and cucurbitadienol tetracyclic triterpene skeletons (6-6-6-5). Isoarborinol is an unusual pentacyclic triterpene (6-6-6-6-5) derived from the additional D-ring expansion of the protosteryl cation based on the C-B-C configuration. Most pentacyclic triterpene skeletons, however, are derived from the dammarenyl cation by D-ring expansion to form lupeol or further E-ring expansion to form β-amyrin via the C-C-C conformation [1]. More than 150 OSC genes have been identified in plants and their catalytic function were mostly identified by heterologous expression in yeast. The Arabidopsis thaliana genome contains 13 OSC genes, including cycloartenol synthase (AtCAS1), lanosterol synthase (AtLSS1), and β-amyrin synthase (LUP4), and several OSCs with mixed functions [8,9,10,11,12]. The amyrin synthase of Eriobotrya japonica (EjAS) can catalyze the conversion of 2,3-oxidosqualene into α- and β-amyrin at a ratio of 17:3 [13]. In OSCs of Malus×domestica (MdOSCs), MdOSC1 and MdOSC3 can produce α-amyrin as the main product, with β-amyrin and lupeol as the minor products. MdOSC4 is characterized as germanicol synthase, and MdOSC5 produces lupeol as a major product [13,14]. Further, the potential of plant OSCs has been exploited in engineered yeast. MdOSC1 introduced Saccharomyces cerevisiae achieve 5.8 times the maximum α-amyrin production [14].
Rosa rugosa is a shrub of Rosaceae and native to North China, Northeast China, Japan, Korea, and Russia, and has a long history of application as a medicinal plant. Its petals and roots have been widely used to staunch bleeding [15], reduce blood lipids, treat diabetes, and as a cure for cancer or cardiovascular diseases [16,17]. In the R. rugosa root extract, the active triterpenoids Kaji ichigoside F1, rosamultin, euscaphic acid, and tormentic acid showed anti-injury and anti-inflammatory effects on animal models; among these compounds, rosamultin had a strong inhibitory effect on human immunodeficiency virus-1 protease [18,19]. These reports point to the key role that terpenoids play in the pharmacological activities of R. rugosa. However, the OSC genes of R. rugosa (RrOSCs) involved in the synthesis of pharmacological terpenoids (e.g., rosamultin) haven’t been reported. This study was aimed at discovering the potential key RrOSCs in the active terpenoids synthesis pathway of R. rugosa by systematic analysis of the RrOSC family.
2. Results
2.1. Identification and Phylogenetic Analysis of RrOSCs
A total of 18 RrOSC genes were identified, and the predicted proteins ranged from 540 to 770 amino acids in length and from 62.283 to 85.712 kDa in molecular weight (Table S1). All OSCs (except RrOSC4, which is extremely short and could belong to a pseudogene or unknown coding gene) were divided into 6 Groups (GI–GVI), which were named by common catalytic function based on the phylogenetic branches (Figure 1). OSCs in GI would produce mixed products and the main components of the mixed products were different; OSCs of GIII including RrOSC9 and RrOSC13 would be cycloartenol synthases. OSCs of GIV including RrOSC12 would be lupeol synthases. OSCs of GV including RrOSC14, RrOSC 15, and RrOSC 16 would be lanosterol synthases. No RrOSCs belonged to GII (β-amyrin synthases) or GVI (specific to monocotyledons).
2.2. The Conserved Motifs and Gene Structures of RrOSCs
The exon number of RrOSCs was in the range of 14–21. The SQHop_cyclase_C and/or SQHop_cyclase_N domains located by constant exon number (18–20 exons) indicated gene structures of RrOSCs were conserved (Figure 2b). Analysis of motifs (Figure 2a) showed that the domains of most RrOSCs composed of motifs 3-8-5-10-2 (corresponding to SQHop_cyclase_C) and motifs 6-7-4-1 (corresponding to SQHop_cyclase_N) were highly aligned. Interestingly, Group I members lack one motif5 in the N-terminal compared while other RrOSCs comprised of two motif5. And motif6 and/or motif7 were absent in SQHop_cyclase_N of Group I members RrOSC1 and RrOSC3.
2.3. Clusters and Gene Duplications of RrOSCs
The RrOSCs are distributed on three (Chr2, Chr4, and Chr7) of all seven chromosomes (Figure 3a). Most members are located on the gene clusters of Chr2 and Chr4 while only RrOSC18 is distributed on the Chr7. According to the gene duplication type (Table S3), RrOSC2/3/5/8/9/14/15 belonged to the tandem duplication type and RrOSC6/7/10/13 belonged to the proximal duplication type. According to the gene pairs of synteny analysis (Table S4), two tandemly duplicate gene pairs RrOSC2-RrOSC3, RrOSC8-RrOSC9 (Figure 3b), and proximal duplicated RrOSC6/7/10 contributed to the origin of the gene cluster on Chr2. RrOSC14-RrOSC15 pair (Figure 3b) and proximal duplicated RrOSC13 explained the origin of the gene cluster on Chr4. The segmentally duplicate events (Figure 3b) were not detected in the RrOSCs.
2.4. Expression Analysis of RrOSCs
To detect the candidate RrOSC genes of the synthetic pathways of R. rugosa active triterpenoids, we investigated the temporal and spatial expression patterns of RrOSCs in roots, flowers, and lateral branches (Figure 4). RrOSC2, RrOSC3, and RrOSC5 of GI and RrOSC14, RrOSC15, and RrOSC16 of G5 expressed in a low abundance (TPM < 1) were not discussed further. RrOSC9, RrOSC10, and RrOSC13 are expressed in all tissues and all stages in a middle or high abundance (minimum TPM > 10), indicating their constitutive expression pattern.
