
Pro-Osteogenic and Anti-Inflammatory Synergistic Effect of Orthosilicic Acid, Vitamin K2, Curcumin, Polydatin and Quercetin Combination in Young and Senescent Bone Marrow-Derived Mesenchymal Stromal Cells
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
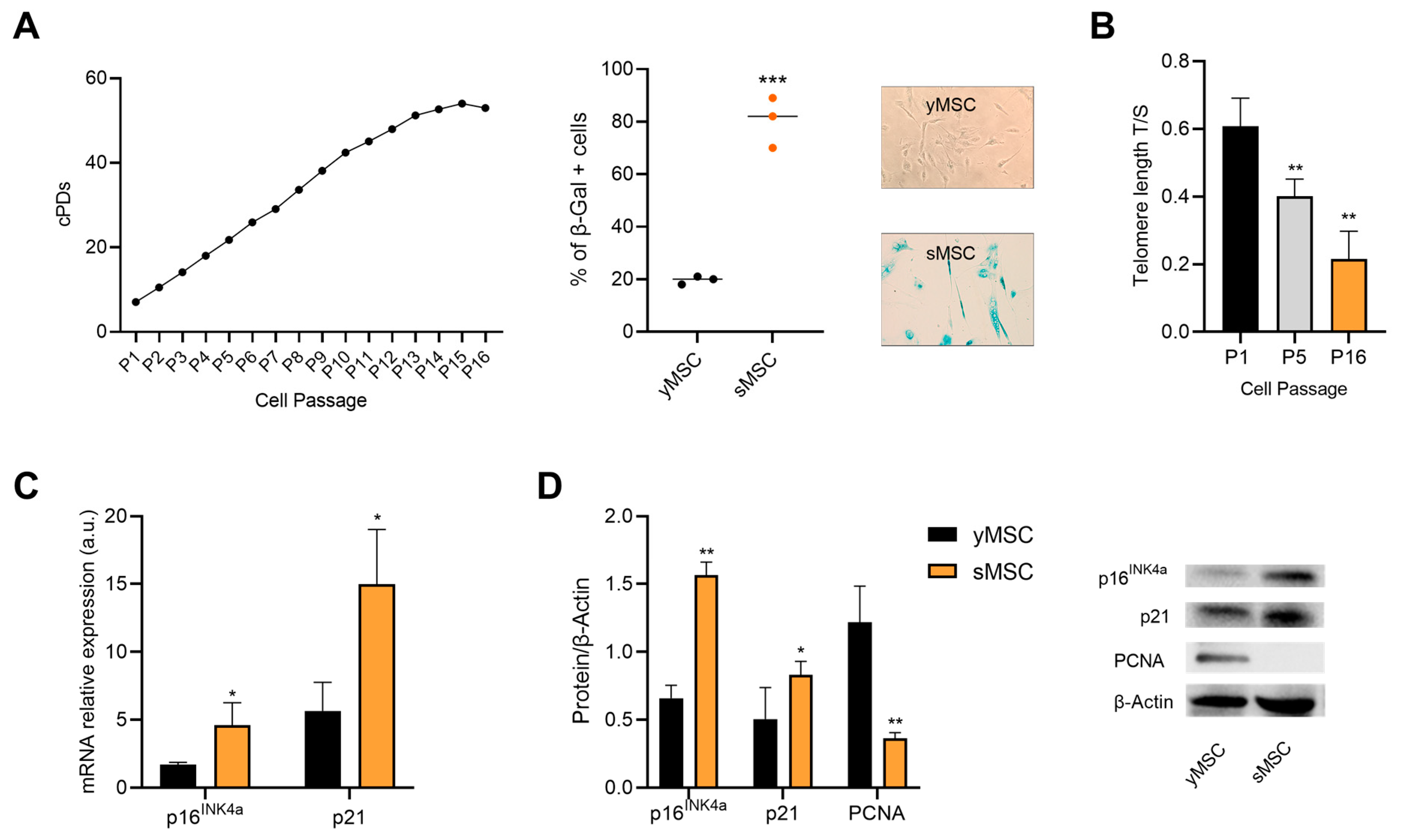
2.1. Replicative Senescence in Human Bone Marrow Mesenchymal Stromal Cells
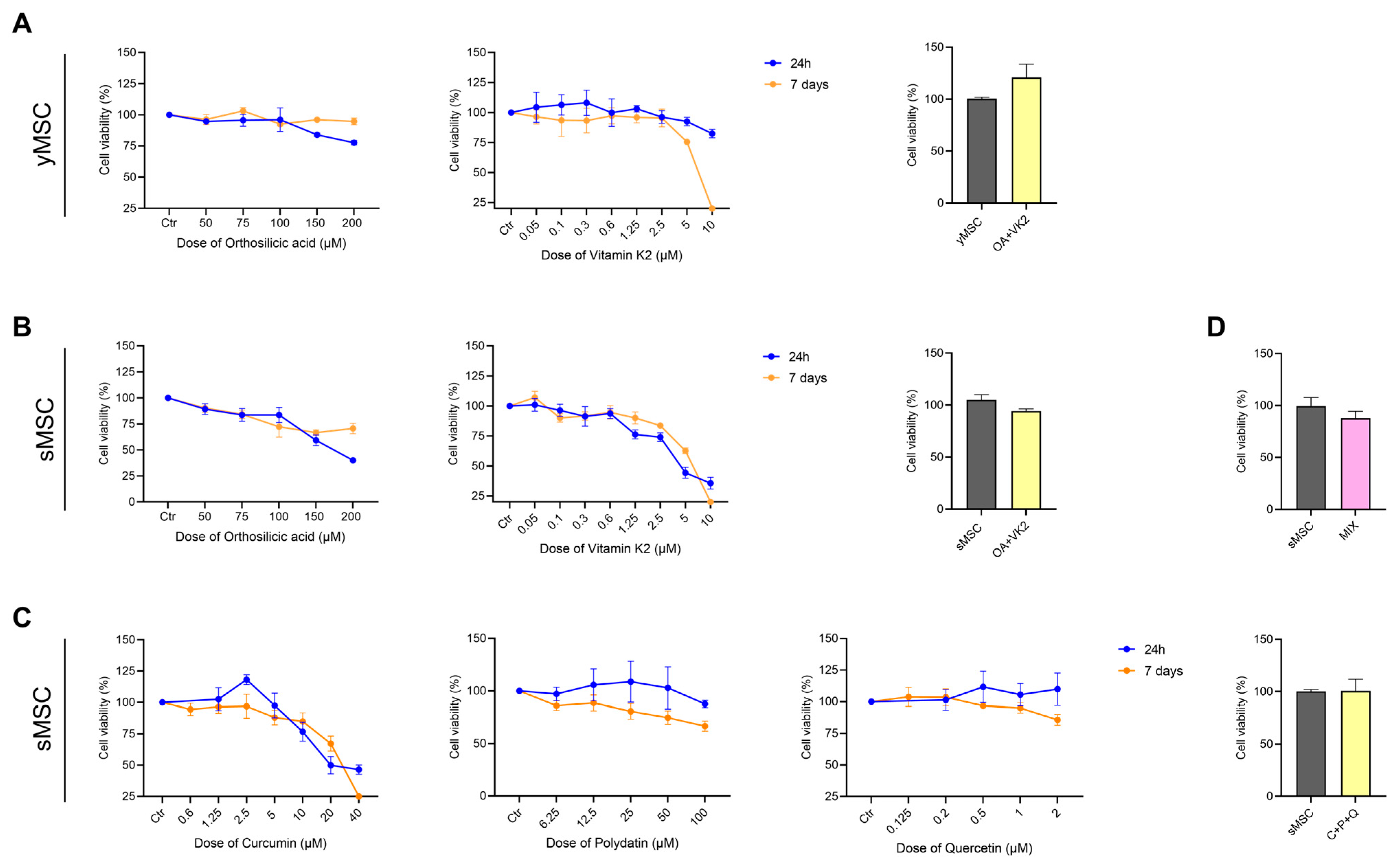
2.2. Orthosilicic Acid, Vitamin K2, Curcumin, Polydatin and Quercetin Effect on yMSC and sMSC Cell Viability
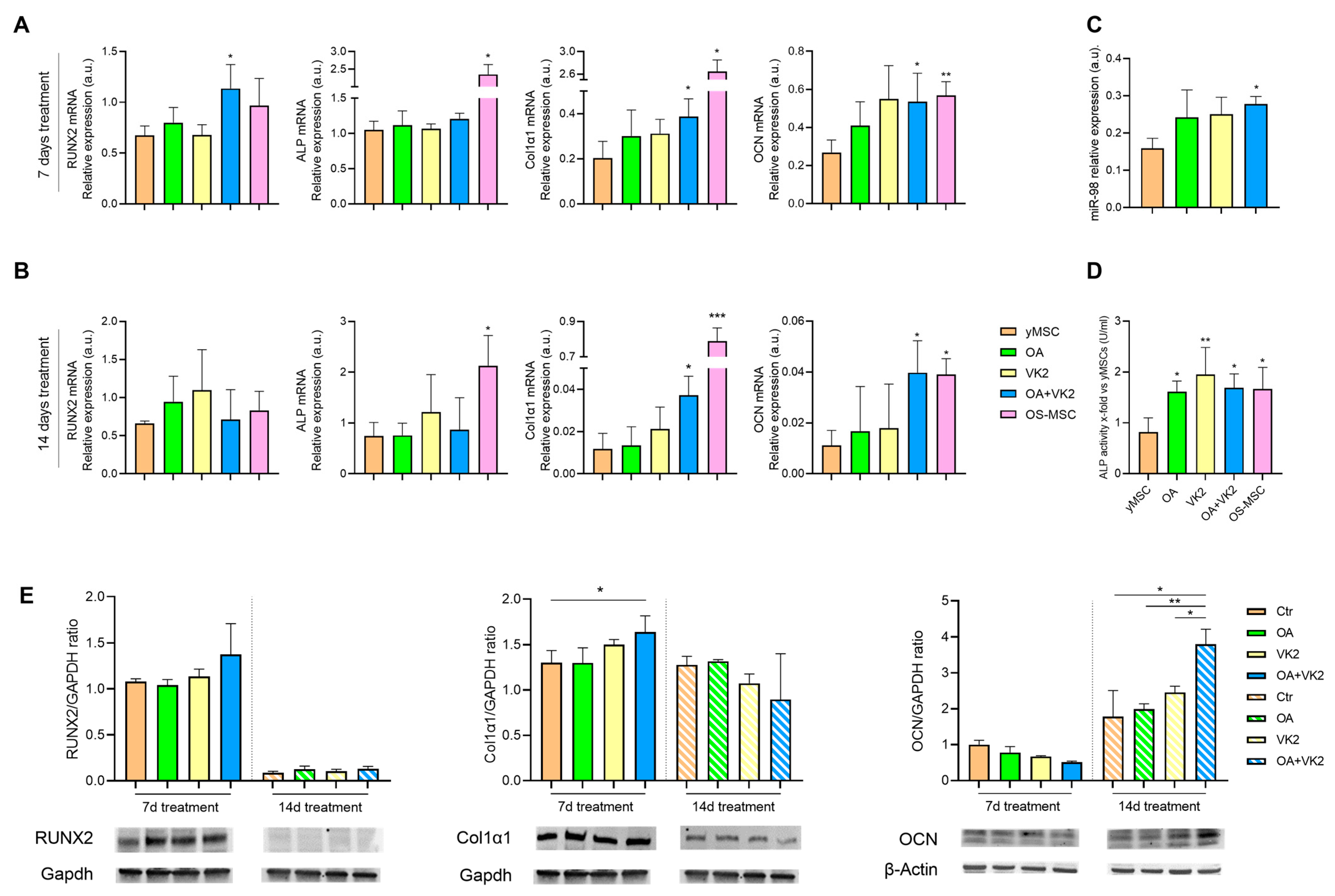
2.3. Orthosilicic Acid and Vitamin K2 Have a Mineralizing Effect on yMSC
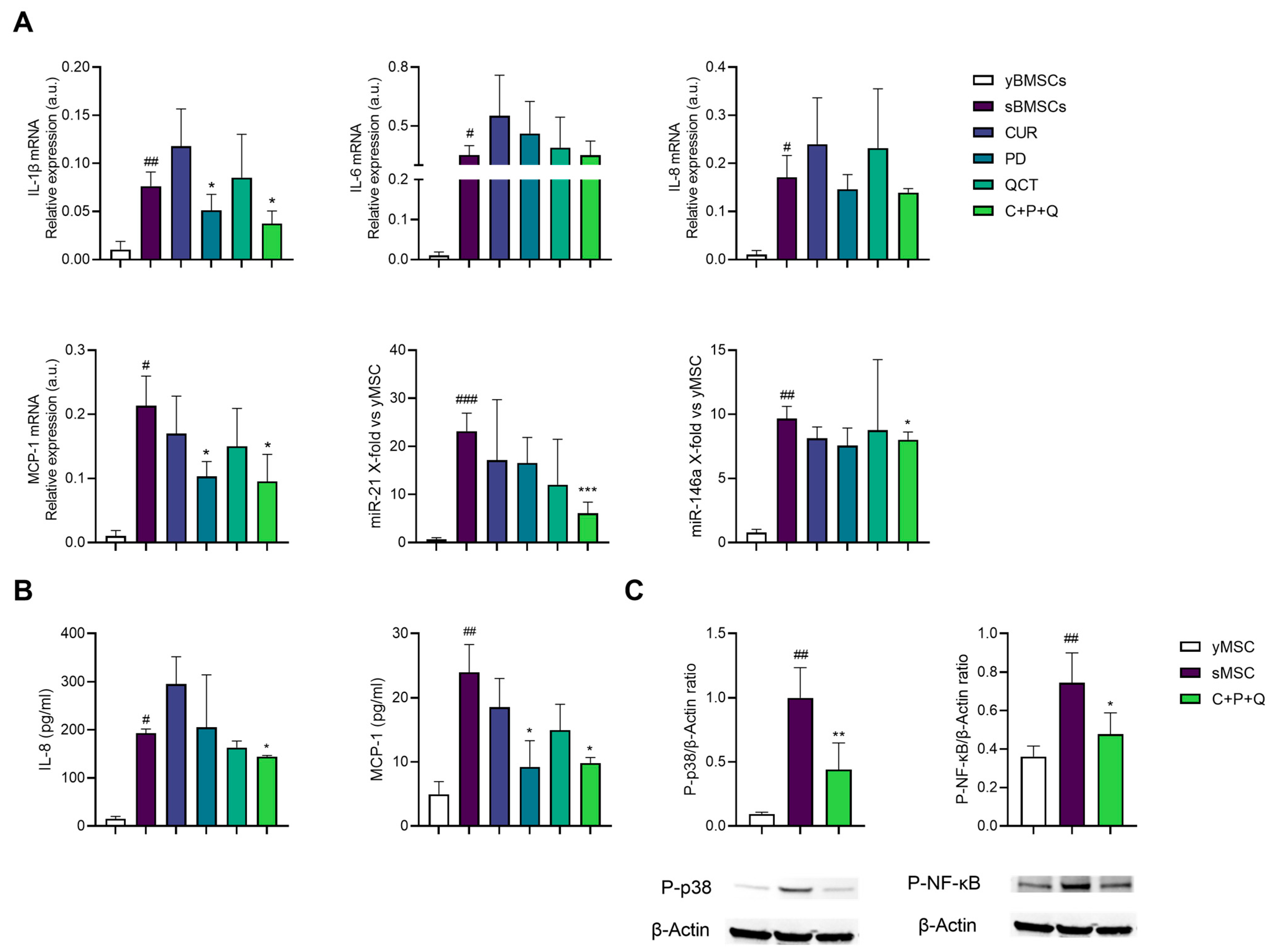
2.4. Curcumin, Polydatin and Quercetin Have Anti-Inflammatory Activity on sMSC
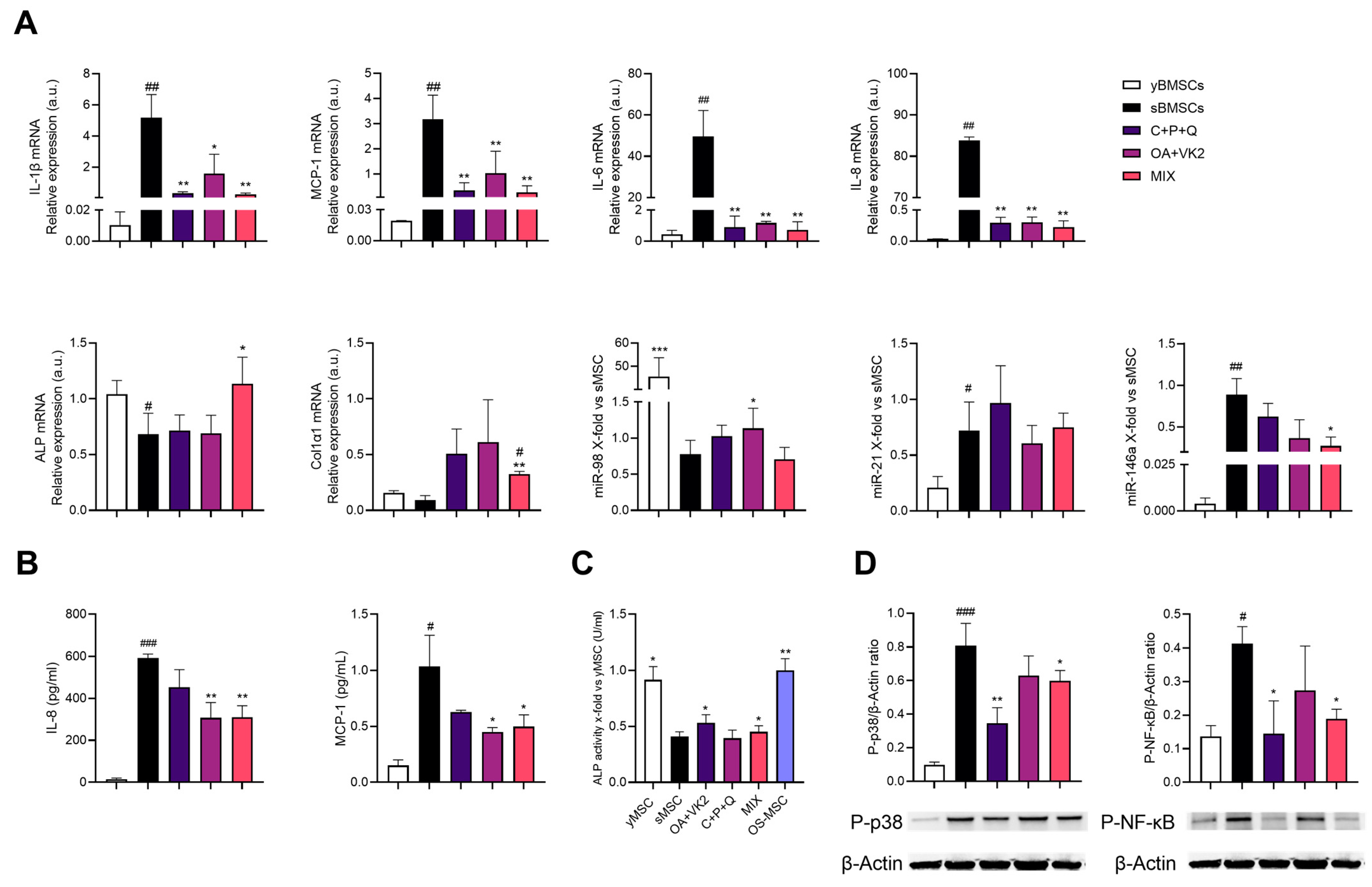
2.5. Natural Compound Combination (MIX) Exerts an Anti-Inflammatory and Osteogenic Effect on sMSC
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. MSC In Vitro Differentiation
4.3. Replicative Senes Cence of Bone Marrow Mesenchymal Stem Cells
4.4. Natural Compounds
4.5. Cell Viability Assay
4.6. Cells Treatments
4.7. RNA Isolation, mRNA Expression and miRNA Analysis by RT-qPCR
4.8. DNA Extraction and Telomere Length Measurement
4.9. Western Blot Analysis
4.10. ELISA Assay
4.11. Alkaline Phosphatase (ALP) Assay
4.12. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lane, N.E. Epidemiology, Etiology, and Diagnosis of Osteoporosis. Am. J. Obstet. Gynecol. 2006, 194, S3–S11. [Google Scholar] [CrossRef]
- Zhu, K.; Prince, R.L. Lifestyle and Osteoporosis. Curr. Osteoporos. Rep. 2015, 13, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Yu, Q. Primary Osteoporosis in Postmenopausal Women. Chronic Dis. Transl. Med. 2015, 1, 9–13. [Google Scholar] [CrossRef]
