
Cardiovascular Characteristics of Zucker Fatty Diabetes Mellitus Rats, an Animal Model for Obesity and Type 2 Diabetes


Abstract
:1. Introduction
2. Results
2.1. The Metabolic Characteristics of ZFDM Rats
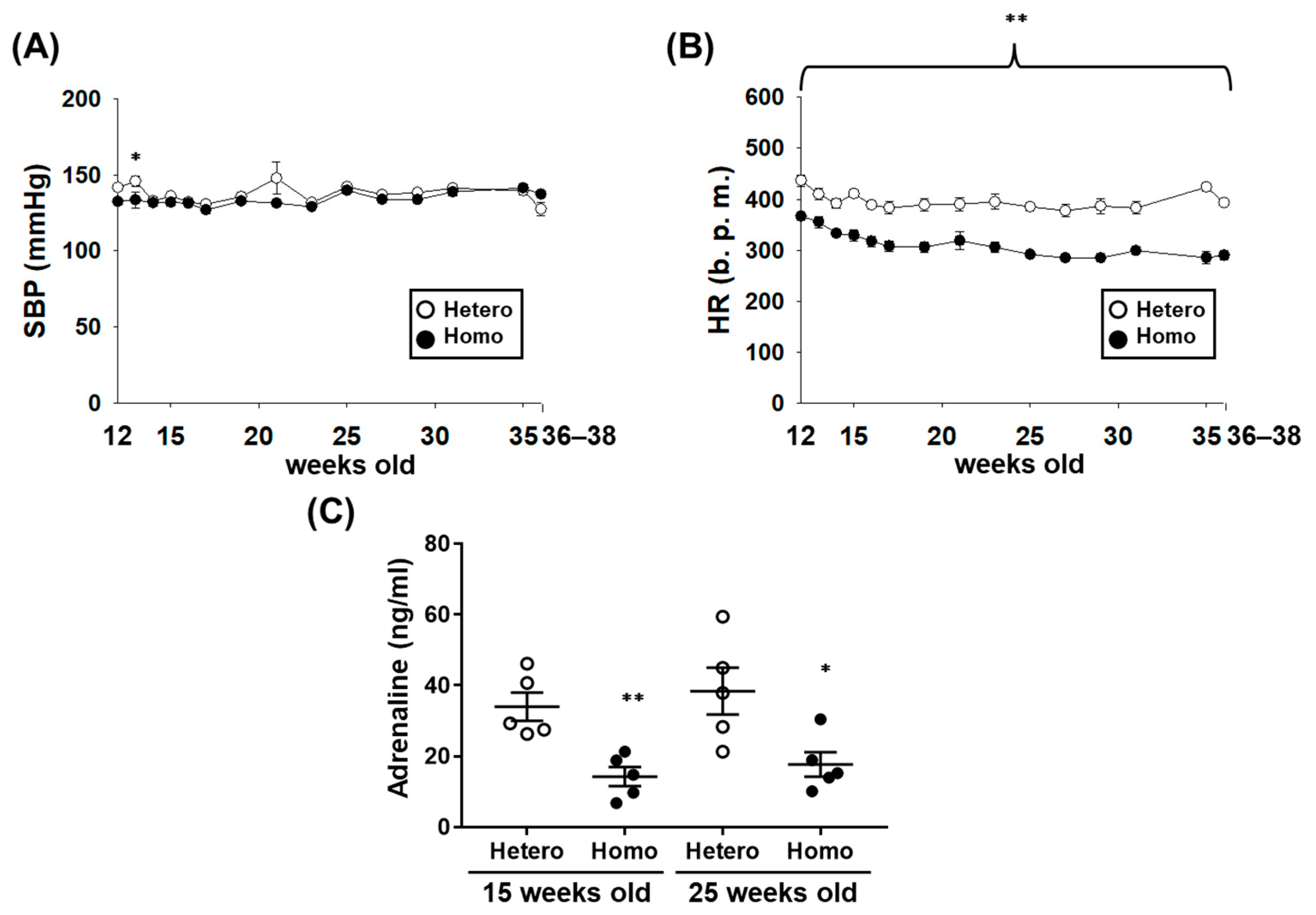
2.2. The Cardiovascular Dynamics of ZFDM Rats
2.3. Histological Analysis of Ventricles and Mesenteric Arteries from ZFDM Rats
2.4. Histology and mRNA Expression in Adipocytes from ZFDM Rats
2.5. Changes in Weight and Biochemical Characteristics of Kidney from ZFDM Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Calculation of BMI
4.3. Blood Glucose Measurement
4.4. Plasma Insulin, Triglyceride, Total Cholesterol, BUN, and Creatinine Measurements
4.5. Measurement of SBP and HR
4.6. Plasma Adrenaline Measurement
4.7. Histological Analysis
4.8. RT-qPCR Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Obesity. Available online: https://www.who.int/health-topics/obesity#tab=tab_1 (accessed on 28 February 2022).