The highly-expressed RrOSC6, RrOSC10, and RrOSC13 in the lateral branches of R. rugosa showed two expression patterns. The expression of RrOSC6 and RrOSC13 increased from the first lateral branch to the third lateral branch. The expression of RrOSC10 was the highest, decreased from the primary lateral branch to the secondary lateral branch, and increased from the secondary lateral branch to the tertiary lateral branch. The three genes all showed the highest abundance in the third lateral branch, and only RrOSC6 specific expressed in branches.
In roots (Figure 4, TIR or OR), the expression levels of RrOSC10, RrOSC12, RrOSC13, and RrOSC18 were high (FPKM > 50), and RrOSC12 was strongly expressed in TIR but almost not at all in OR. In flowers, the expression levels of RrOSC9, RrOSC10, and RrOSC13 were higher. The expression levels of RrOSC9 and RrOSC13 were high in the early stage, decreased at the end of the flowering stage, and reached a maximum at S2 and S3. The expression of RrOSC10 was the highest, and it increased significantly at the end of the flowering stage and reached the maximum at S7. In the lateral branches and petals, more than half of the genes were not expressed or had low expression.
2.5. Functional Characterization of RrOSC12
To elucidate the function of RrOSC12, we detected its products by expression in yeast. Authentic lupeol showed a dominant peak at 17.8 min (Figure 5b). A peak with the corresponding retention time was observed in the total ion chromatogram of RrOSC12 expressed yeast, and mass spectra confirmed that it was that of lupeol (Figure 5c). These results demonstrate that RrOSC12 catalyzes 2,3-oxidosqualene to yield lupeol.
3. Discussion
The OSC gene number (18 members) of R. rugosa was more than the 14 members found in Arabidopsis and 11 members in rice. A comparison of gene structure and motifs showed that the gene structure difference of RrOSCs in the same subgroup didn’t affect their highly-conserved domains including consistent motif structure. Additionally, many RrOSCs in the same subgroup shared more similar expression patterns, such as two different subgroups of GI: RrOSC7, RrOSC11, and RrOSC17 all highly expressed in flowers or RrOSC6, RrOSC10, and RrOSC18 all highly expressed in roots (or branches).
The OSCs in the same subgroup in GI could produce mix-products with accordant main components. RrOSC1-RrOSC6, RrOSC10, and RrOSC18 were clustered with MdOSC3, EjAS, and MdOSC1 of Rosaceae plants as a subgroup. MdOSC1 yields α-amyrin, β-amyrin, and lupeol at a ratio of 85:13:2 [13], EjAS produces α-amyrin and β-amyrin at a ratio of 17:3, and MdOSC3 manufactures α-amyrin, β-amyrin, and lupeol at a ratio of 85:14:1 [20]. This subgroup was most likely a cluster of α-amyrin synthases. In addition, the tandem replication pair RrOSC2-RrOSC3 and proximal duplication genes RrOSC6, and RrOSC10 indicated that gene duplications contributed to the functional redundancy of the seven α-amyrin synthase candidates. RrOSC7, RrOSC8, RrOSC11, and RrOSC17 combined with MdOSC4 and MdOSC5 were clustered into another subgroup. The main product of MdOSC4 is germanicol, that of MdOSC5 is lupeol, and both contain the by-product β-amyrin. Thus, the four candidates are unlikely to be α-amyrin synthases. According to the structural comparison, the rosamultin of R. rugosa should be formed from α-amyrin through cytochrome P450 oxidation to introduce functional groups and finally glycosylated by glycosyltransferase. Except for the four low abundance genes, above, RrOSCs, RrOSC1, RrOSC10, and RrOSC18 were considered as the candidate genes of the rosamultin synthesis pathway.
Lupeol is an important pentacyclic triterpenoid. The applications of lupeol derivatives in the field of medicine are extensively studied because of their pharmacological activities, including antinephrotoxicity, antihepatotoxicity, and anticancer, anti-inflammatory, anti-heart disease, antiarthritis, and antidiabetes properties [21]. Lupeol was also found to be involved in the formation of epicuticular wax crystals on the stem and hypocotyl surfaces [22]. Only OSC of R. rugosa (RrOSC12) belongs to GIV, whose members would catalytic substrate to produce lupeol. The yeast expression proved its catalytic activity of lupeol. Though the lupeol synthase activity in vivo of RrOSC12 needs to be studied further, its remarkably high mRNA abundance in TR and the lupenone and betulinic acid (downstream products of lupeol) accumulation in TR (unpublished metabolome data) indicated that lupeol contributes partly to the medicinal value of R. rugosa roots. In addition, the chromosome terminal locations (near telomere) of RrOSC12 provide insight into DNA methylation, which could be the entry point for further exploring the pathway gene cluster to jointly modify the substrate (i.e., the gene cluster of RrOSC12 and corresponding downstream P450 genes).
The enormous metabolic engineering potential of OSCs is increasingly apparent, e.g., MdOSC1 [14,23], enzymes cycloartenol synthase gene of Panax ginseng and β-amyrin synthase genes of Glycyrrhiza glabra, and P. ginseng [24,25]. Based on the genetic transformation method in hairy roots of R. rugosa that has been established by our own lab, we would like to overexpress RrOSC12 in R. rugosa hairy roots to increase lupeol production. In addition, the two α-amyrin synthase candidates RrOSC10/18, which were highly expressed in roots, should be tested further for increasing the potential of α-amyrin and that of the downstream product rosamultin.
