
SurA-like and Skp-like Proteins as Important Virulence Determinants of the Gram Negative Bacterial Pathogens

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Export of Virulence Factors
- Two-step—proteins first pass through the inner membrane (using the SEC or TAT system), traverse the periplasm, reach the outer membrane (OM), where they are either incorporated into the OM with the participation of the β-barrel assembly machinery (the BAM complex) or exported outside the cell by dedicated secretory systems (e.g., type 2, 5, 8 or 9 secretion systems). Both SEC and TAT substrates contain cleavable N-terminal signal sequences which are cleaved during or shortly after translocation. However, the TAT system translocates folded substrates, whereas the SEC substrates are translocated in unfolded state [18,19].
3. Two-Step Bacterial Secretion Systems
3.1. The Type Five Secretion System (T5SS)
3.2. The Type 2 Secretion System (T2SS)
3.3. The Type 8 Secretion System (T8SS)
3.4. The Type 9 Secretion System (T9SS)
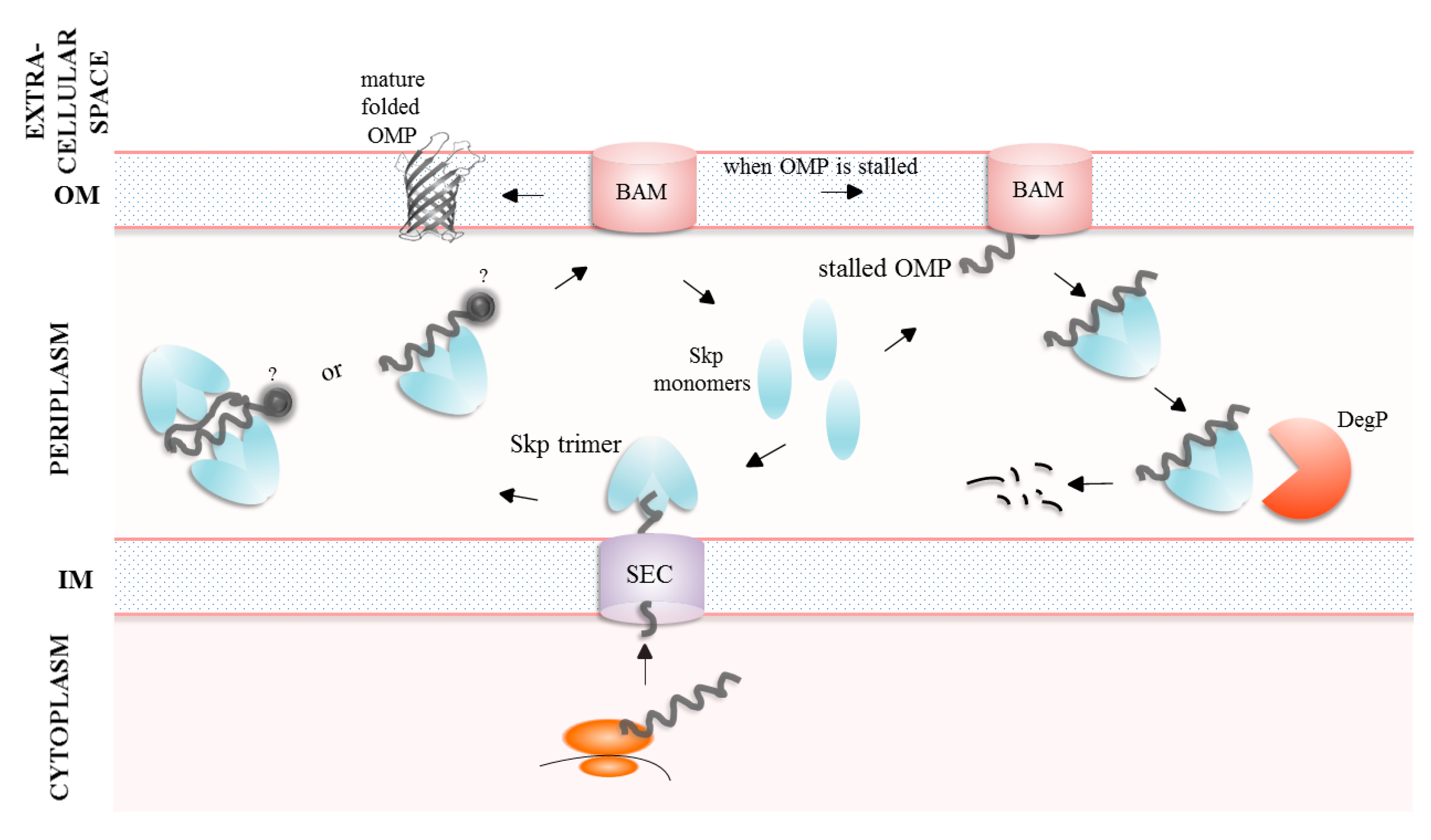
4. Importance of the Periplasmic Folding Factors for the Efficient Transport and Secretion of Proteins
4.1. The SurA Protein
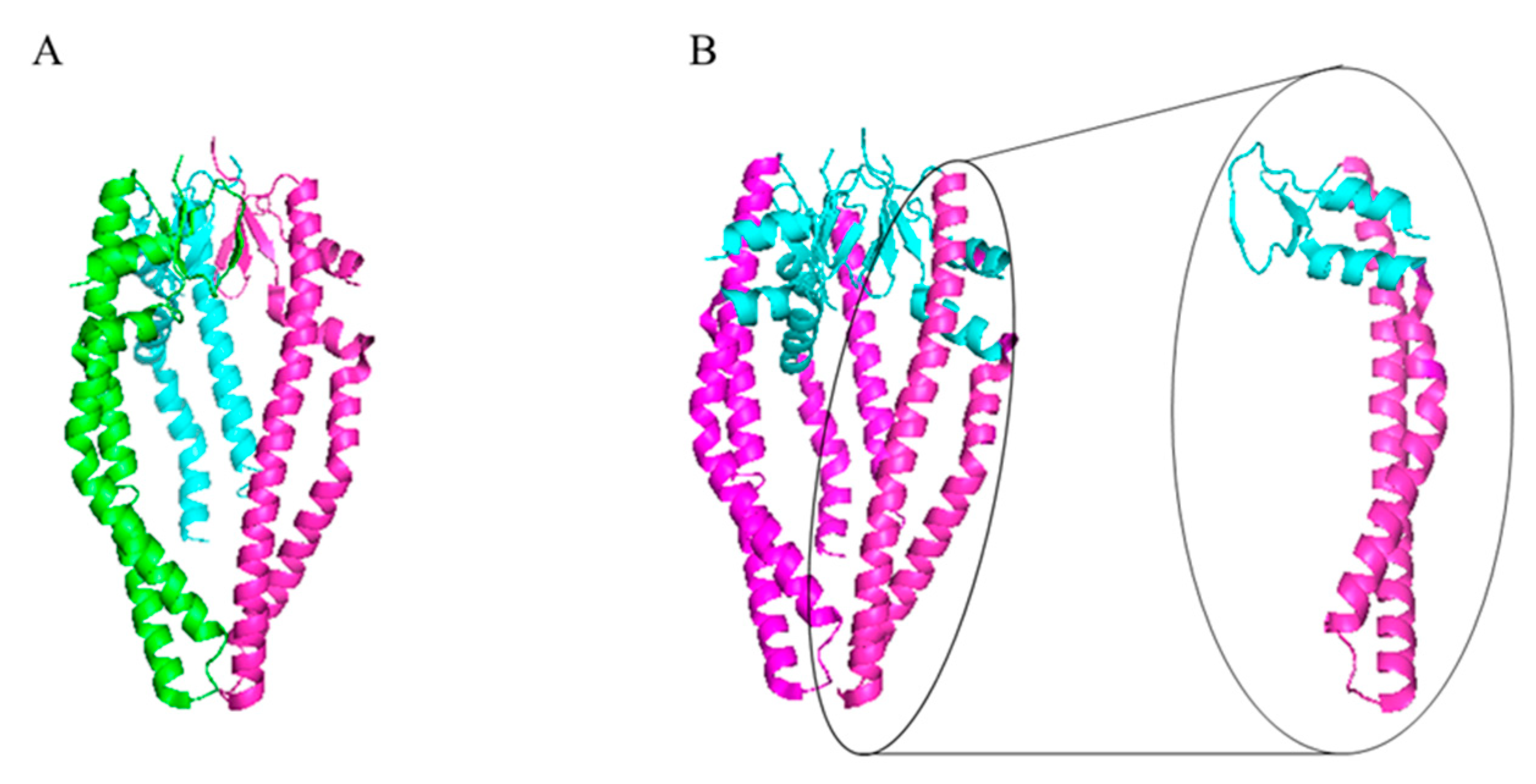
4.2. The Skp Protein
5. Virulence-Related Processes Dependent on the SurA and/or Skp Functions
5.1. Adhession and Dissemination within the Host
5.2. Antibiotic Resistance
5.3. Evasion of the Host Immune Systems
5.4. Acquisition of Nutrients and Metal Ions
5.5. Secretion of Toxins
5.6. Biofilm Formation
6. SurA-like Proteins as the Extracellular Virulence Factors
7. SurA and Skp-like Proteins as Therapeutic Targets
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johnson, D.I. Bacterial virulence factors. In Bacterial Pathogens and Their Virulence Factors; Johnson, D.I., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–38. ISBN 978-3-319-67651-7. [Google Scholar]
- Josenhans, C.; Suerbaum, S. The role of motility as a virulence factor in bacteria. Int. J. Med. Microbiol. 2002, 291, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Klemm, P.; Schembri, M.A. Bacterial adhesins: Function and structure. Int. J. Med. Microbiol. 2000, 290, 27–35. [Google Scholar] [CrossRef]
- Pugazhendhi, A.S.; Wei, F.; Hughes, M.; Coathup, M. Bacterial adhesion, virulence, and biofilm formation. In Musculoskeletal Infection; Coathup, M., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 19–64. ISBN 978-3-030-83251-3. [Google Scholar]
- Zheng, M.; Sun, S.; Zhou, J.; Liu, M. Virulence factors impair epithelial junctions during bacterial infection. J. Clin. Lab. Anal. 2021, 35, e23627. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.B.; McFadden, G. Anti-Immunology: Evasion of the host immune system by bacterial and viral pathogens. Cell 2006, 124, 767–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.W.; Schurr, M.J.; LeBlanc, C.L.; Ramamurthy, R.; Buchanan, K.L.; Nickerson, C.A. Mechanisms of bacterial pathogenicity. Postgrad. Med. J. 2002, 78, 216–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, N.; Kahne, D.; Silhavy, T.J. Advances in understanding bacterial outer-membrane biogenesis. Nat. Rev. Microbiol. 2006, 4, 57–66. [Google Scholar] [CrossRef]
- Depluverez, S.; Devos, S.; Devreese, B. The role of bacterial secretion systems in the virulence of gram-negative airway pathogens associated with cystic fibrosis. Front. Microbiol. 2016, 7, 1336. [Google Scholar] [CrossRef] [Green Version]
- Missiakas, D.; Raina, S. Protein folding in the bacterial periplasm. J. Bacteriol. 1997, 179, 2465–2471. [Google Scholar] [CrossRef] [Green Version]
- Merdanovic, M.; Clausen, T.; Kaiser, M.; Huber, R.; Ehrmann, M. Protein quality control in the bacterial periplasm. Annu. Rev. Microbiol. 2011, 65, 149–168. [Google Scholar] [CrossRef]
- Miot, M.; Betton, J.-M. Protein quality control in the bacterial periplasm. Microb. Cell Fact. 2004, 3, 4. [Google Scholar] [CrossRef]
