
Effect of Triazole Fungicides Titul Duo and Vintage on the Development of Pea (Pisum sativum L.) Symbiotic Nodules
 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Nodulation and Plant Growth Parameters
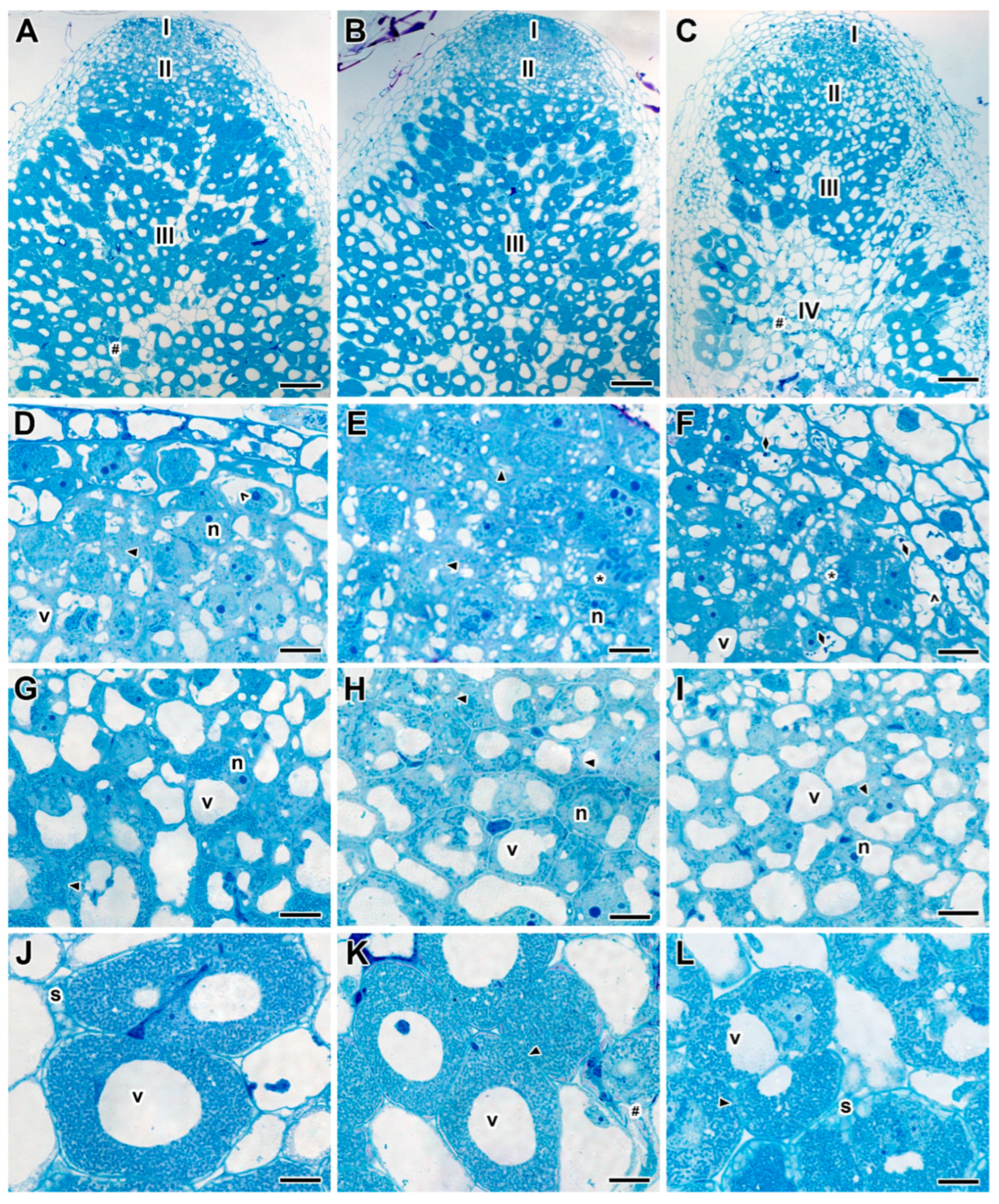
2.2. Nodule Histological Organization
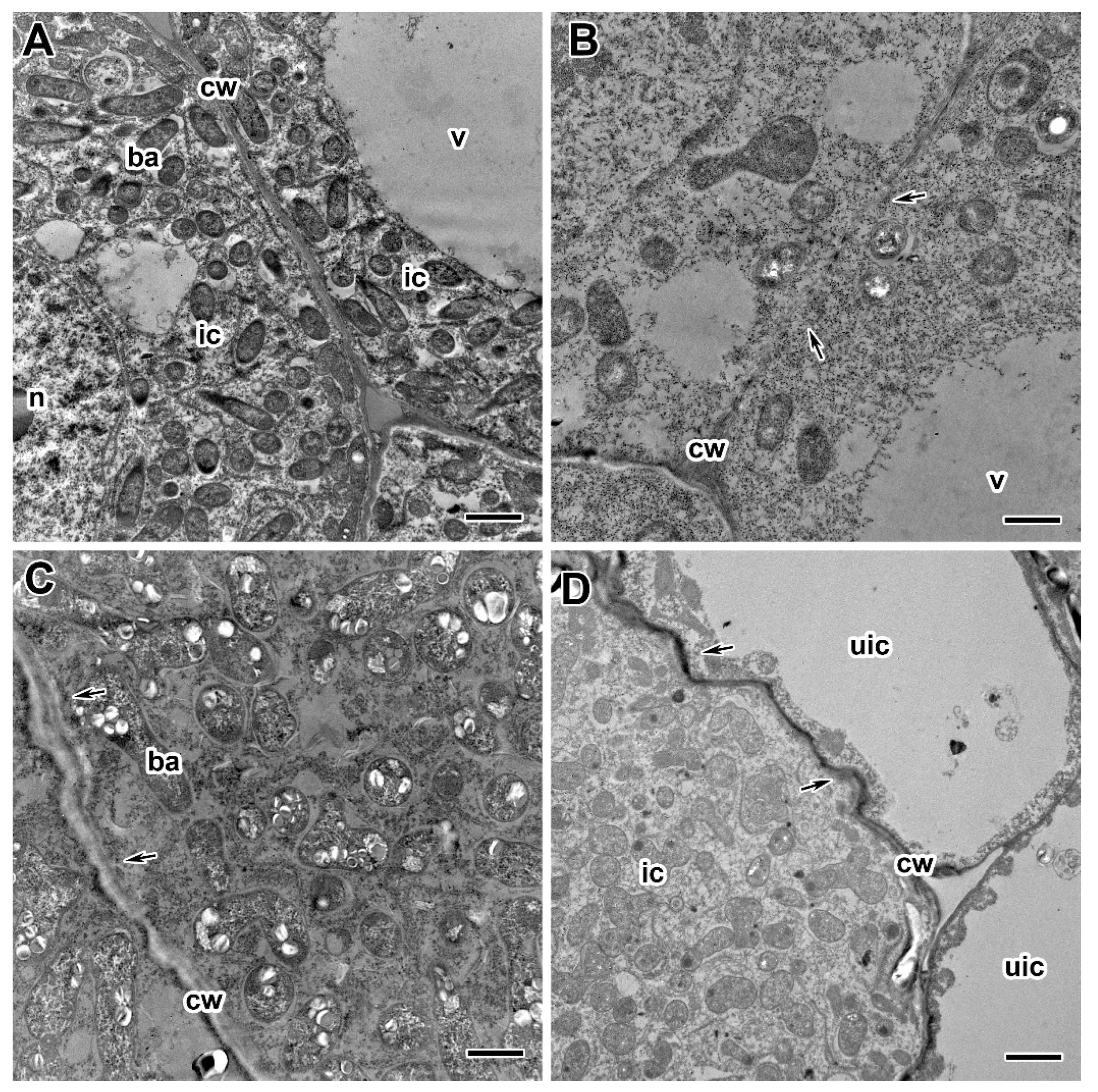
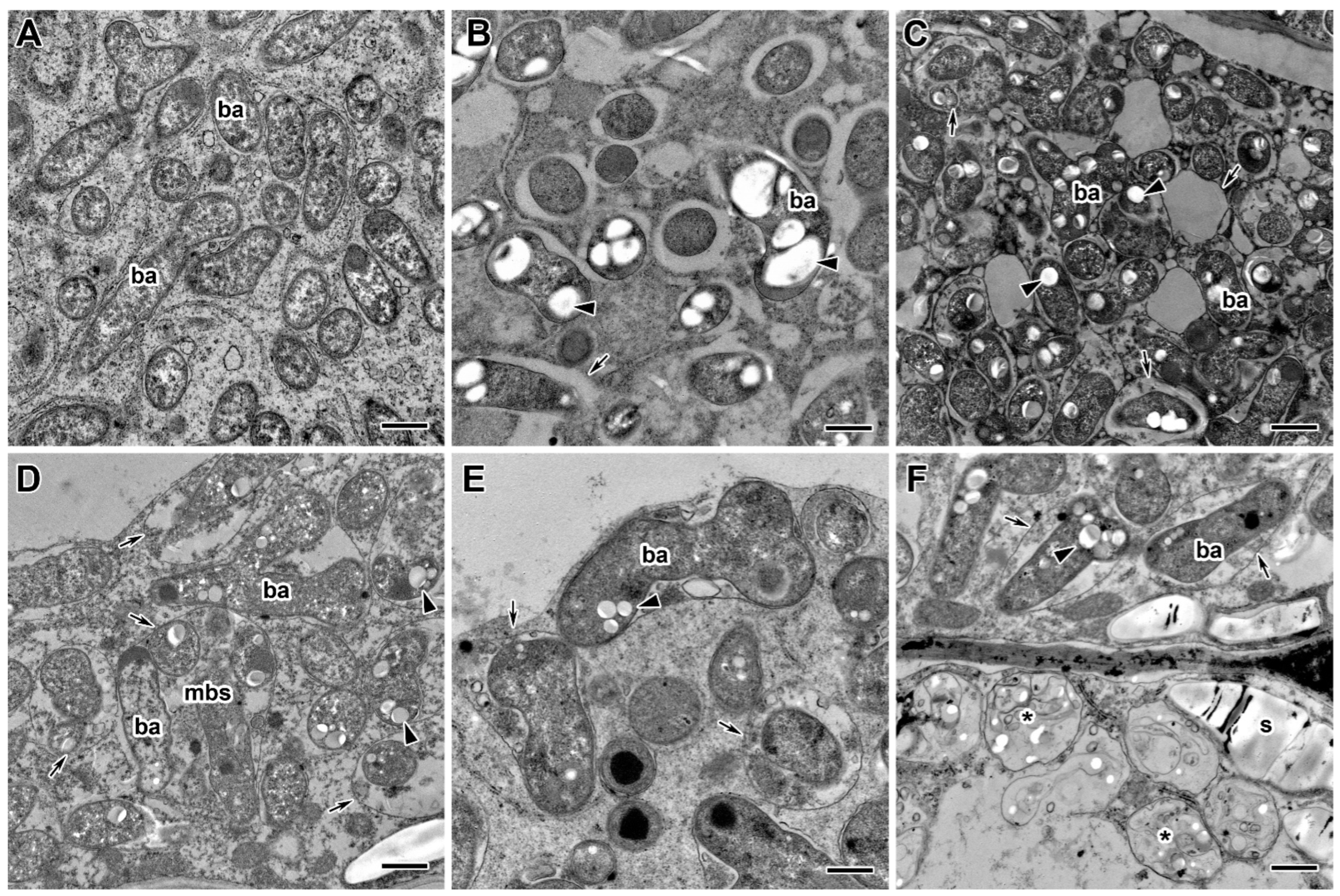
2.3. Ultrastructure of Nodules
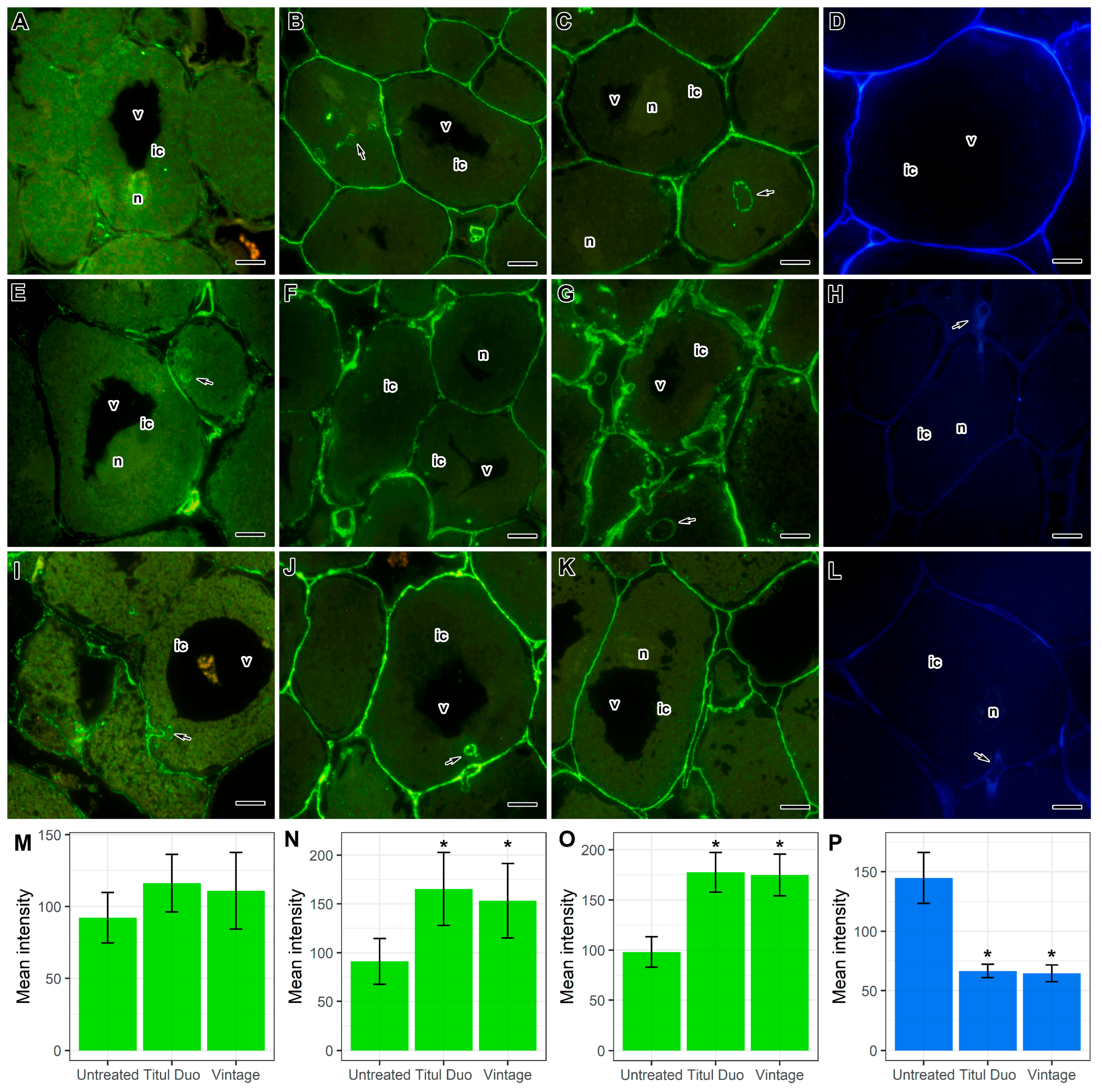
2.4. Immunocytochemical and Histochemical Analyses
2.5. Transcriptome Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Bacterial Strain
4.2. Inoculation and Plant Growth Conditions
4.3. Phenotypic Analysis of Plants and Nodules
4.4. Statistical Analysis
4.5. Electron and Light Microscopy
4.6. Fluorescence Microscopy