4. Materials and Methods
4.1. Plant Materials
The three-year-old cutting seedings of R. rugosa “Zi Zhi” were planted in germplasm in the resource nursery of Yangzhou University (32.391° N, 119.419° E), Yangzhou, China. Samples including roots (OR), primary lateral branch (PB), secondary lateral branch (SB), tertiary lateral branch (TB), flowers of seven stages (S1–S7), and roots (TR) of tissue culture seedlings (two-month-old micro-cutting seedings) were collected and frozen using liquid nitrogen, and then stored at −80 °C for RNA extraction (Figure 6).
4.2. Identification of RrOSCs
RrOSCs were predicted by HMMER 3.0 from the proteome of R. rugosa “Zi Zhi” (unpublished) using the hidden Markov model of the squalene-hopene cyclase N-terminal domain (PF13249) and squalene-hopene cyclase C-terminal domain (PF13243) [26]. The candidate genes with both domains were confirmed by Pfam (http://pfam.xfam.org/, accessed on 1 September 2022) and the Conserved Domain Database [27,28]. The dubious OSC genes were checked by manual inspection. The isoelectric point and molecular weight of RrOSC proteins were obtained by ProtParam tool of Extasy [29].
4.3. Phylogenetic Analyses
RrOSC proteins and 78 OSC proteins of different plant species were aligned using the MAFFT algorithm (https://mafft.cbrc.jp/alignment/server/index.html, accessed on 1 September 2022) [30]. The phylogenetic dendrogram was constructed by the neighbor-joining method of MEGA-X [31], with 1000 bootstrap replications. In addition, the p-distance model of the substitution type, pairwise deletion of Gaps/Missing data treatment, and uniform rates of rates among sites were selected for phylogenetic analysis. The 78 OSC genes of Arabidopsis thaliana, Lotus japonicus, Panax ginseng, Abies magnifica, Adiantum capillus-veneris, Avena strigose, Betula platyphylla, Costus speciosus, Cucurbita pepo, Dioscorea zingiberensis, Glycyrrhiza glabra, Kalanchoe daigremontiana, Kandelia candel, Luffa cylindrica, Oryza sativa, Pisum sativum, Polypodiodes niponica, Rhizophora stylosa, Ricinus communis, Artemisia annua, Aster sedifolius, Bruguiera gymnorrhiza, Euphorbia tirucalli, Medicago truncatula, Nigella sativa, Polygala tenuifolia, Solanum lycopersicum, Vaccaria hispanica, Olea europaea, Taraxacum officinale, Aster tataricus, Stevia rebaudiana, Malus pumila, and Eriobotrya japonica are shown in Table S2.
4.4. Gene Structures and Conserved Motifs Analysis
Conserved motifs of RrOSC proteins were predicted by the MEME suite (http://meme-suite.org/, accessed on 1 September 2022) with the default parameters and a maximum motif number of 8 [32]. The conserved motifs and gene structures with domain location of RrOSCs were illustrated by the gene structure view tool of TBtools toolkit 1.086 [33] based on the genomic DNA and mRNA sequences of the RrOSCs (Tables S5 and S6).
4.5. Chromosome Location and Synteny Analysis
According to the GFF3 file of the reference genome, RrOSC gene location was illustrated by the Gene location visualize function of the GTF/GFF tool of TBtools toolkit 1.086 [33].
Inter-species synteny analysis was conducted by reciprocal BLASTP search for potential homologous gene pairs (E < 10 − 5, top five matches) of the whole genome, and the syntenic regions were predicted by the MCScanX tool of TBtools toolkit 1.086 [34]. The gene duplication events were predicted according to syntenic regions by the ‘duplicate_gene_classifier’ program of MCScanX [34].
4.6. Expression Analysis and Functional Characterization
The RNA of the above 12 tissues (R, TR, PB, SB, TB, and S1–S7) were extracted by the MiniBEST Universal RNA Extraction Kit (TaKaRa, Kusatsu, Japan) for transcriptome libraries building and sequencing on the Illumina NovaSeq 6000 platform. The raw reads could be accessed in the SRA of NCBI (PRJNA725330) and BIG Sub of China National Center for Bioinformation (PRJCA012932). After low-quality reads filtering, clean reads were used for analysis. Based on clean reads, read counts of each gene were calculated by mapping to the genome of R. rugosa “Zi Zhi” using HISAT 2.2.1 (http://daehwankimlab.github.io/hisat2/, accessed on 1 September 2022). The sequencing quality was examined by Q30 value, percentage of clean reads, and alignment coverage (Table S7). Fragments per kilobase of exon model per million reads mapped (FPKM) of RrOSCs were calculated by normalization of read counts using a self-built R script.
RrOSC12 was cloned by the polymerase chain reaction (PCR). The coding sequence of RrOSC12 was sub-cloned to pESC-leu and transformed to yeast strain AM94. Culture extracts of the RrOSC12 expressed yeast were derivatized with trimethylsilylating agents and submitted to gas chromatography–mass spectrometry analysis.
Supplementary Materials
The supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232213884/s1.