- Dai, H.; Alsalhe, T.A.; Chalghaf, N.; Riccò, M.; Bragazzi, N.L.; Wu, J. The Global Burden of Disease Attributable to High Body Mass Index in 195 Countries and Territories, 1990–2017: An Analysis of the Global Burden of Disease Study. PLoS Med. 2020, 17, e1003198. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global Prevalence of Diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Reinehr, T. Type 2 diabetes mellitus in children and adolescents. World J. Diabetes 2013, 4, 270–281. [Google Scholar] [CrossRef]
- Reddy, M.A.; Park, J.T.; Natarajan, R. Epigenetic Modifications in the Pathogenesis of Diabetic Nephropathy. Semin. Nephrol. 2013, 33, 341–353. [Google Scholar] [CrossRef] [Green Version]
- de Boer, I.H.; Rue, T.C.; Hall, Y.N.; Heagerty, P.J.; Weiss, N.S.; Himmelfarb, J. Temporal Trends in the Prevalence of Diabetic Kidney Disease in the United States. JAMA 2011, 305, 2532–2539. [Google Scholar] [CrossRef]
- Viigimaa, M.; Sachinidis, A.; Toumpourleka, M.; Koutsampasopoulos, K.; Alliksoo, S.; Tiinatma, T. Macrovascular Complications of Type 2 Diabetes Mellitus. Curr. Vasc. Pharmacol. 2020, 18, 110–116. [Google Scholar] [CrossRef]
- Peterson, R.G.; Shaw, W.N.; Neel, M.A.; Little, L.A.; Eichberg, J. Zucker Diabetic Fatty Rat as a Model for Non-insulin-dependent Diabetes Mellitus. ILAR J. 1990, 32, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Kawano, K.; Hirashima, T.; Mori, S.; Saitoh, Y.; Kurosumi, M.; Natori, T. Spontaneous Long-Term Hyperglycemic Rat with Diabetic Complications. Diabetes 1992, 41, 7. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Shino, A.; Matsuo, T.; Iwatsuka, H.; Suzuoki, Z. A New Genetically Obese-Hyperglycemic Rat (Wistar Fatty). Diabetes 1981, 30, 6. [Google Scholar] [CrossRef] [PubMed]
- Hummel, K.P.; Dickie, M.M.; Coleman, D.L. Diabetes, a New Mutation in the Mouse. Science 1966, 153, 1127–1128. [Google Scholar] [CrossRef] [PubMed]
- Saito, F.; Kawaguchi, M.; Izumida, J.; Asakura, T.; Maehara, K.; Maruyama, Y. Alteration in haemodynamics and pathological changes in the cardiovascular system during the development of Type 2 diabetes mellitus in OLETF rats. Diabetologia 2003, 46, 1161–1169. [Google Scholar] [CrossRef]
- Marsh, S.A.; Powell, P.C.; Agarwal, A.; Dell’Italia, L.J.; Chatham, J.C. Cardiovascular dysfunction in Zucker obese and Zucker diabetic fatty rats: Role of hydronephrosis. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H292–H298. [Google Scholar] [CrossRef]
- Yokoi, N.; Hoshino, M.; Hidaka, S.; Yoshida, E.; Beppu, M.; Hoshikawa, R.; Sudo, K.; Kawada, A.; Takagi, S.; Seino, S. A Novel Rat Model of Type 2 Diabetes: The Zucker Fatty Diabetes Mellitus ZFDM Rat. J. Diabetes Res. 2013, 2013, 103731. [Google Scholar] [CrossRef]
- Gheni, G.; Yokoi, N.; Beppu, M.; Yamaguchi, T.; Hidaka, S.; Kawabata, A.; Hoshino, Y.; Hoshino, M.; Seino, S. Characterization of the Prediabetic State in a Novel Rat Model of Type 2 Diabetes, the ZFDM Rat. J. Diabetes Res. 2015, 2015, 261418. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, H.; Jacobs, D., Jr. Commentary: Origins and evolution of body mass index (BMI): Continuing saga. Int. J. Epidemiol. 2014, 43, 5. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, S.E.; Gee, L.L.; Wachtel, M.S.; Frezza, E.E. Adipose Tissue: The New Endocrine Organ? A Review Article. Dig. Dis. Sci. 2009, 54, 1847–1856. [Google Scholar] [CrossRef]