4.7. Transcriptomic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gullino, M.L.; Leroux, P.; Smith, C.M. Uses and challenges of novel compounds for plant disease control. Crop Prot. 2000, 19, 1–11. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and challenges in legume breeding for pest and disease resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef]
- Desai, S.; Prasad, R.; Kumar, G.P. Fusarium wilts of chickpea, pigeon pea and lentil and their management. In Microbial Interventions in Agriculture and Environment; Singh, D.P., Prabha, R., Eds.; Volume 3: Soil and Crop Health Management; Springer: Singapore, 2019; pp. 49–68. [Google Scholar] [CrossRef]
- Upadhyay, V.; Medhi, K.; Pandey, P.; Thengal, P.; Paul, S.K.; Kushwah, K. Rust disease of pea: A review. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 416–434. [Google Scholar] [CrossRef]
- Wille, L.; Messmer, M.M.; Studer, B.; Hohmann, P. Insights to plant–microbe interactions provide opportunities to improve resistance breeding against root diseases in grain legumes. Plant Cell Environ. 2019, 42, 20–40. [Google Scholar] [CrossRef]
- Williamson-Benavides, B.A.; Dhingra, A. Understanding root rot disease in agricultural crops. Horticulturae 2021, 7, 33. [Google Scholar] [CrossRef]
- Zotikov, V.; Budarina, G. Diseases of peas and the main methods of crop protection in conditions of central Russia. Plant Prot. Quar. 2015, 5, 11–15. [Google Scholar]
- Xia, X.J.; Huang, Y.Y.; Wang, L.; Huang, L.F.; Yu, Y.L.; Zhou, Y.H.; Yu, J.Q. Pesticides-induced depression of photosynthesis was alleviated by 24-epibrassinolide pretreatment in Cucumis sativus L. Pestic. Biochem. Physiol. 2006, 86, 42–48. [Google Scholar] [CrossRef]
- Schelkovo Agrohim. Available online: https://eng.betaren.ru/catalog/pesticides/fungicides/ (accessed on 23 March 2023).