Author Contributions
Conceptualization, L.F. and J.W.; data curation, P.W., M.X. and Y.C.; writing—Original draft, P.W. and J.W.; writing—Review & editing, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the National Natural Science Foundation of China (grant numbers 31972454, 32171861, 32002076), the National Key R&D Program of China (grant number 2018YFD1000400), the Jiangsu Provincial Natural Science Foundation (grant number SBK2020043530), the Jiangsu Provincial Natural Science Research Project of Universities (grant number 20KJB210004), and the Jiangsu Provincial Agricultural Science and Technology Independent Innovation Fund (grant numbers CX(20)3023, CX(20)3026), and supported by the high-end talent support program of Yangzhou University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Xu, R.; Fazio, G.C.; Matsuda, S.P. On the origins of triterpenoid skeletal diversity. Phytochemistry 2004, 65, 261–291. [Google Scholar] [CrossRef] [PubMed]
- Osbourn, A.; Goss, R.J.; Field, R.A. The saponins: Polar isoprenoids with important and diverse biological activities. Nat. Prod. Rep. 2011, 28, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Haralampidis, K.; Trojanowska, M.; Osbourn, A.E. Biosynthesis of triterpenoid saponins in plants. Adv. Biochem. Eng. Biotechnol. 2002, 75, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M.; Zhang, H.; Endo, A.; Shishikura, K.; Kushiro, T.; Ebizuka, Y. Two branches of the lupeol synthase gene in the molecular evolution of plant oxidosqualene cyclases. Eur. J. Biochem. 1999, 266, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Basyuni, M.; Oku, H.; Tsujimoto, E.; Kinjo, K.; Baba, S.; Takara, K. Triterpene synthases from the Okinawan mangrove tribe, Rhizophoraceae. FEBS J. 2007, 274, 5028–5042. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.R.; Rasbery, J.M.; Bartel, B.; Matsuda, S.P.T. Biosynthetic diversity in plant triterpene cyclization. Curr. Opin. Plant Biol. 2006, 9, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Duan, L.; Liu, D.; Guo, J.; Ge, S.; Dicks, J.; ÓMáille, P.; Osbourn, A.; Qi, X. Divergent evolution of oxidosqualene cyclases in plants. New Phytol. 2012, 193, 1022–1038. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Shibuya, M.; Katsube, Y.; Tsutsumi, T.; Otsuka, M.; Zhang, H.; Masuda, K.; Ebizuka, Y. A new triterpene synthase from Arabidopsis thaliana produces a tricyclic triterpene with two hydroxyl groups. Org. Lett. 2006, 8, 2835–2838. [Google Scholar] [CrossRef]
- Kushiro, T.; Shibuya, M.; Masuda, K.; Ebizuka, Y. A novel multifunctional triterpene synthase from Arabidopsis thaliana. Tetrahedron Lett. 2000, 41, 7705–7710. [Google Scholar] [CrossRef]
- Lodeiro, S.; Xiong, Q.; Wilson, W.K.; Kolesnikova, M.D.; Onak, C.S.; Matsuda, S.P.T. An oxidosqualene cyclase makes numerous products by diverse mechanisms: A challenge to prevailing concepts of triterpene biosynthesis. J. Am. Chem. Soc. 2007, 129, 11213–11222. [Google Scholar]
- Xiong, Q.; Wilson, W.K.; Matsuda, S.P. An Arabidopsis oxidosqualene cyclase catalyzes iridal skeleton formation by Grob fragmentation. Angew. Chem. Int. Ed. Engl. 2006, 45, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.J.R.; Meyer, M.M.; Matsuda, S.P.T. Arabidopsis thaliana LUP1 converts oxidosqualene to multiple triterpene alcohols and a triterpene diol. Org. Lett. 2000, 2, 2257–2259. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.M.; Legay, S.; Deleruelle, A.; Nieuwenhuizen, N.; Punter, M.; Brendolise, C.; Cooney, J.M.; Lateur, M.; Hausman, J.F.; Larondelle, Y.; et al. Multifunctional oxidosqualene cyclases and cytochrome P450 involved in the biosynthesis of apple fruit triterpenic acids. New Phytol. 2016, 211, 1279–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Chang, P.; Yu, H.; Ren, H.; Hong, D.; Li, Z.; Wang, Y.; Song, H.; Huo, Y.; Li, C. Productive Amyrin Synthases for Efficient α-Amyrin Synthesis in Engineered Saccharomyces cerevisiae. ACS Synth. Biol. 2018, 7, 2391–2402. [Google Scholar] [CrossRef] [PubMed]