- Fang, F.C.; Frawley, E.R.; Tapscott, T.; Vázquez-Torres, A. Bacterial stress responses during host infection. Cell Host Microbe 2016, 20, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandeya, A.; Ojo, I.; Alegun, O.; Wei, Y. Periplasmic targets for the development of effective antimicrobials against gram-negative bacteria. ACS Infect. Dis. 2020, 6, 2337–2354. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.; Allan, E.; Coates, A.R.M. Stress Wars: The direct role of host and bacterial molecular chaperones in bacterial infection. Infect. Immun. 2006, 74, 3693–3706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koster, M.; Bitter, W.; Tommassen, J. Protein secretion mechanisms in gram-negative bacteria. Int. J. Med. Microbiol. 2000, 290, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.R.D.; Felisberto-Rodrigues, C.; Meir, A.; Prevost, M.S.; Redzej, A.; Trokter, M.; Waksman, G. Secretion systems in gram-negative bacteria: Structural and mechanistic insights. Nat. Rev. Microbiol. 2015, 13, 343–359. [Google Scholar] [CrossRef]
- Robinson, C.; Bolhuis, A. Tat-dependent protein targeting in prokaryotes and chloroplasts. Biochim. Biophys. Acta BBA Mol. Cell Res. 2004, 1694, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Natale, P.; Brüser, T.; Driessen, A.J.M. SEC- and TAT-mediated protein secretion across the bacterial cytoplasmic membrane—Distinct translocases and mechanisms. Biochim. Biophys. Acta BBA Biomembr. 2008, 1778, 1735–1756. [Google Scholar] [CrossRef] [Green Version]
- Stathopoulos, C.; Hendrixson, D.R.; Thanassi, D.G.; Hultgren, S.J.; Geme, J.W.S., III; Curtiss, R., III. Secretion of virulence determinants by the general secretory pathway in gram-negative pathogens: An evolving story. Microbes Infect. 2000, 2, 1061–1072. [Google Scholar] [CrossRef]
- Wu, H.-J.; Wang, A.H.-J.; Jennings, M.P. Discovery of virulence factors of pathogenic bacteria. Curr. Opin. Chem. Biol. 2008, 12, 93–101. [Google Scholar] [CrossRef]
- Mogensen, J.E.; Otzen, D.E. Interactions between folding factors and bacterial outer membrane proteins. Mol. Microbiol. 2005, 57, 326–346. [Google Scholar] [CrossRef]
- Facey, S.J.; Kuhn, A. Biogenesis of bacterial inner-membrane proteins. Cell. Mol. Life Sci. 2010, 67, 2343–2362. [Google Scholar] [CrossRef] [PubMed]
- Dalbey, R.E.; Kuhn, A. Protein traffic in gram-negative bacteria—How exported and secreted proteins find their way. FEMS Microbiol. Rev. 2012, 36, 1023–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koebnik, R.; Locher, K.P.; van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Fairman, J.W.; Noinaj, N.; Buchanan, S.K. The structural biology of β-barrel membrane proteins: A summary of recent reports. Curr. Opin. Struct. Biol. 2011, 21, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dautin, N. Folding control in the path of type 5 secretion. Toxins 2021, 13, 341. [Google Scholar] [CrossRef]
- Burdette, L.A.; Leach, S.A.; Wong, H.T.; Tullman-Ercek, D. Developing gram-negative bacteria for the secretion of heterologous proteins. Microb. Cell Fact. 2018, 17, 196. [Google Scholar] [CrossRef] [Green Version]
- Heinz, E.; Selkrig, J.; Belousoff, M.J.; Lithgow, T. Evolution of the translocation and assembly module (TAM). Genome Biol. Evol. 2015, 7, 1628–1643. [Google Scholar] [CrossRef] [Green Version]
- Meuskens, I.; Saragliadis, A.; Leo, J.C.; Linke, D. Type V secretion systems: An overview of passenger domain functions. Front. Microbiol. 2019, 10, 1163. [Google Scholar] [CrossRef] [Green Version]
- Coppens, F.; Castaldo, G.; Debraekeleer, A.; Subedi, S.; Moonens, K.; Lo, A.; Remaut, H. Hop-family Helicobacter outer membrane adhesins form a novel class of type 5-like secretion proteins with an interrupted β-barrel domain. Mol. Microbiol. 2018, 110, 33–46. [Google Scholar] [CrossRef]
- Green, E.R.; Mecsas, J. Bacterial secretion systems: An overview. Microbiol. Spectr. 2016, 4, 4.1.13. [Google Scholar] [CrossRef]