- Villani, S.M.; Hulvey, J.; Hily, J.-M.; Cox, K.D. Overexpression of the CYP51A1 gene and repeated elements are associated with differential sensitivity to DMI fungicides in Venturia inaequalis. Phytopathology 2016, 106, 562–571. [Google Scholar] [CrossRef]
- Rodrigues, M.L. The multifunctional fungal ergosterol. MBio 2018, 9, e01755-18. [Google Scholar] [CrossRef]
- Sant, D.; Tupe, S.; Ramana, C.V.; Deshpande, M. Fungal cell membrane—Promising drug target for antifungal therapy. J. Appl. Microbiol. 2016, 121, 1498–1510. [Google Scholar] [CrossRef]
- Remya, V.R.; George, J.S.; Thomas, S. Manufacturing of slow- and controlled-release pesticides. In Controlled Release of Pesticides for Sustainable Agriculture; Rakhimol, K.R., Thomas, S., Volova, T., Jayachandran, K., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 127–139. [Google Scholar] [CrossRef]
- Fletcher, R.A.; Gilley, A.; Sankhla, N.; Davis, T.D. Triazoles as plant growth regulators and stress protectants. Hortic. Rev. 2000, 24, 55–138. [Google Scholar]
- Manivannan, P.; Jaleel, C.A.; Kishorekumar, A.; Sankar, B.; Somasundaram, R.; Sridharan, R.; Panneerselvam, R. Changes in antioxidant metabolism of Vigna unguiculata (L.) Walp. by propiconazole under water deficit stress. Colloids Surf. B Biointerfaces 2007, 57, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Gomathinayagam, M.; Jaleel, C.A.; Lakshmanan, G.A.; Panneerselvam, R. Changes in carbohydrate metabolism by triazole growth regulators in cassava (Manihot esculenta Crantz); effects on tuber production and quality. Comptes Rendus Biol. 2007, 330, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, V.B.; Müller, C.; Rodrigues, A.A.; Amaral, T.S.; Batista, P.F.; Silva, A.A.; Costa, A.C. Do fungicides affect the physiology, reproductive development and productivity of healthy soybean plants? Pestic. Biochem. Physiol. 2021, 172, 104754. [Google Scholar] [CrossRef] [PubMed]
- Gopi, R.; Jaleel, C.A.; Sairam, R.; Lakshmanan, G.; Gomathinayagam, M.; Panneerselvam, R. Differential effects of hexaconazole and paclobutrazol on biomass, electrolyte leakage, lipid peroxidation and antioxidant potential of Daucus carota L. Colloids Surf. B Biointerfaces 2007, 60, 180–186. [Google Scholar] [CrossRef]
- Hajihashemi, S. Physiological, biochemical, antioxidant and growth characterizations of gibberellin and paclobutrazol-treated sweet leaf (Stevia rebaudiana B.) herb. J. Plant Biochem. Biotechnol. 2018, 27, 237–240. [Google Scholar] [CrossRef]
- Kamran, M.; Ahmad, S.; Ahmad, I.; Hussain, I.; Meng, X.; Zhang, X.; Javed, T.; Ullah, M.; Ding, R.; Xu, P. Paclobutrazol application favors yield improvement of maize under semiarid regions by delaying leaf senescence and regulating photosynthetic capacity and antioxidant system during grain-filling stage. Agronomy 2020, 10, 187. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Manivannan, P.; Panneerselvam, R. Responses of antioxidant defense system of Catharanthus roseus (L.) G. Don. to paclobutrazol treatment under salinity. Acta Physiol. Plant. 2007, 29, 205–209. [Google Scholar] [CrossRef]
- Manivannan, P.; Jaleel, C.A.; Kishorekumar, A.; Sankar, B.; Somasundaram, R.; Panneerselvam, R. Protection of Vigna unguiculata (L.) Walp. plants from salt stress by paclobutrazol. Colloids Surf. B Biointerfaces 2008, 61, 315–318. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Improving tolerance against drought in canola by penconazole and calcium. Pestic. Biochem. Physiol. 2018, 149, 123–136. [Google Scholar] [CrossRef]
- Babarashi, E.; Rokhzadi, A.; Pasari, B.; Mohammadi, K. Ameliorating effects of exogenous paclobutrazol and putrescine on mung bean [Vigna radiata (L.) Wilczek] under water deficit stress. Plant Soil Environ. 2021, 67, 40–45. [Google Scholar] [CrossRef]
- Jiang, Y.; Sun, Y.; Zheng, D.; Han, C.; Cao, K.; Xu, L.; Liu, S.; Cao, Y.; Feng, N. Physiological and transcriptome analyses for assessing the effects of exogenous uniconazole on drought tolerance in hemp (Cannabis sativa L.). Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, C.; Garg, N.K.; Hasan, M.; Prathap, V.; Meena, N.L.; Singh, A.; Tyagi, A. Insight of PBZ mediated drought amelioration in crop plants. Front. Plant Sci. 2022, 13, 1008993. [Google Scholar] [CrossRef] [PubMed]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. 2017, 4, 2. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Alleviation of fungicide-induced phytotoxicity in greengram [Vigna radiata (L.) Wilczek] using fungicide-tolerant and plant growth promoting Pseudomonas strain. Saudi J. Biol. Sci. 2012, 19, 451–459. [Google Scholar] [CrossRef]
- Shahid, M.; Khan, M.S.; Kumar, M. Kitazin-pea interaction: Understanding the fungicide induced nodule alteration, cytotoxicity, oxidative damage and toxicity alleviation by Rhizobium leguminosarum. RSC Adv. 2019, 9, 16929–16947. [Google Scholar] [CrossRef]
- Ghosh, D.; Sen, S.; Mohapatra, S. Drought-mitigating Pseudomonas putida GAP-P45 modulates proline turnover and oxidative status in Arabidopsis thaliana under water stress. Ann. Microbiol. 2018, 68, 579–594. [Google Scholar] [CrossRef]
- Shahid, M.; Khan, M.S. Fungicide tolerant Bradyrhizobium japonicum mitigate toxicity and enhance greengram production under hexaconazole stress. J. Environ. Sci. 2019, 78, 92–108. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.S.; Ahmad, E.; Saif, S.; Rizvi, A.; Shahid, M. Growth stimulation and management of diseases of ornamental plants using phosphate solubilizing microorganisms: Current perspective. Acta Physiol. Plant. 2016, 38, 1–21. [Google Scholar] [CrossRef]
- Shahid, M.; Ahmed, B.; Zaidi, A.; Khan, M.S. Toxicity of fungicides to Pisum sativum: A study of oxidative damage, growth suppression, cellular death and morpho-anatomical changes. RSC Adv. 2018, 8, 38483–38498. [Google Scholar] [CrossRef]