- Hashidoko, Y. The phytochemistry of Rosa rugosa. Phytochemistry 1996, 43, 535–549. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, J.D.; Lee, Y.H.; Rhee, H.; Choi, Y.S. Influence of extract of Rosa rugosa roots on lipid levels in serum and liver of rats. Life Sci. 1991, 49, 947–951. [Google Scholar] [CrossRef]
- Na, J.R.; Oh, D.R.; Han, S.; Kim, Y.J.; Choi, E.; Bae, D.; Oh, D.H.; Lee, Y.H.; Kim, S.; Jun, W. Antistress Effects of Rosa rugosa Thunb. On Total Sleep Deprivation-Induced Anxiety-Like Behavior and Cognitive Dysfunction in Rat: Possible Mechanism of Action of 5-HT6 Receptor Antagonist. J. Med. Food 2016, 19, 870–881. [Google Scholar] [CrossRef]
- Jung, H.-J.; Nam, J.-H.; Choi, J.; Lee, K.-T.; Park, H.-J. 19Alpha-hydroxyursane-type triterpenoids: Antinociceptive anti-inflammatory principles of the roots of Rosa rugosa. Biol. Pharm. Bull. 2005, 28, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Park, J.C.; Kim, S.C.; Choi, M.R.; Song, S.H.; Yoo, E.J.; Kim, S.H.; Miyashiro, H.; Hattori, M. Anti-HIV protease activity from rosa family plant extracts and rosamultin from Rosa rugosa. J. Med. Food. 2005, 8, 107–109. [Google Scholar] [CrossRef]
- Liu, X.; Su, M.; Wu, S.; Li, H. Clone of Amyrin Synthase Gene Conservative District from Eriobotrya japonica ‘Jie Fang Zhong’. Subtrop. Plant Sci. 2013, 42, 1–4. (In Chinese) [Google Scholar]
- Liu, K.; Zhang, X.; Xie, L.; Deng, M.; Chen, H.; Song, J.; Long, J.; Li, X.; Luo, J. Lupeol and its derivatives as anticancer and anti-inflammatory agents: Molecular mechanisms and therapeutic efficacy. Pharm. Res. 2021, 164, 105373. [Google Scholar] [CrossRef] [PubMed]
- Guhling, O.; Hobl, B.; Yeats, T.; Jetter, R. Cloning and characterization of a lupeol synthase involved in the synthesis of epicuticular wax crystals on stem and hypocotyl surfaces of Ricinus communis. Arch. Biochem. Biophys. 2006, 448, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Rasool, A.; Liu, H.; Lv, B.; Chang, P.; Song, H.; Wang, Y.; Li, C. Engineering Saccharomyces Cerevisiae for High Yield Production of A-Amyrin Via Synergistic Remodeling of A-Amyrin Synthase and Expanding the Storage Pool. Metab. Eng. 2020, 62, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhao, S.; Zhang, X. Antisense Suppression of Cycloartenol Synthase Results in Elevated Ginsenoside Levels in Panax Ginseng Hairy Roots. Plant Mol. Biol. Rep. 2009, 27, 298–304. [Google Scholar] [CrossRef]
- Dai, Z.; Wang, B.; Liu, Y.; Shi, M.; Wang, D.; Zhang, X.; Liu, T.; Huang, L.; Zhang, X. Producing Aglycons of Ginsenosides in Bakers’ Yeast. Sci. Rep. 2014, 4, 3698. [Google Scholar] [CrossRef] [Green Version]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; C. Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. Expasy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. Mafft Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis of 95 oxidosqualene cyclases (OSCs) of different plant species. The phylogenetic dendrogram was generated using the neighbor-joining method with 1000 bootstrap replicates (numbers in branches). Groups were distinguished with different colors and OSCs of Rosa rugosa (RrOSCs) were highlighted. The information on plant species and corresponding OSCs are listed in Table S2.
Figure 1.
Phylogenetic analysis of 95 oxidosqualene cyclases (OSCs) of different plant species. The phylogenetic dendrogram was generated using the neighbor-joining method with 1000 bootstrap replicates (numbers in branches). Groups were distinguished with different colors and OSCs of Rosa rugosa (RrOSCs) were highlighted. The information on plant species and corresponding OSCs are listed in Table S2.
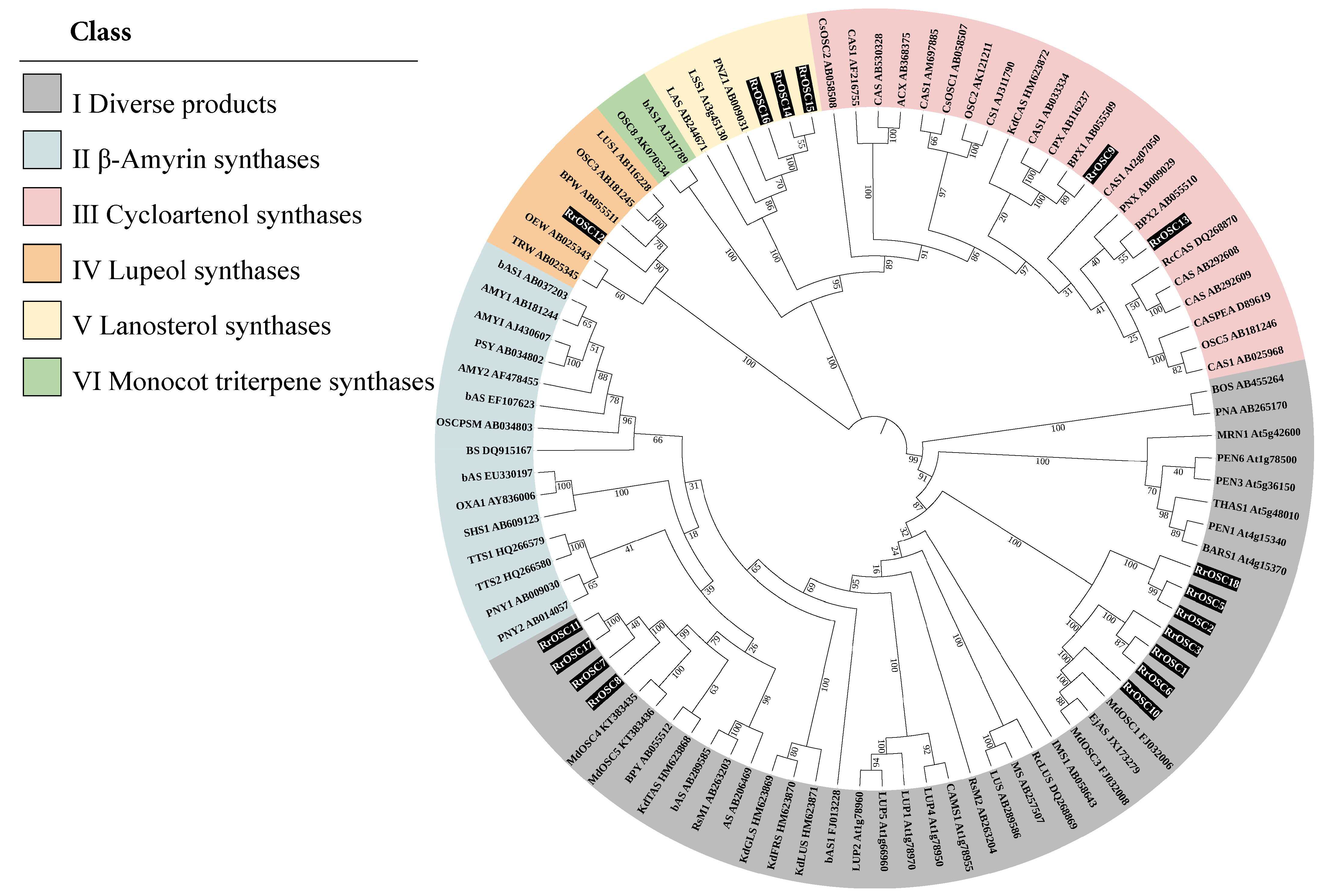
Figure 2.