- Pohlner, J.; Halter, R.; Beyreuther, K.; Meyer, T.F. Gene structure and extracellular secretion of Neisseria gonorrhoeae IgA protease. Nature 1987, 325, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Brandon, L.D.; Goehring, N.; Janakiraman, A.; Yan, A.W.; Wu, T.; Beckwith, J.; Goldberg, M.B. IcsA, a polarly localized autotransporter with an atypical signal peptide, uses the Sec apparatus for secretion, although the Sec apparatus is circumferentially distributed. Mol. Microbiol. 2003, 50, 45–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cover, T.L.; Blanke, S.R. Helicobacter pylori VacA, a paradigm for toxin multifunctionality. Nat. Rev. Microbiol. 2005, 3, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Sandkvist, M. Biology of type II secretion. Mol. Microbiol. 2001, 40, 271–283. [Google Scholar] [CrossRef]
- Nivaskumar, M.; Francetic, O. Type II secretion system: A magic beanstalk or a protein escalator. Biochim. Biophys. Acta 2014, 1843, 1568–1577. [Google Scholar] [CrossRef] [Green Version]
- Naskar, S.; Hohl, M.; Tassinari, M.; Low, H.H. The structure and mechanism of the bacterial type II secretion system. Mol. Microbiol. 2021, 115, 412–424. [Google Scholar] [CrossRef]
- Korotkov, K.V.; Sandkvist, M.; Hol, W.G.J. The type II secretion system: Biogenesis, molecular architecture and mechanism. Nat. Rev. Microbiol. 2012, 10, 336–351. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.M.; Lory, S. A specific targeting domain in mature exotoxin a is required for its extracellular secretion from Pseudomonas aeruginosa. EMBO J. 1996, 15, 429–436. [Google Scholar] [CrossRef]
- Sauvonnet, N.; Pugsley, A.P. Identification of two regions of Klebsiella oxytoca pullulanase that together are capable of promoting beta-lactamase secretion by the general secretory pathway. Mol. Microbiol. 1996, 22, 1–7. [Google Scholar] [CrossRef]
- Francetić, O.; Pugsley, A.P. Towards the identification of type II secretion signals in a nonacylated variant of pullulanase from Klebsiella oxytoca. J. Bacteriol. 2005, 187, 7045–7055. [Google Scholar] [CrossRef]
- Bhoite, S.; van Gerven, N.; Chapman, M.R.; Remaut, H. Curli biogenesis: Bacterial amyloid assembly by the type VIII secretion pathway. EcoSal Plus 2019, 8, 163–171. [Google Scholar] [CrossRef]
- Lasica, A.M.; Ksiazek, M.; Madej, M.; Potempa, J. The type IX secretion system (T9SS): Highlights and recent insights into its structure and function. Front. Cell. Infect. Microbiol. 2017, 7, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorasia, D.G.; Veith, P.D.; Reynolds, E.C. The type IX secretion system: Advances in structure, function and organisation. Microorganisms 2020, 8, 1173. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, M.; Aduse-Opoku, J.; Paramonov, N.; Hashim, A.; Bostanci, N.; Fraser, O.P.; Tarelli, E.; Curtis, M.A. Identification of a second lipopolysaccharide in Porphyromonas gingivalis W50. J. Bacteriol. 2008, 190, 2920–2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffrin, B.; Machin, J.M.; Karamanos, T.K.; Zhuravleva, A.; Brockwell, D.J.; Radford, S.E.; Calabrese, A.N. Dynamic interplay between the periplasmic chaperone SurA and the BAM complex in outer membrane protein folding. Commun. Biol. 2022, 5, 560. [Google Scholar] [CrossRef] [PubMed]
- Behrens, S.; Maier, R.; de Cock, H.; Schmid, F.X.; Gross, C.A. The SurA periplasmic PPIase lacking its parvulin domains functions in vivo and has chaperone activity. EMBO J. 2001, 20, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Sklar, J.G.; Wu, T.; Kahne, D.; Silhavy, T.J. Defining the roles of the periplasmic chaperones SurA, Skp, and DegP in Escherichia coli. Genes Dev. 2007, 21, 2473–2484. [Google Scholar] [CrossRef] [Green Version]
- Lazar, S.W.; Almirón, M.; Tormo, A.; Kolter, R. Role of the Escherichia coli SurA protein in stationary-phase survival. J. Bacteriol. 1998, 180, 5704–5711. [Google Scholar] [CrossRef] [Green Version]
- Tormo, A.; Almirón, M.; Kolter, R. surA, an Escherichia coli gene essential for survival in stationary phase. J. Bacteriol. 1990, 172, 4339–4347. [Google Scholar] [CrossRef] [Green Version]
- Bury-Moné, S.; Nomane, Y.; Reymond, N.; Barbet, R.; Jacquet, E.; Imbeaud, S.; Jacq, A.; Bouloc, P. Global analysis of extracytoplasmic stress signaling in Escherichia coli. PLoS Genet. 2009, 5, e1000651. [Google Scholar] [CrossRef]