- Shahid, M.; Khan, M.S.; Zaidi, A. Fungicide toxicity to legumes and its microbial remediation. In Pesticides in Crop Production; Srivastava, P.K., Singh, V.P., Singh, A., Tripathi, D.K., Singh, S., Prasad, S.M., Chauhan, D.K., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 2020; pp. 15–33. [Google Scholar] [CrossRef]
- Uthman, T.O.; Awosanya, A.O.; Ayanwale, A.O.S.; Ayinde, T.G. The effect of fungicides on the nodulation of cowpea (Vigna unguiculata L. Walp) in a tropical rainforest environment. J. Sustain. Dev. 2017, 7, 56–61. [Google Scholar]
- Erohin, A.I. Influence of joint application of preparations biostim maslichnyj, fungicide Titul Duo, KKR and insecticide Kinfos, CE on the productivity of peas with foliar treatment of plants. Legumes Groat Crops 2018, 1, 36. [Google Scholar] [CrossRef]
- Erokhin, A.I.; Tsukanova, Z.R.; Latyntseva, E.V. Prospects for the use of liquid peat potassium humate and fungicide Titul Duo, KKR for foliar treatment of pea plants. Legumes Groat Crops 2020, 3, 32–36. [Google Scholar] [CrossRef]
- Rezvyakova, S.B.; Eryomin, L.P. Increasing soybean yield based on protection against fungal diseases. Bull. Agric. Sci. 2021, 3, 77–83. [Google Scholar] [CrossRef]
- Chakrabarty, M.; Hossen, F.; Begum, A.; Akhter, H. Plant growth promoting (PGP) activities of Rhizobial isolates from Sesbania bispinosa in response to pesticides. Bangladesh J. Med. Microbiol. 2021, 38, 31–37. [Google Scholar] [CrossRef]
- Erokhin, A.I. Effectiveness of foliar (leaf) treatment of pea plants with the preparation Humate+7. Legumes and Groat Crops 2022, 1, 55–60. [Google Scholar] [CrossRef]
- Ahemad, M. A comparative analysis of Tebuconazole mediated phytotoxicity to legumes. J. Environ. Sci. Technol. 2011, 4, 630–637. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Effect of tebuconazole-tolerant and plant growth promoting Rhizobium isolate MRP1 on pea–Rhizobium symbiosis. Sci. Hortic. 2011, 129, 266–272. [Google Scholar] [CrossRef]
- Mamenko, T.P.; Kots, S.Y.; Khomenko, Y.O. The intensity of ethylene release by soybean plants under the influence of fungicides in the early stages of legume-rhizobial symbiosis. Regul. Mech. Biosyst. 2020, 11, 98–104. [Google Scholar] [CrossRef]
- You, M.P.; Lamichhane, J.R.; Aubertot, J.-N.; Barbetti, M.J. Understanding why effective fungicides against individual soilborne pathogens are ineffective with soilborne pathogen complexes. Plant Dis. 2020, 104, 904–920. [Google Scholar] [CrossRef]
- Roman, D.L.; Voiculescu, D.I.; Filip, M.; Ostafe, V.; Isvoran, A. Effects of triazole fungicides on soil microbiota and on the activities of enzymes found in soil: A review. Agriculture 2021, 11, 893. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Productivity of greengram in tebuconazole-stressed soil, by using a tolerant and plant growth-promoting Bradyrhizobium sp. MRM6 strain. Acta Physiol. Plant. 2012, 34, 245–254. [Google Scholar] [CrossRef]
- Tsyganov, V.E.; Morzhina, E.V.; Stefanov, S.Y.; Borisov, A.Y.; Lebsky, V.K.; Tikhonovich, I.A. The pea (Pisum sativum L.) genes sym33 and sym40 control infection thread formation and root nodule function. Mol. Gen. Genet. 1998, 259, 491–503. [Google Scholar] [CrossRef]
- Gorshkov, A.P.; Tsyganova, A.V.; Vorobiev, M.G.; Tsyganov, V.E. The fungicide tetramethylthiuram disulfide negatively affects plant cell walls, infection thread walls, and symbiosomes in pea (Pisum sativum L.) symbiotic nodules. Plants 2020, 9, 1488. [Google Scholar] [CrossRef] [PubMed]
- Jeyakumar, P.; Boominathan, P. Combined effect of trifloxystrobin and tebuconazole on growth and yield attributes of groundnut. Legume Res. 2016, 39, 605–609. [Google Scholar] [CrossRef]
- Machado, F.J.; Santana, F.M.; Lau, D.; Del Ponte, E.M. Quantitative review of the effects of triazole and benzimidazole fungicides on Fusarium head blight and wheat yield in Brazil. Plant Dis. 2017, 101, 1633–1641. [Google Scholar] [CrossRef]
- Tak, P.S.; Pannu, P.P.S.; Kaur, J. Effect of foliar application of fungicides, Neem extract and Butter Milk on yellow rust, plant growth and yield parameters of wheat. Agric. Res. J. 2021, 58, 232–239. [Google Scholar] [CrossRef]
- Kang, Z.; Huang, L.; Krieg, U.; Mauler-Machnik, A.; Buchenauer, H. Effects of tebuconazole on morphology, structure, cell wall components and trichothecene production of Fusarium culmorum in vitro. Pest Manag. Sci. 2001, 57, 491–500. [Google Scholar] [CrossRef]
- Li, N.; Zhao, J.; Zhang, R.; Deng, L.; Li, J.; Gao, Y.; Liu, C. Effect of tebuconazole enantiomers and environmental factors on fumonisin accumulation and FUM gene expression in Fusarium verticillioides. J. Agric. Food Chem. 2018, 66, 13107–13115. [Google Scholar] [CrossRef]
- Li, C.; Fan, S.; Wen, Y.; Tan, Z.; Liu, C. Enantioselective effect of flutriafol on growth, deoxynivalenol production, and TRI gene transcript levels in Fusarium graminearum. J. Agric. Food Chem. 2021, 69, 1684–1692. [Google Scholar] [CrossRef]
- Rafique, T.; Naseem, S.; Usmani, T.H.; Bashir, E.; Khan, F.A.; Bhanger, M.I. Geochemical factors controlling the occurrence of high fluoride groundwater in the Nagar Parkar area, Sindh, Pakistan. J. Hazard. Mater. 2009, 171, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Cantonwine, E.G. Post-infection activities of fungicides against Cercospora arachidicola of peanut (Arachis hypogaea). Pest Manag. Sci. 2014, 70, 1202–1206. [Google Scholar] [CrossRef] [PubMed]
- Melloy, P.; Del Ponte, E.M.; Sparks, A.H. Timing of triazole-based spray schedules for managing mungbean powdery mildew in Australia: A meta-analysis. Plant Dis. 2022, 106, 918–924. [Google Scholar] [CrossRef]