Motifs (a), gene structures (b), and motif sequences (c) of RrOSCs. The coloring boxes represent motifs whose locations were labeled by the scale plate of amino acid residue (a). Exons (rectangles) separated by introns (lines) were colored yellow (coding sequence) and green (untranslated region), and the locations were labeled by the nucleotide scale plate. Domains were colored purple (SQHop_cyclase_C) and blue (SQHop_cyclase_N) (b). The top 10 significantly enriched motifs are listed as sequence logos (c).
Figure 2.
Motifs (a), gene structures (b), and motif sequences (c) of RrOSCs. The coloring boxes represent motifs whose locations were labeled by the scale plate of amino acid residue (a). Exons (rectangles) separated by introns (lines) were colored yellow (coding sequence) and green (untranslated region), and the locations were labeled by the nucleotide scale plate. Domains were colored purple (SQHop_cyclase_C) and blue (SQHop_cyclase_N) (b). The top 10 significantly enriched motifs are listed as sequence logos (c).
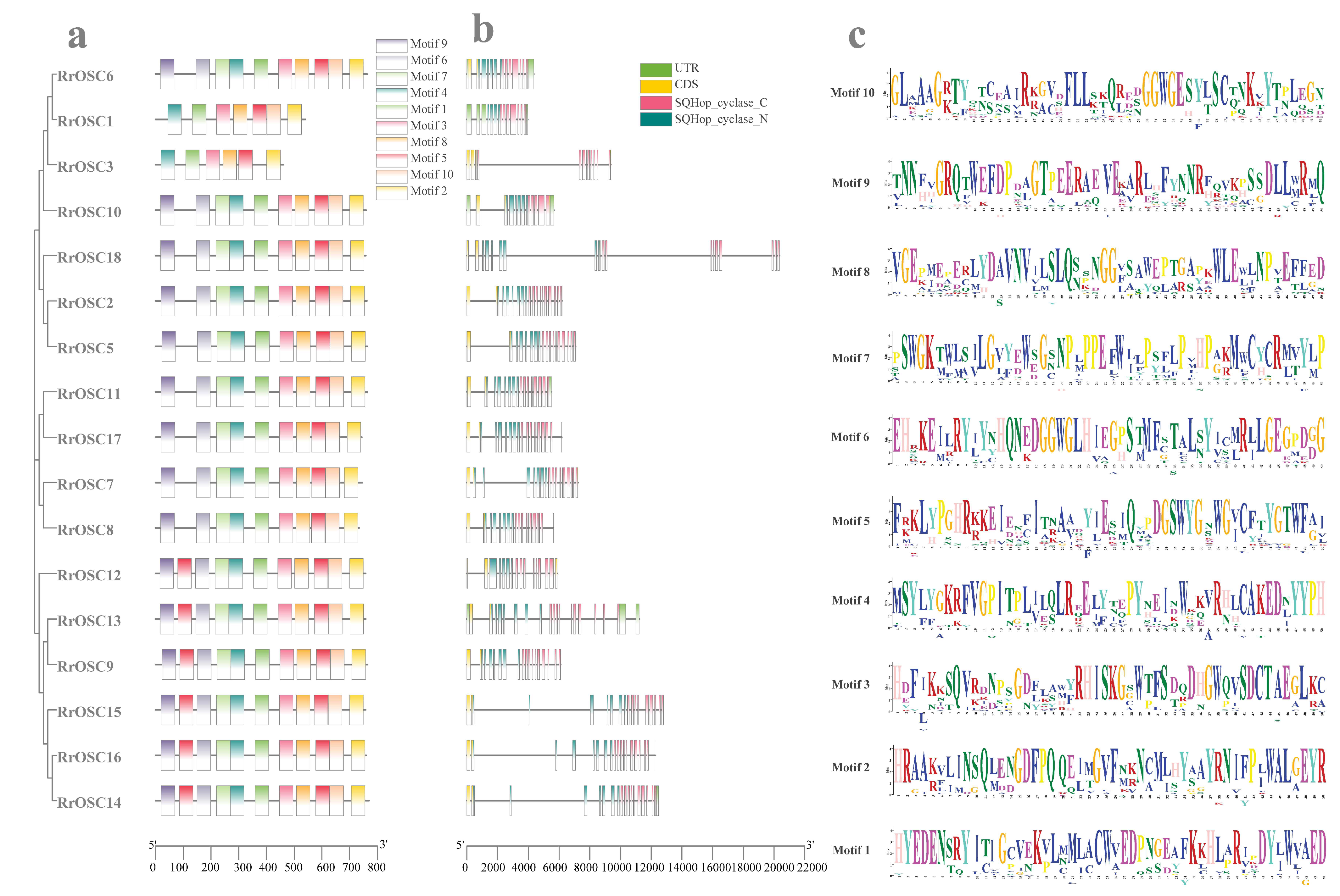
Figure 3.
Chromosomal localization (a) and tandem duplication (b) analysis of RrOSCs. Blue lines indicated the duplication gene pairs according to the synteny analysis.
Figure 3.
Chromosomal localization (a) and tandem duplication (b) analysis of RrOSCs. Blue lines indicated the duplication gene pairs according to the synteny analysis.