- Bełtowski, J. Adiponectin and resistin-new hormones of white adipose tissue. Med. Sci. Monit. 2003, 9, RA55–RA61. [Google Scholar]
- Chabowska-Kita, A.; Kozak, L.P. The critical period for brown adipocyte development: Genetic and environmental influences. Obesity 2016, 24, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Pagani, M.; Malliani, A. Interpreting oscillations of muscle sympathetic nerve activity and heart rate variability. J. Hypertens. 2000, 18, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Aizawa-Abe, M.; Ogawa, Y.; Masuzaki, H.; Ebihara, K.; Satoh, N.; Iwai, H.; Matsuoka, N.; Hayashi, T.; Hosoda, K.; Inoue, G.; et al. Pathophysiological role of leptin in obesity-related hypertension. J. Clin. Investig. 2000, 105, 1243–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmouni, K.; Morgan, D.A. Hypothalamic arcuate nucleus mediates the sympathetic and arterial pressure responses to leptin. Hypertension 2007, 49, 647–652. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, T.; Murakami, T.; Iida, M.; Kuwajima, M.; Shima, K. Leptin Receptor of Zucker Fatty Rat Performs Reduced Signal Transduction. Diabetes 1997, 46, 1077–1080. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, R.H.; Abel, B.D. Basic Mechanisms of Diabetic Heart Disease. Circ. Res. 2020, 126, 1501–1525. [Google Scholar] [CrossRef]
- Fredersdorf, S.; Thumann, C.; Ulucan, C.; Griese, D.P.; Luchner, A.; Riegger, G.A.; Kromer, E.P.; Weil, J. Myocardial hypertrophy and enhanced left ventricular contractility in Zucker diabetic fatty rats. Cardiovasc. Pathol. 2004, 13, 9–11. [Google Scholar]
- Schäfer, S.; Huber, J.; Wihler, C.; Rütten, H.; Busch, A.E.; Linz, W. Impaired left ventricular relaxation in type 2 diabetic rats is related to myocardial accumulation of N(epsilon)-(carboxymethyl) lysine. Eur. J. Heart Fail. 2006, 8, 2–6. [Google Scholar] [CrossRef]
- Makino, N.; Maeda, T.; Oyama, J.; Higuchi, Y.; Mimori, K. Improving insulin sensitivity via activation of PPAR-γ increases telomerase activity in the heart of OLETF rats. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H2188–H2195. [Google Scholar] [CrossRef]
- Howarth, F.C.; Jacobson, M.; Shafiullah, M.; Adeghate, E. Long-term effects of type 2 diabetes mellitus on heart rhythm in the Goto-Kakizaki rat. Exp. Physiol. 2008, 93, 362–369. [Google Scholar] [CrossRef]
- Chandler, M.P.; Morgan, E.E.; McElfresh, T.A.; Kung, T.A.; Rennison, J.H.; Hoit, B.D.; Young, M.E. Heart failure progression is accelerated following myocardial infarction in type 2 diabetic rats. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1609–H1616. [Google Scholar] [CrossRef] [PubMed]
- Hepler, C.; Gupta, R.K. The Expanding Problem of Adipose Depot Remodeling and Postnatal Adipocyte Progenitor Recruitment. Mol. Cell Endocrinol. 2017, 445, 95–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, J.; Han, P.W. Cellularity of rat adipose tissue: Effects of growth, starvation, and obesity. J. Lipid Res. 1969, 10, 77–82. [Google Scholar] [CrossRef]