- Vasić, T.; Vojinović, U.; Žujović, S.; Krnjaja, V.; Živković, S.; Marković, J.; Stević, M. In vitro toxicity of fungicides with different modes of action to alfalfa anthracnose fungus, Colletotrichum destructivum. J. Environ. Sci. Health B 2019, 54, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Hussain, S.; Bano, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.A.; Khan, F.; Chen, Y.; Wu, C. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: Consequences for changing environment. Environ. Sci. Pollut. Res. 2015, 22, 4907–4921. [Google Scholar] [CrossRef]
- Liu, C.; Feng, N.; Zheng, D.; Cui, H.; Sun, F.; Gong, X. Uniconazole and diethyl aminoethyl hexanoate increase soybean pod setting and yield by regulating sucrose and starch content. J. Sci. Food Agric. 2019, 99, 748–758. [Google Scholar] [CrossRef]
- Karikalan, L.; Rajan, S.; Gopi, R.; Sujatha, B.; Panneerselvam, R. Induction of salt tolerants by triadimefon in pigeon pea (Cajanus cajan L.) Millsp. Indian J Exp. Biol. 1999, 37, 825–829. [Google Scholar]
- Hu, H.; Feng, N.; Shen, X.; Zhao, L.; Zheng, D. Transcriptomic analysis of Vigna radiata in response to chilling stress and uniconazole application. BMC Genom. 2022, 23, 205. [Google Scholar] [CrossRef]
- Petit, A.-N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant-Gaveau, N. Fungicide impacts on photosynthesis in crop plants. Photosynth. Res. 2012, 111, 315–326. [Google Scholar] [CrossRef]
- Zhu, L.-H.; van de Peppel, A.; Li, X.-Y.; Welander, M. Changes of leaf water potential and endogenous cytokinins in young apple trees treated with or without paclobutrazol under drought conditions. Sci. Hortic. 2004, 99, 133–141. [Google Scholar] [CrossRef]
- Zhang, M.; Duan, L.; Tian, X.; He, Z.; Li, J.; Wang, B.; Li, Z. Uniconazole-induced tolerance of soybean to water deficit stress in relation to changes in photosynthesis, hormones and antioxidant system. J. Plant Physiol. 2007, 164, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R.; Hofstra, G. Triazoles as potential plant protectants. In Sterol Biosynthesis Inhibitors in Plant Protection; Berg, D., Plempel, M., Eds.; Ellis Horwood Ltd.: Cambridge, UK, 1988; pp. 31–331. [Google Scholar]
- Pal, S.; Zhao, J.; Khan, A.; Yadav, N.S.; Batushansky, A.; Barak, S.; Rewald, B.; Fait, A.; Lazarovitch, N.; Rachmilevitch, S. Paclobutrazol induces tolerance in tomato to deficit irrigation through diversified effects on plant morphology, physiology and metabolism. Sci. Rep. 2016, 6, 39321. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-H.; Lin, C.-H. The effect of paclobutrazol on physiological and biochemical changes in the primary roots of pea. J. Exp. Bot. 1992, 43, 1367–1372. [Google Scholar] [CrossRef]
- Shishatskaya, E.; Menzyanova, N.; Zhila, N.; Prudnikova, S.; Volova, T.; Thomas, S. Toxic effects of the fungicide tebuconazole on the root system of fusarium-infected wheat plants. Plant Physiol. Biochem. 2018, 132, 400–407. [Google Scholar] [CrossRef]
- Svobodníková, L.; Kummerová, M.; Zezulka, Š.; Babula, P.; Sendecká, K. Root response in Pisum sativum under naproxen stress: Morphoanatomical, cytological, and biochemical traits. Chemosphere 2020, 258, 127411. [Google Scholar] [CrossRef] [PubMed]
- Kummerová, M.; Zezulka, Š.; Babula, P.; Váňová, L. Root response in Pisum sativum and Zea mays under fluoranthene stress: Morphological and anatomical traits. Chemosphere 2013, 90, 665–673. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. R. 1999, 63, 968–989. [Google Scholar] [CrossRef]
- Gaind, S.; Rathi, M.S.; Kaushik, B.D.; Nain, L.; Verma, O.P. Survival of bio-inoculants on fungicides-treated seeds of wheat, pea and chickpea and subsequent effect on chickpea yield. J. Environ. Sci. Health B 2007, 42, 663–668. [Google Scholar] [CrossRef]
- Yang, C.; Hamel, C.; Vujanovic, V.; Gan, Y. Nontarget effects of foliar fungicide application on the rhizosphere: Diversity of nifH gene and nodulation in chickpea field. J. Appl. Microbiol. 2012, 112, 966–974. [Google Scholar] [CrossRef]
- Dhungana, S.K.; Kim, I.-D.; Kwak, H.-S.; Shin, D.-H. Unraveling the effect of structurally different classes of insecticide on germination and early plant growth of soybean [Glycine max (L.) Merr.]. Pestic. Biochem. Physiol. 2016, 130, 39–43. [Google Scholar] [CrossRef]
- Schneider, M.; Keiblinger, K.M.; Paumann, M.; Soja, G.; Mentler, A.; Golestani-Fard, A.; Retzmann, A.; Prohaska, T.; Zechmeister-Boltenstern, S.; Wenzel, W. Fungicide application increased copper-bioavailability and impaired nitrogen fixation through reduced root nodule formation on alfalfa. Ecotoxicology 2019, 28, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Lal, A.A.; Simon, S. Efficacy of botanicals and bio-agents against powdery mildew disease of garden pea (Pisum sativum L.). J. Pharmacogn. Phytochem. 2017, 6, 1125–1126. [Google Scholar]
- Ju, C.; Xu, J.; Wu, X.; Dong, F.; Liu, X.; Tian, C.; Zheng, Y. Effects of hexaconazole application on soil microbes community and nitrogen transformations in paddy soils. Sci. Total Environ. 2017, 609, 655–663. [Google Scholar] [CrossRef]
- Kengar, Y.D.; Patil, B.J. Ureide content of guar under influence of hexaconazole and triazophos. J. Fertil. Pestic 2017, 8, 176. [Google Scholar] [CrossRef]
- Li, Y.Y.; Yang, H. Bioaccumulation and degradation of pentachloronitrobenzene in Medicago sativa. J. Environ. Manag. 2013, 119, 143–150. [Google Scholar] [CrossRef]
- Yakubu, H.; Buji, I.; Sandabe, M. Effects of seed-dressing fungicides on germination, nodulation, N2-fixation and yields of two groundnut varieties in semi-arid region of Nigeria. Int. J. Appl. Agric. Res. 2011, 6, 121–129. [Google Scholar]
- Zemenu, E.A.; Gizachew, H.B.; Ahmed, M.Y. Evaluations of different seed-dressing fungicides with chickpea varieties for the management of fusarium wilt in Eastern Amhara, Ethiopia. Adv. Agric. 2022, 2022, 9324559. [Google Scholar] [CrossRef]