- David, K.; Narinx, N.; Antonio, L.; Evenepoel, P.; Claessens, F.; Decallonne, B.; Vanderschueren, D. Bone Health in Ageing Men. Rev. Endocr. Metab. Disord. 2022, 23, 1173–1208. [Google Scholar] [CrossRef]
- Raggatt, L.J.; Partridge, N.C. Cellular and Molecular Mechanisms of Bone Remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of Osteoblast Differentiation by Transcription Factors. J. Cell. Biochem. 2006, 99, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Infante, A.; Rodríguez, C.I. Osteogenesis and Aging: Lessons from Mesenchymal Stem Cells. Stem Cell Res. Ther. 2018, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Pignolo, R.J.; Law, S.F.; Chandra, A. Bone Aging, Cellular Senescence, and Osteoporosis. JBMR Plus 2021, 5, e10488. [Google Scholar] [CrossRef]
- Xu Feng, J.M.M. Disorders of Bone Remodelling. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 121–145. [Google Scholar] [CrossRef]
- Bergman, R.J.; Gazit, D.; Kahn, A.J.; Gruber, H.; Mcdougall, S.; Hahn, T.J. Age-Related Changes in Osteogenic Stem Cells in Mice. J. Bone Miner. Res. 1996, 11, 568–577. [Google Scholar] [CrossRef]
- Pietschmann, P.; Rauner, M.; Sipos, W.; Kerschan-Schindl, K. Osteoporosis: An Age-Related and Gender-Specific Disease—A Mini-Review. Gerontology 2009, 55, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Föger-Samwald, U.; Kerschan-Schindl, K.; Butylina, M.; Pietschmann, P. Age Related Osteoporosis: Targeting Cellular Senescence. Int. J. Mol. Sci. 2022, 23, 2701. [Google Scholar] [CrossRef] [PubMed]
- Farr, J.N.; Rowsey, J.L.; Eckhardt, B.A.; Thicke, B.S.; Fraser, D.G.; Tchkonia, T.; Kirkland, J.L.; Monroe, D.G.; Khosla, S. Independent Roles of Estrogen Deficiency and Cellular Senescence in the Pathogenesis of Osteoporosis: Evidence in Young Adult Mice and Older Humans. J. Bone Miner. Res. 2019, 34, 1407–1418. [Google Scholar] [CrossRef]
- Rizzoli, R. Postmenopausal Osteoporosis: Assessment and Management. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 739–757. [Google Scholar] [CrossRef] [PubMed]