- Lazar, S.W.; Kolter, R. SurA assists the folding of Escherichia coli outer membrane proteins. J. Bacteriol. 1996, 178, 1770–1773. [Google Scholar] [CrossRef] [Green Version]
- Missiakas, D.; Betton, J.-M.; Raina, S. New components of protein folding in extracytoplasmic compartments of Escherichia coli SurA, FkpA and Skp/OmpH. Mol. Microbiol. 1996, 21, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Rouvière, P.E.; Gross, C.A. SurA, a periplasmic protein with peptidyl-prolyl isomerase activity, participates in the assembly of outer membrane porins. Genes Dev. 1996, 10, 3170–3182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahfeld, J.U.; Rücknagel, K.P.; Schelbert, B.; Ludwig, B.; Hacker, J.; Mann, K.; Fischer, G. Confirmation of the existence of a third family among peptidyl-prolyl cis/trans isomerases. Amino acid sequence and recombinant production of parvulin. FEBS Lett. 1994, 352, 180–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitto, E.; McKay, D.B. Crystallographic structure of SurA, a molecular chaperone that facilitates folding of outer membrane porins. Structure 2002, 10, 1489–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, A.N.; Schiffrin, B.; Watson, M.; Karamanos, T.K.; Walko, M.; Humes, J.R.; Horne, J.E.; White, P.; Wilson, A.J.; Kalli, A.C.; et al. Inter-domain dynamics in the chaperone SurA and multi-site binding to its outer membrane protein clients. Nat. Commun. 2020, 11, 2155. [Google Scholar] [CrossRef]
- Marx, D.C.; Leblanc, M.J.; Plummer, A.M.; Krueger, S.; Fleming, K.G. Domain interactions determine the conformational ensemble of the periplasmic chaperone SurA. Protein Sci. 2020, 29, 2043–2053. [Google Scholar] [CrossRef]
- Humes, J.R.; Schiffrin, B.; Calabrese, A.N.; Higgins, A.J.; Westhead, D.R.; Brockwell, D.J.; Radford, S.E. The role of SurA PPIase domains in preventing aggregation of the outer-membrane proteins TOmpA and OmpT. J. Mol. Biol. 2019, 431, 1267–1283. [Google Scholar] [CrossRef]
- Justice, S.S.; Hunstad, D.A.; Harper, J.R.; Duguay, A.R.; Pinkner, J.S.; Bann, J.; Frieden, C.; Silhavy, T.J.; Hultgren, S.J. Periplasmic peptidyl prolyl cis-trans isomerases are not essential for viability, but SurA is required for pilus biogenesis in Escherichia coli. J. Bacteriol. 2005, 187, 7680–7686. [Google Scholar] [CrossRef] [Green Version]
- Vertommen, D.; Ruiz, N.; Leverrier, P.; Silhavy, T.J.; Collet, J.-F. Characterization of the role of the Escherichia coli periplasmic chaperone SurA using differential proteomics. Proteomics 2009, 9, 2432–2443. [Google Scholar] [CrossRef]
- Wu, T.; McCandlish, A.C.; Gronenberg, L.S.; Chng, S.-S.; Silhavy, T.J.; Kahne, D. Identification of a protein complex that assembles lipopolysaccharide in the outer membrane of Escherichia coli. Proc. Natl. Acad. Sci. USA 2006, 103, 11754–11759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitto, E.; McKay, D.B. The periplasmic molecular chaperone protein SurA binds a peptide motif that is characteristic of integral outer membrane proteins. J. Biol. Chem. 2003, 278, 49316–49322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitto, E.; McKay, D.B. Binding of phage-display-selected peptides to the periplasmic chaperone protein SurA mimics binding of unfolded outer membrane proteins. FEBS Lett. 2004, 568, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennecke, G.; Nolte, J.; Volkmer-Engert, R.; Schneider-Mergener, J.; Behrens, S. The periplasmic chaperone SurA exploits two features characteristic of integral outer membrane proteins for selective substrate recognition. J. Biol. Chem. 2005, 280, 23540–23548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Wang, S.; Hu, Y.-X.; McKay, D.B. The periplasmic bacterial molecular chaperone SurA adapts its structure to bind peptides in different conformations to assert a sequence preference for aromatic residues. J. Mol. Biol. 2007, 373, 367–381. [Google Scholar] [CrossRef] [Green Version]
- Mas, G.; Thoma, J.; Hiller, S. The periplasmic chaperones Skp and SurA. In Bacterial Cell Walls and Membranes; Subcellular Biochemistry; Kuhn, A., Ed.; Springer International Publishing: Cham, Switzerland, 2019; Volume 92, pp. 169–186. ISBN 978-3-030-18767-5. [Google Scholar]