- Mourad, B.; Baha-Eddine, B.; Mokhtar, B. The responses of the antioxidant defence system of a legume green bean Phaseolus Vulgaris cv. Djedida, exposed to a xenobiotic hexaconazole. Int. J. Adv. Eng. Manag. 2017, 2, 270–278. [Google Scholar] [CrossRef]
- Sankar, B.; Jaleel, C.A.; Manivannan, P.; Kishorekumar, A.; Somasundaram, R.; Panneerselvam, R. Effect of paclobutrazol on water stress amelioration through antioxidants and free radical scavenging enzymes in Arachis hypogaea L. Colloids Surf. B Biointerfaces 2007, 60, 229–235. [Google Scholar] [CrossRef]
- Liu, R.; Li, J.; Zhang, L.; Feng, T.; Zhang, Z.; Zhang, B. Fungicide difenoconazole induced biochemical and developmental toxicity in wheat (Triticum aestivum L.). Plants 2021, 10, 2304. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Baird, L.M.; Escuredo, P.R.; Dalton, D.A.; Minchin, F.R.; Iturbe-Ormaetxe, I.; Rubio, M.C.; Moran, J.F.; Gordon, A.J.; Becana, M. Stress-induced legume root nodule senescence. Physiological, biochemical, and structural alterations. Plant Physiol. 1999, 121, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Tsyganova, A.V.; Seliverstova, E.V.; Tsyganov, V.E. Influence of mutation in pea (Pisum sativum L.) cdt (cadmium tolerance) gene on histological and ultrastructural nodule organization. Ecol. Genet. 2019, 17, 71–80. [Google Scholar] [CrossRef]
- Tsyganov, V.E.; Tsyganova, A.V.; Gorshkov, A.P.; Seliverstova, E.V.; Kim, V.E.; Chizhevskaya, E.P.; Belimov, A.A.; Serova, T.A.; Ivanova, K.A.; Kulaeva, O.A. Efficacy of a plant-microbe system: Pisum sativum (L.) cadmium-tolerant mutant and Rhizobium leguminosarum strains, expressing pea metallothionein genes PsMT1 and PsMT2, for cadmium phytoremediation. Front. Microbiol. 2020, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Dhanushkodi, R.; Matthew, C.; McManus, M.T.; Dijkwel, P.P. Drought-induced senescence of Medicago truncatula nodules involves serpin and ferritin to control proteolytic activity and iron levels. New Phytol. 2018, 220, 196–208. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef]
- Sánchez-Pardo, B.; Fernández-Pascual, M.; Zornoza, P. Copper microlocalisation, ultrastructural alterations and antioxidant responses in the nodules of white lupin and soybean plants grown under conditions of copper excess. Environ. Exp. Bot. 2012, 84, 52–60. [Google Scholar] [CrossRef]
- de María, N.; de Felipe, M.R.; Fernández-Pascual, M. Alterations induced by glyphosate on lupin photosynthetic apparatus and nodule ultrastructure and some oxygen diffusion related proteins. Plant Physiol. Biochem. 2005, 43, 985–996. [Google Scholar] [CrossRef]
- Bolaños, L.; El-Hamdaoui, A.; Bonilla, I. Recovery of development and functionality of nodules and plant growth in salt-stressed Pisum sativum-Rhizobium leguminosarum symbiosis by boron and calcium. J. Plant Physiol. 2003, 160, 1493–1497. [Google Scholar] [CrossRef]
- Wilmowicz, E.; Kućko, A.; Alché, J.D.D.; Czeszewska-Rosiak, G.; Florkiewicz, A.B.; Kapusta, M.; Karwaszewski, J. Remodeling of cell wall components in root nodules and flower abscission zone under drought in yellow lupine. Int. J. Mol. Sci. 2022, 23, 1680. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Kasowska, D.; Gediga, K.; Banasiewicz, J.; Stępkowski, T. Lotus corniculatus-rhizobia symbiosis under Ni, Co and Cr stress on ultramafic soil. Plant Soil 2020, 451, 459–484. [Google Scholar] [CrossRef]
- Díaz-Cacho, P.; Moral, R.; Encina, A.; Luis Acebes, J.; Alvarez, J. Cell wall modifications in bean (Phaseolus vulgaris) callus cultures tolerant to isoxaben. Physiol. Plant. 1999, 107, 54–59. [Google Scholar] [CrossRef]
- Manfield, I.W.; Orfila, C.; McCartney, L.; Harholt, J.; Bernal, A.J.; Scheller, H.V.; Gilmartin, P.M.; Mikkelsen, J.D.; Paul Knox, J.; Willats, W.G. Novel cell wall architecture of isoxaben-habituated Arabidopsis suspension-cultured cells: Global transcript profiling and cellular analysis. Plant J. 2004, 40, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, C.; Bilkova, A.; Jackson, P.; Dabravolski, S.; Riber, W.; Didi, V.; Houser, J.; Gigli-Bisceglia, N.; Wimmerova, M.; Budínská, E. Dirigent proteins in plants: Modulating cell wall metabolism during abiotic and biotic stress exposure. J. Exp. Bot. 2017, 68, 3287–3301. [Google Scholar] [CrossRef] [PubMed]
- Coculo, D.; Lionetti, V. The plant invertase/pectin methylesterase inhibitor superfamily. Front. Plant Sci. 2022, 13, 863892. [Google Scholar] [CrossRef]
- Redondo-Nieto, M.; Pulido, L.; Reguera, M.; Bonilla, I.; Bolaños, L. Developmentally regulated membrane glycoproteins sharing antigenicity with rhamnogalacturonan II are not detected in nodulated boron deficient Pisum sativum. Plant Cell Environ. 2007, 30, 1436–1443. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Borucki, W. Pectins esterification in the apoplast of aluminum-treated pea root nodules. J. Plant Physiol. 2015, 184, 1–7. [Google Scholar] [CrossRef]
- Ivanova, K.A.; Tsyganova, A.V.; Brewin, N.J.; Tikhonovich, I.A.; Tsyganov, V.E. Induction of host defences by Rhizobium during ineffective nodulation of pea (Pisum sativum L.) carrying symbiotically defective mutations sym40 (PsEFD), sym33 (PsIPD3/PsCYCLOPS) and sym42. Protoplasma 2015, 252, 1505–1517. [Google Scholar] [CrossRef]