- Lagoumtzi, S.M.; Chondrogianni, N. Senolytics and Senomorphics: Natural and Synthetic Therapeutics in the Treatment of Aging and Chronic Diseases. Free Radic. Biol. Med. 2021, 171, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Reffitt, D.M.; Ogston, N.; Jugdaohsingh, R.; Cheung, H.F.J.; Evans, B.A.J.; Thompson, R.P.H.; Powell, J.J.; Hampson, G.N. Orthosilicic Acid Stimulates Collagen Type 1 Synthesis and Osteoblastic Differentiation in Human Osteoblast-like Cells In Vitro. Bone 2003, 32, 127–135. [Google Scholar] [CrossRef]
- Jugdaohsingh, R.; Tucker, K.L.; Qiao, N.; Cupples, L.A.; Kiel, D.P.; Powell, J.J. Dietary Silicon Intake Is Positively Associated with Bone Mineral Density in Men and Premenopausal Women of the Framingham Offspring Cohort. J. Bone Miner. Res. 2004, 19, 297–307. [Google Scholar] [CrossRef]
- Anderson, J.D. Not Significantly Different, Either in Size Silicon: A Possible Factor in Bone Calcification. October 1970, 1363, 4–5. [Google Scholar]
- Zhou, H.; Jiao, G.; Dong, M.; Chi, H.; Wang, H.; Wu, W.; Liu, H.; Ren, S.; Kong, M.; Li, C.; et al. Orthosilicic Acid Accelerates Bone Formation in Human Osteoblast-Like Cells through the PI3K–Akt–MTOR Pathway. Biol. Trace Elem. Res. 2019, 190, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Akbari, S.; Rasouli-Ghahroudi, A.A. Vitamin K and Bone Metabolism: A Review of the Latest Evidence in Preclinical Studies. Biomed Res. Int. 2018, 2018, 4629383. [Google Scholar] [CrossRef]
- Abu-Amer, Y. NF-ΚB Signaling and Bone Resorption. Osteoporos. Int. 2013, 24, 2377–2386. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Weitzmann, M.N. Vitamin K2 Stimulates Osteoblastogenesis and Suppresses Osteoclastogenesis by Suppressing NF-ΚB Activation. Int. J. Mol. Med. 2011, 27, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Gurău, F.; Baldoni, S.; Prattichizzo, F.; Espinosa, E.; Amenta, F.; Procopio, A.D.; Albertini, M.C.; Bonafè, M.; Olivieri, F. Anti-Senescence Compounds: A Potential Nutraceutical Approach to Healthy Aging. Ageing Res. Rev. 2018, 46, 14–31. [Google Scholar] [CrossRef]