- Marx, D.C.; Plummer, A.M.; Faustino, A.M.; Devlin, T.; Roskopf, M.A.; Leblanc, M.J.; Lessen, H.J.; Amann, B.T.; Fleming, P.J.; Krueger, S.; et al. SurA is a cryptically grooved chaperone that expands unfolded outer membrane proteins. Proc. Natl. Acad. Sci. USA 2020, 117, 28026–28035. [Google Scholar] [CrossRef]
- Plummer, A.M.; Fleming, K.G. From chaperones to the membrane with a BAM! Trends Biochem. Sci. 2016, 41, 872–882. [Google Scholar] [CrossRef] [Green Version]
- Chamachi, N.; Hartmann, A.; Ma, M.Q.; Svirina, A.; Krainer, G.; Schlierf, M. Chaperones Skp and SurA dynamically expand unfolded OmpX and synergistically disassemble oligomeric aggregates. Proc. Natl. Acad. Sci. USA 2022, 119, e2118919119. [Google Scholar] [CrossRef]
- Li, G.; He, C.; Bu, P.; Bi, H.; Pan, S.; Sun, R.; Zhao, X.S. Single-molecule detection reveals different roles of Skp and SurA as chaperones. ACS Chem. Biol. 2018, 13, 1082–1089. [Google Scholar] [CrossRef]
- He, C.; Pan, S.; Li, G.; Zhao, X.S. Molecular mechanism of SurA’s chaperoning function to outer membrane proteins revealed by purification-after-crosslinking single-molecule FRET. Sci. China Chem. 2020, 63, 1142–1152. [Google Scholar] [CrossRef]
- Alcock, F.H.; Grossmann, J.G.; Gentle, I.E.; Likić, V.A.; Lithgow, T.; Tokatlidis, K. Conserved substrate binding by chaperones in the bacterial periplasm and the mitochondrial intermembrane space. Biochem. J. 2007, 409, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weirich, J.; Bräutigam, C.; Mühlenkamp, M.; Franz-Wachtel, M.; Macek, B.; Meuskens, I.; Skurnik, M.; Leskinen, K.; Bohn, E.; Autenrieth, I.; et al. Identifying components required for OMP biogenesis as novel targets for antiinfective drugs. Virulence 2017, 8, 1170–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southern, S.J.; Scott, A.E.; Jenner, D.C.; Ireland, P.M.; Norville, I.H.; Sarkar-Tyson, M. Survival protein A is essential for virulence in Yersinia pestis. Microb. Pathog. 2016, 92, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Obi, I.R.; Nordfelth, R.; Francis, M.S. Varying dependency of periplasmic peptidylprolyl cis–trans isomerases in promoting Yersinia pseudotuberculosis stress tolerance and pathogenicity. Biochem. J. 2011, 439, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, A.; Phansopa, C.; Suwannachart, C.; Craven, C.J.; Rafferty, J.B.; Kelly, D.J. The virulence factor PEB4 (Cj0596) and the periplasmic protein Cj1289 are two structurally related SurA-like chaperones in the human pathogen Campylobacter jejuni. J. Biol. Chem. 2011, 286, 21254–21265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathbun, K.M.; Thompson, S.A. Mutation of PEB4 alters the outer membrane protein profile of Campylobacter jejuni. FEMS Microbiol. Lett. 2009, 300, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Klein, K.; Sonnabend, M.S.; Frank, L.; Leibiger, K.; Franz-Wachtel, M.; Macek, B.; Trunk, T.; Leo, J.C.; Autenrieth, I.B.; Schütz, M.; et al. Deprivation of the periplasmic chaperone SurA reduces virulence and restores antibiotic susceptibility of multidrug-resistant Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Gallagher, L.A.; Thongdee, M.; Staudinger, B.J.; Lippman, S.; Singh, P.K.; Manoil, C. General and condition-specific essential functions of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 5189–5194. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, B.E.; Yang, R.; Clatworthy, A.E.; White, T.; Osmulski, S.J.; Li, L.; Penaranda, C.; Lander, E.S.; Shoresh, N.; Hung, D.T. Defining the core essential genome of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2019, 116, 10072–10080. [Google Scholar] [CrossRef] [Green Version]
- Karash, S.; Yahr, T.L. Genome-wide identification of Pseudomonas aeruginosa genes important for desiccation tolerance on inanimate surfaces. mSystems 2022, 7, e00114-22. [Google Scholar] [CrossRef]
- www.uniprot.org. Available online: https://www.uniprot.org/uniprotkb/Q7VU12/entry (accessed on 19 December 2022).