- Kotzbeck, P.; Giordano, A.; Mondini, E.; Murano, I.; Severi, I.; Venema, W.; Cecchini, M.P.; Kershaw, E.E.; Barbatelli, G.; Haemmerle, G.; et al. Brown adipose tissue whitening leads to brown adipocyte death and adipose tissue inflammation. J. Lipid Res. 2018, 59, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Bost, F.; Kaminski, L. The metabolic modulator PGC-1α in cancer. Am. J. Cancer Res. 2019, 9, 198–211. [Google Scholar]
- Fenzl, A.; Kiefer, F.W. Brown. adipose tissue and thermogenesis. Horm. Mol. Biol. Clin. Investig. 2014, 19, 25–37. [Google Scholar] [CrossRef]
- Chondronikola, M.; Volpi, E.; Børsheim, E.; Porter, C.; Annamalai, P.; Enerbäck, S.; Lidell, M.E.; Saraf, M.K.; Labbe, S.M.; Hurren, N.M.; et al. Brown Adipose Tissue Improves Whole-Body Glucose Homeostasis and Insulin Sensitivity in Humans. Diabetes 2014, 63, 4089–4099. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.C.; Brownlee, M.; Susztak, K.; Sharma, K.; Jandeleit-Dahm, K.A.M.; Zoungas, S.; Rossing, P.; Groop, P.H.; Cooper, M.E. Diabetic kidney disease. Nat. Rev. Dis. Primes 2015, 1, 15018. [Google Scholar] [CrossRef]
- Siwy, J.; Zoja, C.; Klein, J.; Benigni, A.; Mullen, W.; Mayer, B.; Mischak, H.; Jankowski, J.; Stevens, R.; Vlahou, A.; et al. Evaluation of the Zucker diabetic fatty (ZDF) rat as a model for human disease based on urinary peptidomic profiles. PLoS ONE 2012, 7, e51334. [Google Scholar] [CrossRef]
- Togashi, Y.; Miyamoto, Y. Urinary cystatin C as a biomarker for diabetic nephropathy and its immunohistochemical localization in kidney in Zucker diabetic fatty (ZDF) rats. Exp. Toxicol. Pathol. 2013, 65, 615–622. [Google Scholar] [CrossRef]
- Miyata, K.; Ohashi, N.; Suzaki, Y.; Katsurada, A.; Kobori, H. Sequential activation of the reactive oxygen species/angiotensinogen/renin-angiotensin system axis in renal injury of type 2 diabetic rats. Clin. Exp. Pharmacol. Physiol. 2008, 35, 922–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuzawa, Y.; Watanabe, Y.; Inaguma, D.; Hotta, N. Evaluation of glomerular lesion and abnormal urinary findings in OLETF rats resulting from a long-term diabetic state. J. Lab. Clin. Med. 1996, 128, 568–578. [Google Scholar] [CrossRef]
- Yabuki, A.; Tahara, T.; Taniguchi, K.; Matsumoto, M.; Suzuki, S. Neuronal nitric oxide synthase and cyclooxygenase-2 in diabetic nephropathy of type 2 diabetic OLETF rats. Exp. Anim. 2006, 55, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashima, S.; Inoue, K.; Matsumoto, M. Low creatinine levels in diabetes mellitus among older individuals: The Yuport Medical Checkup Center Study. Sci. Rep. 2021, 11, 15167. [Google Scholar] [CrossRef]