- Weiskirchen, S.; Weiskirchen, R. Resveratrol: How Much Wine Do You Have to Drink to Stay Healthy? Adv. Nutr. 2016, 7, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Bielak-Zmijewska, A.; Grabowska, W.; Ciolko, A.; Bojko, A.; Mosieniak, G.; Bijoch, Ł.; Sikora, E. The Role of Curcumin in the Modulation of Ageing. Int. J. Mol. Sci. 2019, 20, 1239. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.C.; Takada, Y.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits receptor activator of NF-kappa B ligand-induced NF-kappa B activation in osteoclast precursors and suppresses osteoclastogenesis. J. Immunol. 2004, 172, 5940–5947. [Google Scholar] [CrossRef] [PubMed]
- Hatefi, M.; Ahmadi, M.R.H.; Rahmani, A.; Dastjerdi, M.M.; Asadollahi, K. Effects of Curcumin on Bone Loss and Biochemical Markers of Bone Turnover in Patients with Spinal Cord Injury. World Neurosurg. 2018, 114, e785–e791. [Google Scholar] [CrossRef]
- Zhan, J.; Luo, D.; Zhao, B.; Chen, S.; Luan, J.; Luo, J.; Hou, Y.; Hou, Y.; Xu, W.; Yan, W.; et al. Polydatin Administration Attenuates the Severe Sublesional Bone Loss in Mice with Chronic Spinal Cord Injury. Aging 2022, 14, 8856–8875. [Google Scholar] [CrossRef] [PubMed]
- Eren, T.; Baysal, G.; Doğan, F. Biocidal Activity of Bone Cements Containing Curcumin and Pegylated Quaternary Polyethylenimine. J. Polym. Environ. 2020, 28, 2469–2480. [Google Scholar] [CrossRef]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [CrossRef]