- Hodak, H.; Wohlkönig, A.; Smet-Nocca, C.; Drobecq, H.; Wieruszeski, J.-M.; Sénéchal, M.; Landrieu, I.; Locht, C.; Jamin, M.; Jacob-Dubuisson, F. The peptidyl-prolyl isomerase and chaperone Par27 of Bordetella pertussis as the prototype for a new group of parvulins. J. Mol. Biol. 2008, 376, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Volokhina, E.B.; Grijpstra, J.; Stork, M.; Schilders, I.; Tommassen, J.; Bos, M.P. Role of the periplasmic chaperones Skp, SurA, and DegQ in outer membrane protein biogenesis in Neisseria meningitidis. J. Bacteriol. 2011, 193, 1612–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, T.A.; Sousa, M.C. Crystal structure of Skp, a prefoldin-like chaperone that protects soluble and membrane proteins from aggregation. Mol. Cell 2004, 15, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Korndörfer, I.P.; Dommel, M.K.; Skerra, A. Structure of the periplasmic chaperone Skp suggests functional similarity with cytosolic chaperones despite differing architecture. Nat. Struct. Mol. Biol. 2004, 11, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Combs, A.N.; Silhavy, T.J. The sacrificial adaptor protein Skp functions to remove stalled substrates from the β-barrel assembly machine. Proc. Natl. Acad. Sci. USA 2022, 119, e2114997119. [Google Scholar] [CrossRef]
- Mas, G.; Burmann, B.M.; Sharpe, T.; Claudi, B.; Bumann, D.; Hiller, S. Regulation of chaperone function by coupled folding and oligomerization. Sci. Adv. 2020, 6, eabc5822. [Google Scholar] [CrossRef]
- Walton, T.A.; Sandoval, C.M.; Fowler, C.A.; Pardi, A.; Sousa, M.C. The cavity-chaperone Skp protects its substrate from aggregation but allows independent folding of substrate domains. Proc. Natl. Acad. Sci. USA 2009, 106, 1772–1777. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, B.; Calabrese, A.N.; Devine, P.W.A.; Harris, S.A.; Ashcroft, A.E.; Brockwell, D.J.; Radford, S.E. Skp is a multivalent chaperone of outer-membrane proteins. Nat. Struct. Mol. Biol. 2016, 23, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Jarchow, S.; Lück, C.; Görg, A.; Skerra, A. Identification of potential substrate proteins for the periplasmic Escherichia coli chaperone Skp. Proteomics 2008, 8, 4987–4994. [Google Scholar] [CrossRef]
- Hirakawa, H.; Suzue, K.; Takita, A.; Kamitani, W.; Tomita, H. Roles of OmpX, an outer membrane protein, on virulence and flagellar expression in uropathogenic Escherichia coli. Infect. Immun. 2021, 89, e00721-20. [Google Scholar] [CrossRef]
- Papadopoulos, A.; Busch, M.; Reiners, J.; Hachani, E.; Baeumers, M.; Berger, J.; Schmitt, L.; Jaeger, K.-E.; Kovacic, F.; Smits, S.H.J.; et al. The periplasmic chaperone Skp prevents misfolding of the secretory lipase A from Pseudomonas aeruginosa. Front. Mol. Biosci. 2022, 9, 1026724. [Google Scholar] [CrossRef] [PubMed]
- Behrens-Kneip, S. The role of SurA factor in outer membrane protein transport and virulence. Int. J. Med. Microbiol. 2010, 300, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, M.A.; Lopez-Boado, Y.S.; Wilson, C.L.; Roth, R.; Parks, W.C.; Heuser, J.; Hultgren, S.J. Induction and evasion of host defenses by type 1-piliated uropathogenic Escherichia coli. Science 1998, 282, 1494–1497. [Google Scholar] [CrossRef] [PubMed]
- Justice, S.S.; Lauer, S.R.; Hultgren, S.J.; Hunstad, D.A. Maturation of intracellular Escherichia coli communities requires SurA. Infect. Immun. 2006, 74, 4793–4800. [Google Scholar] [CrossRef] [Green Version]
- Watts, K.M.; Hunstad, D.A. Components of SurA required for outer membrane biogenesis in uropathogenic Escherichia coli. PLoS ONE 2008, 3, e3359. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, M.B.; Theriot, J.A.; Sansonetti, P.J. Regulation of surface presentation of IcsA, a Shigella protein essential to intracellular movement and spread, is growth phase dependent. Infect. Immun. 1994, 62, 5664–5668. [Google Scholar] [CrossRef] [Green Version]
- Wagner, J.K.; Heindl, J.E.; Gray, A.N.; Jain, S.; Goldberg, M.B. Contribution of the periplasmic chaperone Skp to efficient presentation of the autotransporter IcsA on the surface of Shigella flexneri. J. Bacteriol. 2009, 191, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Roggenkamp, A.; Neuberger, H.R.; Flügel, A.; Schmoll, T.; Heesemann, J. Substitution of two histidine residues in YadA protein of Yersinia enterocolitica abrogates collagen binding, cell adherence and mouse virulence. Mol. Microbiol. 1995, 16, 1207–1219. [Google Scholar] [CrossRef]
- Isberg, R.R.; Leong, J.M. Multiple β1 chain integrins are receptors for invasin, a protein that promotes bacterial penetration into mammalian cells. Cell 1990, 60, 861–871. [Google Scholar] [CrossRef]
- Asakura, H.; Yamasaki, M.; Yamamoto, S.; Igimi, S. Deletion of peb4 gene impairs cell adhesion and biofilm formation in Campylobacter jejuni. FEMS Microbiol. Lett. 2007, 275, 278–285. [Google Scholar] [CrossRef]
- Rathbun, K.M.; Hall, J.E.; Thompson, S.A. Cj0596 is a periplasmic peptidyl prolyl cis-trans isomerase involved in Campylobacter jejuni motility, invasion, and colonization. BMC Microbiol. 2009, 9, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, C.L.; Shah, D.H.; Dhillon, A.S.; Call, D.R.; Ahn, S.; Haldorson, G.J.; Davitt, C.; Konkel, M.E.Y. Campylobacter jejuni invade chicken LMH cells inefficiently and stimulate differential expression of the chicken CXCLi1 and CXCLi2 cytokines. Microbiology 2008, 154, 3835–3847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular bacterial biofilm-like pods in urinary tract infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justice, S.S.; Hung, C.; Theriot, J.A.; Fletcher, D.A.; Anderson, G.G.; Footer, M.J.; Hultgren, S.J. Differentiation and developmental pathways of uropathogenic Escherichia coli in urinary tract pathogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 1333–1338. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, T.F.; Watts, K.M.; Hunstad, D.A. OmpA of uropathogenic Escherichia coli promotes postinvasion pathogenesis of cystitis. Infect. Immun. 2009, 77, 5245–5251. [Google Scholar] [CrossRef] [Green Version]