- Engelbregt, M.J.T.; van Weissenbruch, M.M.; Popp-Snijders, C.; Lips, P.; Delemarre-van de Waal, H.A. Body Mass Index, Body Composition, and Leptin at Onset of Puberty in Male and Female Rats after Intrauterine Growth Retardation and after Early Postnatal Food Restriction. Pediatr. Res. 2001, 50, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Sugisawa, R.; Hiramoto, E.; Matsuoka, S.; Iwai, S.; Takai, R.; Yamazaki, T.; Mori, N.; Okada, Y.; Takeda, N.; Yamamura, K. Impact of feline AIM on the susceptibility of cats to renal disease. Sci. Rep. 2016, 6, 35251. [Google Scholar] [CrossRef] [Green Version]
- Otani, K.; Yokoya, M.; Kodama, T.; Hori, K.; Matsumoto, K.; Okada, M.; Yamawaki, H. Plasma exosomes regulate systemic blood pressure in rats. Biochem. Biophys. Res. Commun. 2018, 503, 776–783. [Google Scholar] [CrossRef]
- Sugiyama, A.; Okada, M.; Yamawaki, H. Canstatin suppresses isoproterenol-induced cardiac hypertrophy through inhibition of calcineurin/nuclear factor of activated T-cells pathway in rats. Eur. J. Pharmacol. 2020, 871, 172849. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hetero (n = 8) | Homo (n = 9) | |
|---|---|---|
| BW (g) | 450.2 ± 8.8 | 389.6 ± 14.7 ** |
| Body length (cm) | 23.1 ± 0.1 | 20.3 ± 0.2 ** |
| BMI (g/cm2) | 0.85 ± 0.01 | 0.94 ± 0.03 ** |
| Blood glucose level (mg/dL) | 162.5 ± 8.2 | 504.3 ± 20.0 ** |
| Triglyceride (mg/dL) | 56.5 ± 8.8 | 681.2 ± 85.2 ** |
| Total cholesterol (mg/dL) | 90.9 ± 4.3 | 271.8 ± 18.7 ** |
| Heart/BW (mg/g) | 3.2 ± 0.1 | 3.5 ± 0.1 * |
| SBP (mmHg) | 130.5 ± 1.9 | 127.4 ± 4.4 |
| HR (b. p. m.) | 393.8 ± 8.6 | 282.9 ± 11.2 ** |
| Perirenal adipocytes/BW (mg/g) | 13.8 ± 1.0 | 48.5 ± 3.5 ** |
| Interscapular adipocytes/BW (mg/g) | 1.5 ± 0.1 | 2.1 ± 0.3 * |
| Kidney/BW (mg/g) | 5.8 ± 0.2 | 8.6 ± 0.2 ** |
| BUN (mg/dL) | 24.1 ± 0.7 | 27.4 ± 0.8 * |
| Creatinine (mg/dL) | 0.65 ± 0.03 | 0.33 ± 0.01 ** |
| Products | Primer Sequences | Accession Number | |
|---|---|---|---|
| MCP-1 | Reverse | 5′-CCAATGAGTCGGCTGGAGAACT-3′ | NM_031530.1 |
| Forward | 5′-AGTGCTTGAGGTGGTTGTGGAA-3′ | ||
| UCP-1 | Reverse | 5′-GCCTCTACGATACGGTCCAA-3′ | NM_012682.2 |
| Forward | 5′-CTGACCTTCACCACCTCTGT-3′ | ||
| PGC-1α | Reverse | 5′-ACCCACAGGATCAGAACAAACC-3′ | NM_031347.1 |
| Forward | 5′-GACAAATGCTCTTTGCTTTATTGC-3′ | ||
| Adiponectin | Reverse | 5′-GAAGGGAGACGCAGGTGTTC-3′ | NM_144744.3 |
| Forward | 5′-GGGAACATTGGGGACAGTGA-3′ | ||
| GAPDH | Reverse | 5′-GAAGACGCCAGTAGACTCCA-3′ | NM_017008.4 |
| Forward | 5′-GAGAATGGGAAGCTGGTCAT-3′ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otani, K.; Funada, H.; Teranishi, R.; Okada, M.; Yamawaki, H. Cardiovascular Characteristics of Zucker Fatty Diabetes Mellitus Rats, an Animal Model for Obesity and Type 2 Diabetes. Int. J. Mol. Sci. 2022, 23, 4228. https://doi.org/10.3390/ijms23084228
Otani K, Funada H, Teranishi R, Okada M, Yamawaki H. Cardiovascular Characteristics of Zucker Fatty Diabetes Mellitus Rats, an Animal Model for Obesity and Type 2 Diabetes. International Journal of Molecular Sciences. 2022; 23(8):4228. https://doi.org/10.3390/ijms23084228
Chicago/Turabian StyleOtani, Kosuke, Hiroshi Funada, Risa Teranishi, Muneyoshi Okada, and Hideyuki Yamawaki. 2022. "Cardiovascular Characteristics of Zucker Fatty Diabetes Mellitus Rats, an Animal Model for Obesity and Type 2 Diabetes" International Journal of Molecular Sciences 23, no. 8: 4228. https://doi.org/10.3390/ijms23084228