- Braun, K.F.; Ehnert, S.; Freude, T.; Egaña, J.T.; Schenck, T.L.; Buchholz, A.; Schmitt, A.; Siebenlist, S.; Schyschka, L.; Neumaier, M.; et al. Quercetin Protects Primary Human Osteoblasts Exposed to Cigarette Smoke through Activation of the Antioxidative Enzymes HO-1 and SOD-1. Sci. World J. 2011, 11, 2348–2357. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Y.; Wang, Z.H.; Deng, L.H.; Wang, H.; Zheng, Q. Oral Administration of Quercetin or Its Derivatives Inhibit Bone Loss in Animal Model of Osteoporosis. Oxid. Med. Cell. Longev. 2020, 2020, 6080597. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.Y.; Ima-Nirwana, S. Quercetin as an Agent for Protecting the Bone: A Review of the Current Evidence. Int. J. Mol. Sci. 2020, 21, 6448. [Google Scholar] [CrossRef] [PubMed]
- Teissier, T.; Temkin, V.; Pollak, R.D.; Cox, L.S. Crosstalk between Senescent Bone Cells and the Bone Tissue Microenvironment Influences Bone Fragility during Chronological Age and in Diabetes. Front. Physiol. 2022, 13, 1019741. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Weitzmann, M.N. Quercetin, a Potent Suppressor of NF-ΚB and Smad Activation in Osteoblasts. Int. J. Mol. Med. 2011, 28, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Chen, G.; Feng, S.; Wang, P.; Zhu, X.; Zhang, R. Quercetin Promotes the Osteogenic Differentiationof Rat Mesenchymal Stem Cells via Mitogen-Activated Protein Kinase Signaling. Exp. Ther. Med. 2015, 9, 2072–2080. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.G.; Cong, Y.; Bao, N.R.; Li, Y.G.; Zhao, J.N. Quercetin Stimulates Bone Marrow Mesenchymal Stem Cell Differentiation through an Estrogen Receptor-Mediated Pathway. Biomed Res. Int. 2018, 2018, 1799. [Google Scholar] [CrossRef]
- Raisz, L.G. Pathogenesis of Osteoporosis: Concepts, Conflicts, and Prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef]
- Gao, X.L.; Cao, M.G.; Ai, G.G.; Hu, Y.B. Mir-98 Reduces the Expression of HMGA2 and Promotes Osteogenic Differentiation of Mesenchymal Stem Cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3311–3317. [Google Scholar] [CrossRef]