- Tsyganova, A.V.; Tsyganov, V.E. Plant cell wall in symbiotic interactions. Pectins. Agric. Biol. 2019, 446. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Borucki, W.; Znojek, E. Structural changes in Medicago truncatula root nodules caused by short-term aluminum stress. Symbiosis 2012, 58, 161–170. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Borucki, W. Localization of hydrogen peroxide accumulation and diamine oxidase activity in pea root nodules under aluminum stress. Micron 2014, 57, 13–22. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Ważny, R. Metal resistant rhizobia and ultrastructure of Anthyllis vulneraria nodules from zinc and lead contaminated tailing in Poland. Int. J. Phytoremediat. 2018, 20, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Tsyganova, A.V.; Seliverstova, E.V.; Brewin, N.J.; Tsyganov, V.E. Bacterial release is accompanied by ectopic accumulation of cell wall material around the vacuole in nodules of Pisum sativum sym33-3 allele encoding transcription factor PsCYCLOPS/PsIPD3. Protoplasma 2019, 256, 1449–1453. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Díez, B.; Quiñones, M.A.; Fajardo, S.; Morcillo, C.; Fernández-Pascual, M. Possible reasons for tolerance to mercury of Lupinus albus cv. G1 inoculated with Hg-resistant and sensitive Bradyrhizobium canariense strains. Symbiosis 2015, 67, 91–102. [Google Scholar] [CrossRef]
- Trainer, M.A.; Charles, T.C. The role of PHB metabolism in the symbiosis of rhizobia with legumes. Appl. Microbiol. Biotechnol. 2006, 71, 377–386. [Google Scholar] [CrossRef]
- Borucki, W.; Sujkowska, M. The effects of sodium chloride-salinity upon growth, nodulation, and root nodule structure of pea (Pisum sativum L.) plants. Acta Physiol. Plant. 2008, 30, 293–301. [Google Scholar] [CrossRef]
- Tsyganova, A.V.; Kitaeva, A.B.; Gorshkov, A.P.; Kusakin, P.G.; Sadovskaya, A.R.; Borisov, Y.G.; Tsyganov, V.E. Glycyrrhiza uralensis nodules: Histological and ultrastructural organization and tubulin cytoskeleton dynamics. Agronomy 2021, 11, 2508. [Google Scholar] [CrossRef]
- Pankhurst, C.E.; Craig, A.S.; Jones, W.T. Effectiveness of Lotus root nodules: I. Morphology and flavolan content of nodules formed on Lotus pedunculatus by fast-growing Lotus rhizobia. J. Exp. Bot. 1979, 30, 1085–1093. [Google Scholar] [CrossRef]
- Li, X.; Bao, H.; Wang, Z.; Wang, M.; Fan, B.; Zhu, C.; Chen, Z. Biogenesis and function of multivesicular bodies in plant immunity. Front. Plant Sci. 2018, 9, 979. [Google Scholar] [CrossRef]
- Wang, M.; Li, X.; Luo, S.; Fan, B.; Zhu, C.; Chen, Z. Coordination and crosstalk between autophagosome and multivesicular body pathways in plant stress responses. Cells 2020, 9, 119. [Google Scholar] [CrossRef]
- Krishnan, H.B.; Kim, W.-S.; Sun-Hyung, J.; Kim, K.Y.; Jiang, G. Citrate synthase mutants of Sinorhizobium fredii USDA257 form ineffective nodules with aberrant ultrastructure. Appl. Environ. Microbiol. 2003, 69, 3561–3568. [Google Scholar] [CrossRef]
- Engvild, K.C. Nodulation and nitrogen fixation mutants of pea, Pisum sativum. Theor. Appl. Genet. 1987, 74, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.S. Variation in nitrate tolerance of nitrogen fixation in the pea/Rhizobium symbiosis. Plant Breed. 1987, 98, 130–135. [Google Scholar] [CrossRef]
- Glenn, A.; Poole, P.; Hudman, J. Succinate uptake by free-living and bacteroid forms of Rhizobium leguminosarum. Microbiology 1980, 119, 267–271. [Google Scholar] [CrossRef]
- Beringer, J.E. R factor transfer in Rhizobium leguminosarum. Microbiology 1974, 84, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Fåhraeus, G. The infection of clover root hairs by nodule bacteria studied by a simple glass slide technique. J. Gen. Microbiol. 1957, 16, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Serova, T.A.; Tsyganova, A.V.; Tsyganov, V.E. Early nodule senescence is activated in symbiotic mutants of pea (Pisum sativum L.) forming ineffective nodules blocked at different nodule developmental stages. Protoplasma 2018, 255, 1443–1459. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, C.D.; Pittman, F.E. A simple methylene blue-azure II-basic fuchsin stain for epoxy-embedded tissue sections. Stain. Technol. 1974, 49, 9–14. [Google Scholar] [CrossRef]
- Musielak, T.J.; Schenkel, L.; Kolb, M.; Henschen, A.; Bayer, M. A simple and versatile cell wall staining protocol to study plant reproduction. Plant Reprod. 2015, 28, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Verhertbruggen, Y.; Marcus, S.E.; Haeger, A.; Ordaz-Ortiz, J.J.; Knox, J.P. An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr. Res. 2009, 344, 1858–1862. [Google Scholar] [CrossRef]
- Liners, F.; Letesson, J.-J.; Didembourg, C.; Van Cutsem, P. Monoclonal antibodies against pectin: Recognition of a conformation induced by calcium. Plant Physiol. 1989, 91, 1419–1424. [Google Scholar] [CrossRef]
- Pattathil, S.; Avci, U.; Baldwin, D.; Swennes, A.G.; McGill, J.A.; Popper, Z.; Bootten, T.; Albert, A.; Davis, R.H.; Chennareddy, C. A comprehensive toolkit of plant cell wall glycan-directed monoclonal antibodies. Plant Physiol. 2010, 153, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Kusakin, P.G.; Serova, T.A.; Gogoleva, N.E.; Gogolev, Y.V.; Tsyganov, V.E. Laser microdissection of Pisum sativum L. nodules followed by RNA-Seq analysis revealed crucial transcriptomic changes during infected cell differentiation. Agronomy 2021, 11, 2504. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorshkov, A.P.; Kusakin, P.G.; Borisov, Y.G.; Tsyganova, A.V.; Tsyganov, V.E. Effect of Triazole Fungicides Titul Duo and Vintage on the Development of Pea (Pisum sativum L.) Symbiotic Nodules. Int. J. Mol. Sci. 2023, 24, 8646. https://doi.org/10.3390/ijms24108646
Gorshkov AP, Kusakin PG, Borisov YG, Tsyganova AV, Tsyganov VE. Effect of Triazole Fungicides Titul Duo and Vintage on the Development of Pea (Pisum sativum L.) Symbiotic Nodules. International Journal of Molecular Sciences. 2023; 24(10):8646. https://doi.org/10.3390/ijms24108646
Chicago/Turabian StyleGorshkov, Artemii P., Pyotr G. Kusakin, Yaroslav G. Borisov, Anna V. Tsyganova, and Viktor E. Tsyganov. 2023. "Effect of Triazole Fungicides Titul Duo and Vintage on the Development of Pea (Pisum sativum L.) Symbiotic Nodules" International Journal of Molecular Sciences 24, no. 10: 8646. https://doi.org/10.3390/ijms24108646