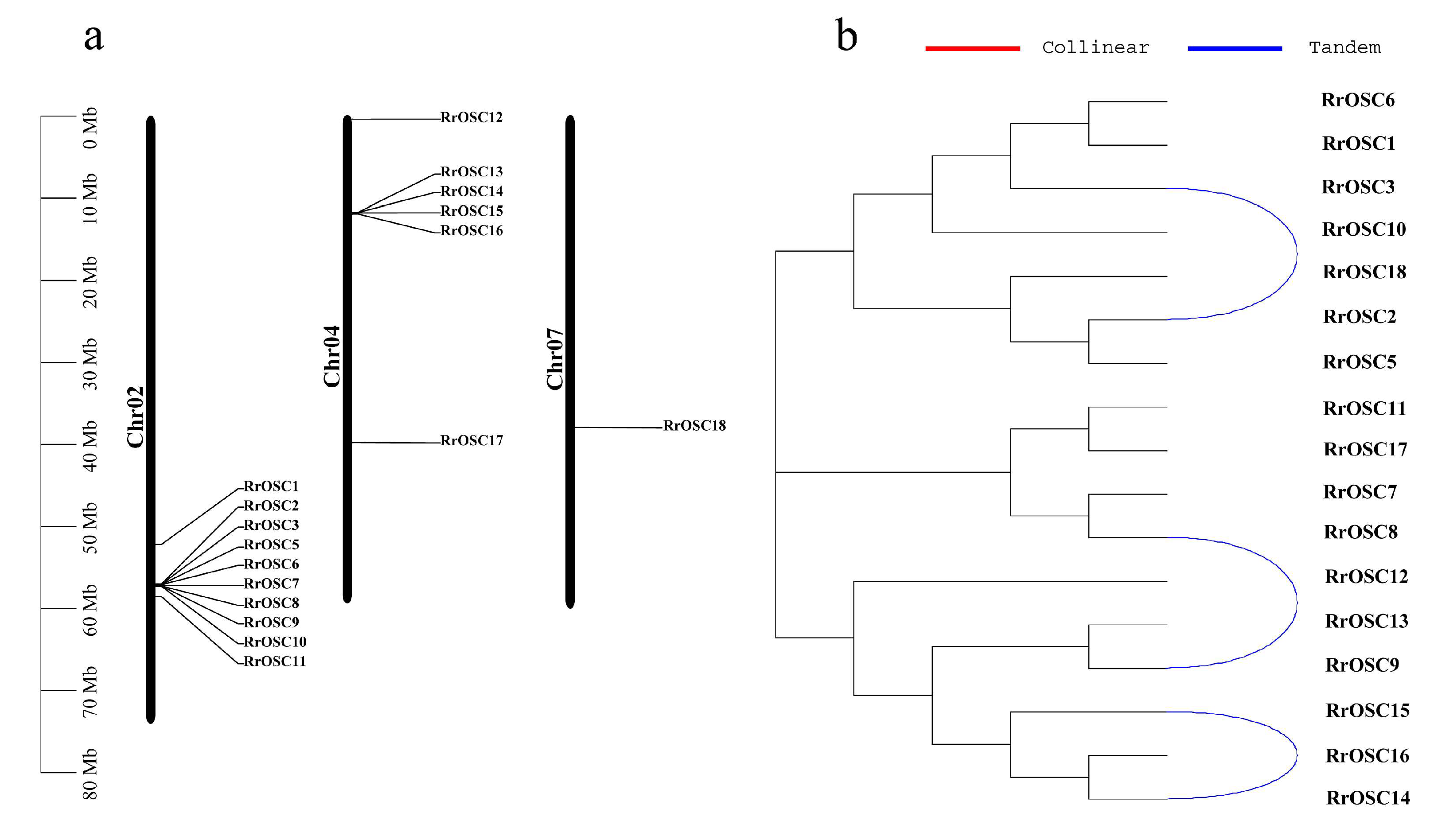
Figure 4.
Digital gene expression profiles in the roots, lateral branches, and flowers of R. rugosa “Zi Zhi”. A heat map was generated based on the normalized Log2 FPKM represented by the blue-yellow gradation. The numbers in the heat map were FPKM from RNA-seq data. The OR and TR columns represent the roots of open-air and tissue culture seedlings, respectively The columns of FB, SB, and TB represent the primary, secondary, and tertiary lateral branches, respectively. The columns of S1–S7 represent the seven flower stages, respectively.
Figure 4.
Digital gene expression profiles in the roots, lateral branches, and flowers of R. rugosa “Zi Zhi”. A heat map was generated based on the normalized Log2 FPKM represented by the blue-yellow gradation. The numbers in the heat map were FPKM from RNA-seq data. The OR and TR columns represent the roots of open-air and tissue culture seedlings, respectively The columns of FB, SB, and TB represent the primary, secondary, and tertiary lateral branches, respectively. The columns of S1–S7 represent the seven flower stages, respectively.
Figure 5.
Gas chromatography–mass spectrometry analysis of yeast extracts overexpressing the R. rugosa lupeol synthase gene. Chromatogram for the yeast with empty vector (a), transformed yeast with the RrOSC12 gene (b), and the authentic lupeol standard (c). Mass spectra in a retention time of 17.8 min for lupeol in the transformed yeast (d) and the authentic lupeol standard (e).
Figure 5.
Gas chromatography–mass spectrometry analysis of yeast extracts overexpressing the R. rugosa lupeol synthase gene. Chromatogram for the yeast with empty vector (a), transformed yeast with the RrOSC12 gene (b), and the authentic lupeol standard (c). Mass spectra in a retention time of 17.8 min for lupeol in the transformed yeast (d) and the authentic lupeol standard (e).