- Liu, Q.; Guo, Y.; Wang, Y.; Zou, X.; Yan, Z. MiR-98-5p Promotes Osteoblast Differentiation in MC3T3-E1 Cells by Targeting CKIP-1. Mol. Med. Rep. 2018, 17, 4797–4802. [Google Scholar] [CrossRef]
- Yu, B.; Wang, C.Y. Osteoporosis: The Result of an ‘Aged’ Bone Microenvironment. Trends Mol. Med. 2016, 22, 641–644. [Google Scholar] [CrossRef] [PubMed]
- Matacchione, G.; Valli, D.; Silvestrini, A.; Giuliani, A.; Sabbatinelli, J.; Giordani, C.; Coppari, S.; Rippo, M.R.; Albertini, M.C.; Olivieri, F. Curcumin, Polydatin and Quercetin Synergistic Activity Protects from High-Glucose-Induced Inflammation and Oxidative Stress. Antioxidants 2022, 11, 1037. [Google Scholar] [CrossRef]
- Olivieri, F.; Prattichizzo, F.; Giuliani, A.; Matacchione, G.; Rippo, M.R.; Sabbatinelli, J.; Bonafè, M. MiR-21 and MiR-146a: The MicroRNAs of Inflammaging and Age-Related Diseases. Ageing Res. Rev. 2021, 70, 101374. [Google Scholar] [CrossRef] [PubMed]
- Journal, T.; North, T.; Menopause, A.; Vol, S.; North, T.; Menopause, A. Management of Osteoporosis in Postmenopausal Women: The 2021 Position Statement of The North American Menopause Society. Menopause 2021, 28, 973–997. [Google Scholar] [CrossRef]
- Ebina, K.; Noguchi, T.; Hirao, M.; Kaneshiro, S.; Tsukamoto, Y.; Yoshikawa, H. Comparison of the Effects of 12 Months of Monthly Minodronate Monotherapy and Monthly Minodronate Combination Therapy with Vitamin K2 or Eldecalcitol in Patients with Primary Osteoporosis. J. Bone Miner. Metab. 2016, 34, 243–250. [Google Scholar] [CrossRef]
- Zhao, B.; Zhao, W.; Wang, Y.; Zhao, Z.; Zhao, C.; Wang, S.; Gao, C. Prior Administration of Vitamin K2 Improves the Therapeutic Effects of Zoledronic Acid in Ovariectomized Rats by Antagonizing Zoledronic Acid-Induced Inhibition of Osteoblasts Proliferation and Mineralization. PLoS ONE 2018, 13, e0202269. [Google Scholar] [CrossRef]