- Hunstad, D.A.; Justice, S.S.; Hung, C.S.; Lauer, S.R.; Hultgren, S.J. Suppression of bladder epithelial cytokine responses by uropathogenic Escherichia coli. Infect. Immun. 2005, 73, 3999–4006. [Google Scholar] [CrossRef] [Green Version]
- Hilbert, D.W.; Pascal, K.E.; Libby, E.K.; Mordechai, E.; Adelson, M.E.; Trama, J.P. Uropathogenic Escherichia coli dominantly suppress the innate immune response of bladder epithelial cells by a lipopolysaccharide- and Toll-like receptor 4-independent pathway. Microbes Infect. 2008, 10, 114–121. [Google Scholar] [CrossRef]
- Lau, M.E.; Loughman, J.A.; Hunstad, D.A. YbcL of uropathogenic Escherichia coli suppresses transepithelial neutrophil migration. Infect. Immun. 2012, 80, 4123–4132. [Google Scholar] [CrossRef] [Green Version]
- Dunkelberger, J.R.; Song, W.-C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [Green Version]
- Biedzka-Sarek, M.; Venho, R.; Skurnik, M. Role of YadA, ail, and lipopolysaccharide in serum resistance of Yersinia enterocolitica serotype O:3. Infect. Immun. 2005, 73, 2232–2244. [Google Scholar] [CrossRef]
- Kirjavainen, V.; Jarva, H.; Biedzka-Sarek, M.; Blom, A.M.; Skurnik, M.; Meri, S. Yersinia enterocolitica serum resistance proteins YadA and ail bind the complement regulator C4b-binding protein. PLoS Pathog. 2008, 4, e1000140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilz, D.; Vocke, T.; Heesemann, J.; Brade, V. Mechanism of YadA-mediated serum resistance of Yersinia enterocolitica serotype O3. Infect. Immun. 1992, 60, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindler, M.K.H.; Schütz, M.S.; Mühlenkamp, M.C.; Rooijakkers, S.H.M.; Hallström, T.; Zipfel, P.F.; Autenrieth, I.B. Yersinia enterocolitica YadA mediates complement evasion by recruitment and inactivation of C3 products. J. Immunol. 2012, 189, 4900–4908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlenkamp, M.C.; Hallström, T.; Autenrieth, I.B.; Bohn, E.; Linke, D.; Rinker, J.; Riesbeck, K.; Singh, B.; Leo, J.C.; Hammerschmidt, S.; et al. Vitronectin binds to a specific stretch within the head region of Yersinia adhesin a and thereby modulates Yersinia enterocolitica host interaction. J. Innate Immun. 2017, 9, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Kapach, G.; Nuri, R.; Schmidt, C.; Danin, A.; Ferrera, S.; Savidor, A.; Gerlach, R.G.; Shai, Y. Loss of the Periplasmic Chaperone Skp and mutations in the efflux pump AcrAB-TolC play a role in acquired resistance to antimicrobial peptides in Salmonella typhimurium. Front. Microbiol. 2020, 11, 189. [Google Scholar] [CrossRef] [PubMed]
- Sandkvist, M. Type II Secretion and Pathogenesis. Infect. Immun. 2001, 69, 3523–3535. [Google Scholar] [CrossRef] [Green Version]
- Otto, B.R.; van Dooren, S.J.M.; Dozois, C.M.; Luirink, J.; Oudega, B. Escherichia coli hemoglobin protease autotransporter contributes to synergistic abscess formation and heme-dependent growth of Bacteroides fragilis. Infect. Immun. 2002, 70, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Otto, B.R.; van Dooren, S.J.M.; Nuijens, J.H.; Luirink, J.; Oudega, B. Characterization of a hemoglobin protease secreted by the pathogenic Escherichia coli strain EB1. J. Exp. Med. 1998, 188, 1091–1103. [Google Scholar] [CrossRef]
- Sauri, A.; Soprova, Z.; Wickström, D.; de Gier, J.-W.; Van der Schors, R.C.; Smit, A.B.; Jong, W.S.P.; Luirink, J. The Bam (Omp85) complex is involved in secretion of the autotransporter haemoglobin protease. Microbiology 2009, 155, 3982–3991. [Google Scholar] [CrossRef] [Green Version]
- Brunder, W.; Schmidt, H.; Karch, H. EspP, a novel extracellular serine protease of enterohaemorrhagic Escherichia coli O157:H7 cleaves human coagulation factor V. Mol. Microbiol. 1997, 24, 767–778. [Google Scholar] [CrossRef]
- In, J.; Lukyanenko, V.; Foulke-Abel, J.; Hubbard, A.L.; Delannoy, M.; Hansen, A.-M.; Kaper, J.B.; Boisen, N.; Nataro, J.P.; Zhu, C.; et al. Serine protease EspP from enterohemorrhagic Escherichia coli is sufficient to induce Shiga toxin macropinocytosis in intestinal epithelium. PLoS ONE 2013, 8, e69196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziva, F.; Mahajan, A.; Cameron, P.; Currie, C.; McKendrick, I.J.; Wallis, T.S.; Smith, D.G.E.; Stevens, M.P. EspP, a Type V-secreted serine protease of enterohaemorrhagic Escherichia coli O157:H7, influences intestinal colonization of calves and adherence to bovine primary intestinal epithelial cells. FEMS Microbiol. Lett. 2007, 271, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Ieva, R.; Bernstein, H.D. Interaction of an autotransporter passenger domain with BamA during its translocation across the bacterial outer membrane. Proc. Natl. Acad. Sci. USA 2009, 106, 19120–19125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Perez, F.; Henderson, I.R.; Leyton, D.L.; Rossiter, A.E.; Zhang, Y.; Nataro, J.P. Roles of periplasmic chaperone proteins in the biogenesis of serine protease autotransporters of Enterobacteriaceae. J. Bacteriol. 2009, 191, 6571–6583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia-Pastrana, G.; Chavez-Dueñas, L.; Lanz-Mendoza, H.; Teter, K.; Navarro-García, F. VirK Is a periplasmic protein required for efficient secretion of plasmid-encoded toxin from enteroaggregative Escherichia coli. Infect. Immun. 2012, 80, 2276–2285. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, D.; Shivapriya, P.M.; Gautam, P.K.; Misra, K.; Sahoo, A.K.; Samanta, S.K. A review on basic biology of bacterial biofilm infections and their treatments by nanotechnology-based approaches. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 243–259. [Google Scholar] [CrossRef]