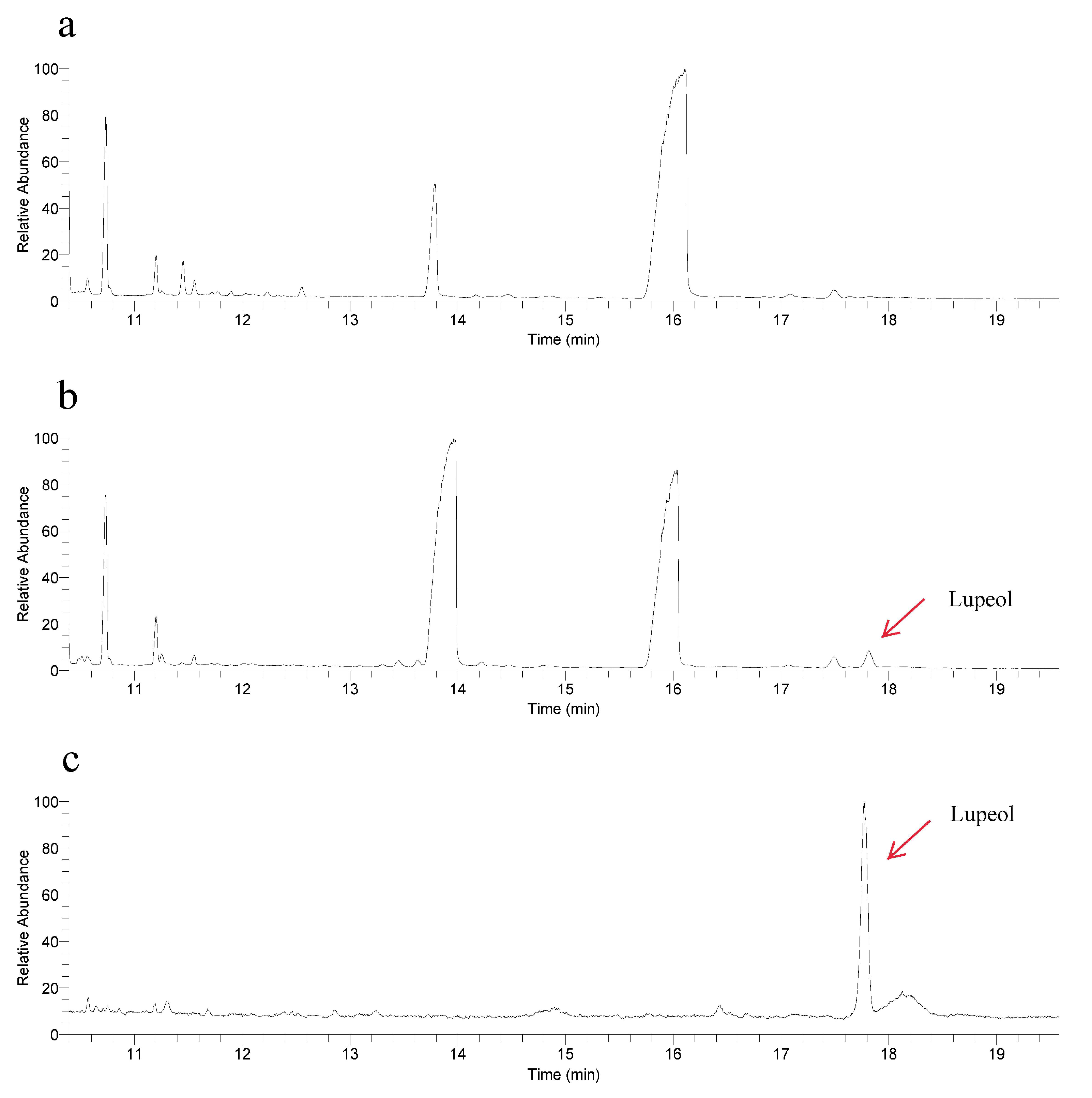

Figure 6.
The lateral branches (a), primary lateral branch (1), secondary lateral branch (2), tertiary lateral branch (3). Roots (b) and roots of tissue culture seedlings (c) (4). Petals at different development stages (d), S1: Large bud stage, S2: Reddish stage, S3: Flowering initiation stage, S4: Flower bud half-opening stage, S5: Initial opening stage, S6: Semi-opening stage, S7: Full opening stage.
Figure 6.
The lateral branches (a), primary lateral branch (1), secondary lateral branch (2), tertiary lateral branch (3). Roots (b) and roots of tissue culture seedlings (c) (4). Petals at different development stages (d), S1: Large bud stage, S2: Reddish stage, S3: Flowering initiation stage, S4: Flower bud half-opening stage, S5: Initial opening stage, S6: Semi-opening stage, S7: Full opening stage.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, J.; Wang, P.; Xu, M.; Chen, Y.; Feng, L. Systematic Identification and Analysis of OSC Gene Family of Rosa rugosa Thunb. Int. J. Mol. Sci. 2022, 23, 13884. https://doi.org/10.3390/ijms232213884
AMA Style
Wang J, Wang P, Xu M, Chen Y, Feng L. Systematic Identification and Analysis of OSC Gene Family of Rosa rugosa Thunb. International Journal of Molecular Sciences. 2022; 23(22):13884. https://doi.org/10.3390/ijms232213884
Chicago/Turabian StyleWang, Jianwen, Pengqing Wang, Mengmeng Xu, Yudie Chen, and Liguo Feng. 2022. "Systematic Identification and Analysis of OSC Gene Family of Rosa rugosa Thunb" International Journal of Molecular Sciences 23, no. 22: 13884. https://doi.org/10.3390/ijms232213884
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.