- Chou, L.Y.; Ho, C.T.; Hung, S.C. Paracrine Senescence of Mesenchymal Stromal Cells Involves Inflammatory Cytokines and the NF-ΚB Pathway. Cells 2022, 11, 3324. [Google Scholar] [CrossRef]
- Vimalraj, S. Alkaline Phosphatase: Structure, Expression and Its Function in Bone Mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef] [PubMed]
- Kushida, K.; Takahashi, M.; Kawana, K.; Inoue, T. Comparison of Markers for Bone Formation and Resorption in Premenopausal and Postmenopausal Subjects, and Osteoporosis Patients. J. Clin. Endocrinol. Metab. 1995, 80, 2447–2450. [Google Scholar] [CrossRef]
- Marshall, C.D.; Brett, E.A.; Moore, A.L.; Wan, D.C.; Longaker, M.T. In Vitro and In Vivo Osteogenic Differentiation of Human Adipose-Derived Stromal Cells. Methods Mol. Biol. 2019, 1891, 9–18. [Google Scholar]
- Cawthon, R.M. Telomere Measurement by Quantitative PCR. Nucleic Acids Res. 2002, 30, 1–6. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| yMSC—7 Days | RUNX2 | ALP | Col1α1 | OCN | ||||
| Effect vs. yMSC (%) | Synergistic Effect * | Effect vs. yMSC (%) | Synergistic Effect * | Effect vs. yMSC (%) | Synergistic Effect * | Effect vs. yMSC (%) | Synergistic Effect * | |
| Orthosilicic acid | 18 | Y | 6 | Y | 48 | N | 64 | N |
| Vitamin K2 | 0 | 1 | 53 | 154 | ||||
| OA+VK2 | 68 | 15 | 91 | 133 | ||||
| yMSC—14 days | RUNX2 | ALP | Col1α1 | OCN | ||||
| Effect vs. yMSC (%) | Synergistic Effect * | Effect vs. yMSC (%) | Synergistic Effect * | Effect vs. yMSC (%) | Synergistic Effect * | Effect vs. yMSC (%) | Synergistic Effect * | |
| Orthosilicic acid | 43 | N | 1 | N | 14 | Y | 49 | Y |
| Vitamin K2 | 66 | 63 | 79 | 63 | ||||
| OA+VK2 | 7 | 16 | 214 | 232 | ||||