- Lu, R.; Sun, J.; Osei-Adjei, G.; Zhang, Y.; Huang, X. The periplasmic chaperone SurA affects motility and biofilm formation via the RcsCDB pathway in Salmonella Enterica Serovar Typhi. J. Nanosci. Nanotechnol. 2019, 19, 5503–5509. [Google Scholar] [CrossRef]
- Li, T.; Wang, J.; Cao, Q.; Li, F.; Han, J.; Zhu, B.; Zhang, Y.; Niu, H. Identification of novel genes involved in Escherichia coli persistence to tosufloxacin. Front. Cell. Infect. Microbiol. 2020, 10, 581986. [Google Scholar] [CrossRef]
- Rowley, G.; Skovierova, H.; Stevenson, A.; Rezuchova, B.; Homerova, D.; Lewis, C.; Sherry, A.; Kormanec, J.; Roberts, M. The Periplasmic Chaperone Skp is required for successful Salmonella typhimurium infection in a murine typhoid model. Microbiology 2011, 157, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Purdy, G.E.; Fisher, C.R.; Payne, S.M. IcsA surface presentation in Shigella flexneri requires the periplasmic chaperones DegP, Skp, and SurA. J. Bacteriol. 2007, 189, 5566–5573. [Google Scholar] [CrossRef]
- Sydenham, M.; Douce, G.; Bowe, F.; Ahmed, S.; Chatfield, S.; Dougan, G. Salmonella enterica serovar Typhimurium surA mutants are attenuated and effective live oral vaccines. Infect. Immun. 2000, 68, 1109–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamayo, R.; Ryan, S.S.; McCoy, A.J.; Gunn, J.S. Identification and genetic characterization of PmrA-regulated genes and genes involved in polymyxin b resistance in Salmonella enterica serovar Typhimurium. Infect. Immun. 2002, 70, 6770–6778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, S.; Rowley, G.; Stevenson, A.; Kenyon, W.J.; Spector, M.P.; Roberts, M. Role of periplasmic peptidyl-prolyl isomerases in Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 2003, 71, 5386–5388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, C.; Pathak, S.K.; Bhattacharyya, A.; Pathak, S.; Basu, J.; Kundu, M. The secreted peptidyl-prolyl cis, trans-isomerase HP0175 of Helicobacter pylori induces apoptosis of gastric epithelial cells in a TLR4- and apoptosis signal-regulating kinase 1-dependent manner. J. Immunol. 2005, 174, 5672–5680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Pathak, S.K.; Chatterjee, G.; Pathak, S.; Basu, J.; Kundu, M. Helicobacter pylori protein HP0175 transactivates epidermal growth factor receptor through TLR4 in gastric epithelial cells. J. Biol. Chem. 2008, 283, 32369–32376. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.L.; Atzingen, M.V.; Oliveira, T.R.; Oliveira, R.; Andrade, D.M.; Vasconcellos, S.A.; Nascimento, A.L.T.O. In vitro identification of novel plasminogen-binding receptors of the pathogen Leptospira interrogans. PLoS ONE 2010, 5, e11259. [Google Scholar] [CrossRef] [Green Version]
- Luu, L.D.W.; Octavia, S.; Aitken, C.; Zhong, L.; Raftery, M.J.; Sintchenko, V.; Lan, R. Surfaceome analysis of australian epidemic Bordetella pertussis reveals potential vaccine antigens. Vaccine 2020, 38, 539–548. [Google Scholar] [CrossRef]
- He, Y.; Wang, S.; Yin, X.; Sun, F.; He, B.; Liu, X. Comparison of extracellular proteins from virulent and avirulent Vibrio parahaemolyticus strains to identify potential virulence factors. J. Food Prot. 2019, 83, 155–162. [Google Scholar] [CrossRef]
- Neckers, L.; Tatu, U. Molecular chaperones in pathogen virulence: Emerging new targets for therapy. Cell Host Microbe 2008, 4, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Scheuplein, N.J.; Bzdyl, N.M.; Kibble, E.A.; Lohr, T.; Holzgrabe, U.; Sarkar-Tyson, M. Targeting protein folding: A novel approach for the treatment of pathogenic bacteria. J. Med. Chem. 2020, 63, 13355–13388. [Google Scholar] [CrossRef]
- Lehman, K.M.; Grabowicz, M. Countering Gram-negative antibiotic resistance: Recent progress in disrupting the outer membrane with novel therapeutics. Antibiotics 2019, 8, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, E.W.; Zheng, E.J.; Ryno, L.M. Identification of inhibitors of the E. coli chaperone SurA using in silico and in vitro techniques. Bioorg. Med. Chem. Lett. 2018, 28, 3540–3548. [Google Scholar] [CrossRef] [PubMed]
- Delpino, M.V.; Estein, S.M.; Fossati, C.A.; Baldi, P.C.; Cassataro, J. Vaccination with Brucella recombinant DnaK and SurA proteins induces protection against Brucella abortus infection in BALB/c mice. Vaccine 2007, 25, 6721–6729. [Google Scholar] [CrossRef] [PubMed]
- Atanassov, C.; Pezennec, L.; d’Alayer, J.; Grollier, G.; Picard, B.; Fauchère, J.-L. Novel antigens of Helicobacter pylori correspond to ulcer-related antibody pattern of sera from infected patients. J. Clin. Microbiol. 2002, 40, 547–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Chen, M.; Xu, Y.; Dong, N.; Sang, Z.; Liu, J.; Yuan, Y. Subtractive hybridization analysis of gastric diseases-associated Helicobacter pylori identifies peptidyl-prolyl isomerase as a potential marker for gastric cancer. FEMS Microbiol. Lett. 2011, 320, 103–109. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figaj, D.; Ambroziak, P.; Rzepka, I.; Skórko-Glonek, J. SurA-like and Skp-like Proteins as Important Virulence Determinants of the Gram Negative Bacterial Pathogens. Int. J. Mol. Sci. 2023, 24, 295. https://doi.org/10.3390/ijms24010295
Figaj D, Ambroziak P, Rzepka I, Skórko-Glonek J. SurA-like and Skp-like Proteins as Important Virulence Determinants of the Gram Negative Bacterial Pathogens. International Journal of Molecular Sciences. 2023; 24(1):295. https://doi.org/10.3390/ijms24010295
Chicago/Turabian StyleFigaj, Donata, Patrycja Ambroziak, Iga Rzepka, and Joanna Skórko-Glonek. 2023. "SurA-like and Skp-like Proteins as Important Virulence Determinants of the Gram Negative Bacterial Pathogens" International Journal of Molecular Sciences 24, no. 1: 295. https://doi.org/10.3390/ijms24010295