| sMSC 7 Days | IL-1β | MCP-1 | IL-6 | IL-8 | ALP | COL1α1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Effect vs. yMSC (%) | Synergistic effect * | Effect vs. yMSC (%) | Synergistic effect * | Effect vs. yMSC (%) | Synergistic effect * | Effect vs. yMSC (%) | Synergistic effect * | Effect vs. yMSC (%) | Synergistic effect * | Effect vs. yMSC (%) | Synergistic effect * | |
| OA+VK2 | 70 | N | 67 | N | 97 | N | 99 | N | 11 | 623 | ||
| C+P+Q | 94 | 88 | 98 | 99 | 10 | Y | 497 | N | ||||
| MIX | 96 | 91 | 98 | 99 | 87 | 310 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordani, C.; Matacchione, G.; Giuliani, A.; Valli, D.; Scarpa, E.S.; Antonelli, A.; Sabbatinelli, J.; Giacchetti, G.; Sabatelli, S.; Olivieri, F.; et al. Pro-Osteogenic and Anti-Inflammatory Synergistic Effect of Orthosilicic Acid, Vitamin K2, Curcumin, Polydatin and Quercetin Combination in Young and Senescent Bone Marrow-Derived Mesenchymal Stromal Cells. Int. J. Mol. Sci. 2023, 24, 8820. https://doi.org/10.3390/ijms24108820
Giordani C, Matacchione G, Giuliani A, Valli D, Scarpa ES, Antonelli A, Sabbatinelli J, Giacchetti G, Sabatelli S, Olivieri F, et al. Pro-Osteogenic and Anti-Inflammatory Synergistic Effect of Orthosilicic Acid, Vitamin K2, Curcumin, Polydatin and Quercetin Combination in Young and Senescent Bone Marrow-Derived Mesenchymal Stromal Cells. International Journal of Molecular Sciences. 2023; 24(10):8820. https://doi.org/10.3390/ijms24108820
Chicago/Turabian StyleGiordani, Chiara, Giulia Matacchione, Angelica Giuliani, Debora Valli, Emanuele Salvatore Scarpa, Antonella Antonelli, Jacopo Sabbatinelli, Gilberta Giacchetti, Sofia Sabatelli, Fabiola Olivieri, and et al. 2023. "Pro-Osteogenic and Anti-Inflammatory Synergistic Effect of Orthosilicic Acid, Vitamin K2, Curcumin, Polydatin and Quercetin Combination in Young and Senescent Bone Marrow-Derived Mesenchymal Stromal Cells" International Journal of Molecular Sciences 24, no. 10: 8820. https://doi.org/10.3390/ijms24108820