
Exploring Redox Modulation of Plant UDP-Glucose Pyrophosphorylase



1
Department of Plant Physiology, Umeå Plant Science Centre, Umeå University, 90187 Umeå, Sweden
2
Diamyd Medical, 90621 Umeå, Sweden
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(10), 8914; https://doi.org/10.3390/ijms24108914
Submission received: 1 April 2023
/
Revised: 10 May 2023
/
Accepted: 15 May 2023
/
Published: 17 May 2023
(This article belongs to the Collection Feature Papers in Molecular Plant Sciences)
Abstract
:UDP-glucose (UDPG) pyrophosphorylase (UGPase) catalyzes a reversible reaction, producing UDPG, which serves as an essential precursor for hundreds of glycosyltransferases in all organisms. In this study, activities of purified UGPases from sugarcane and barley were found to be reversibly redox modulated in vitro through oxidation by hydrogen peroxide or oxidized glutathione (GSSG) and through reduction by dithiothreitol or glutathione. Generally, while oxidative treatment decreased UGPase activity, a subsequent reduction restored the activity. The oxidized enzyme had increased Km values with substrates, especially pyrophosphate. The increased Km values were also observed, regardless of redox status, for UGPase cysteine mutants (Cys102Ser and Cys99Ser for sugarcane and barley UGPases, respectively). However, activities and substrate affinities (Kms) of sugarcane Cys102Ser mutant, but not barley Cys99Ser, were still prone to redox modulation. The data suggest that plant UGPase is subject to redox control primarily via changes in the redox status of a single cysteine. Other cysteines may also, to some extent, contribute to UGPase redox status, as seen for sugarcane enzymes. The results are discussed with respect to earlier reported details of redox modulation of eukaryotic UGPases and regarding the structure/function properties of these proteins.
1. Introduction
UDP-glucose (UDPG) pyrophosphorylase, a cytosolic protein, is a key enzyme of carbohydrate biosynthesis, producing UDPG, which serves as a substrate (or precursor) for the biosynthesis of a plethora of oligo- and polysaccharides, including sucrose (the main carbon transporting molecule in plants) as well as cellulose, hemicellulose, and other cell wall components [1,2,3,4,5]. Although UDPG production is thought to represent the major function of UGPase, its reaction is fully reversible. From thermodynamic principles, because of its reversibility and high activity, UGPase is considered an “equilibrating” enzyme coupled through UDPG production to several slow irreversible reactions (“engine” enzymes) [6,7,8]. UGPase uses UTP and glucose-1-P as substrates in the so-called forward direction of the reaction, as well as UDPG and pyrophosphate (PPi) as substrates in the reverse direction. As a consequence, UGPase sits at the crucial junction of plant metabolism, being indirectly involved in the synthesis of complex carbohydrates as well as in sugar and energy metabolism [8,9,10]. A central role in integrating primary metabolism was also proposed for UGPases from other eukaryotes [11,12,13].
Plant UGPase has been well characterized with respect to its physical/kinetic properties and regulation [5,14,15,16,17]. Surprisingly, however, no focused studies were carried out on possible redox modulation of the higher plant enzyme, a process known to affect hundreds of proteins [18,19]. Earlier studies on simple single-celled eukaryotes have revealed that UGPase activity is modulated by a redox mechanism involving oxidation and reduction of specific cysteine (Cys) residues [12,20,21]. For plant UGPases, the first hints of its possible redox control came from high-throughput proteomics studies [22,23], where seed UGPase was identified as one of many targets for in vivo interaction with thioredoxins, small proteins mediating redox control during oxidative stress conditions [24]. UGPase was also one of many protein targets that were S-glutathionylated at Cys residues in Arabidopsis cell culture during oxidative stress [25]. Glutathione (GSH), a peptide containing three amino acids (Glu-Cys-Gly), serves important functions in plants as a reductant and, along with its oxidized form (GSSG), is well known to participate in plant oxidative stress responses by reversibly modifying selected proteinaceous Cys residues [26,27,28,29]. More recently, a report by Soares et al. [30] demonstrated that the activity of purified sugarcane UGPase decreased upon oxidation by hydrogen peroxide (H2O2), whereas subsequent reduction by dithiothreitol (DTT) restored the activity. Since H2O2 is well known as a cellular signal of oxidative stress [31], whereas DTT can mimic the effects of thioredoxins and GSH [26,32,33], this suggested that redox modulation may indeed represent yet another level of regulation of plant UGPases in vivo.
Crystal structures of Arabidopsis and sugarcane UGPases have revealed that the proteins contain a Cys residue (Cys95 and Cys102, respectively) at the so-called nucleotide-binding (NB) loop [34,35]. Earlier kinetic studies on a barley UGPase mutant lacking Cys99 (homologous to Arabidopsis Cys95 and sugarcane Cys102) demonstrated that it had a decreased affinity for its substrates, especially for PPi [36].
To examine in more detail the redox control of plant UGPase activity, in the present study, we used purified wild-type (wt) UGPases from both sugarcane and barley. In addition, their respective mutants lacking a single Cys at the NB loop were used to check the role of this Cys in redox responses. Both sugarcane and barley wt UGPases were sensitive to redox modulation, which affected both the activity and substrate affinity of the enzymes, with a major, but not exclusive, involvement of the Cys99 and Cys102 of barley and sugarcane UGPases, respectively. We have also shown that glutathione in its reduced (GSH) and oxidized (GSSG) forms can readily substitute for DTT and H2O2, respectively, as effective modulators of UGPase activity. The results are discussed with respect to the structure-function properties of plant UGPase.
2. Results
2.1. Redox Modulation of Sugarcane UGPase
To study redox effects on sugarcane UGPase, we used both wt protein and its Cys102Ser mutant and treated them with either H2O2 or oxidized glutathione (GSSG). Oxidation by up to 20 mM of GSSG or H2O2 for 30 min led to a decrease in activity for both wt and Cys102Ser mutant enzymes, and the effects of GSSG were stronger than those of H2O2. The mutant appeared less sensitive to H2O2 treatment but was almost as strongly affected as wt by GSSG (Figure 1). Both H2O2 and GSSG had stronger inhibitory effects on sugarcane wt UGPase activity when compared to its Cys102Ser mutant (Figure 1).
The GSSG-treated enzyme could regain its activity when reduced by DTT or GSH, and this concerned both wt UGPase and its Cys102Ser mutant (Figure 2), indicating that the oxidation process could be reversed by the reductant. The GSSG-oxidized sugarcane proteins were more sensitive to DTT and GSH reduction than the untreated enzymes (Figure 2). Earlier, the DTT-stimulated recovery of activity was observed for H2O2-pre-treated sugarcane UGPase, but the effects of GSSG and GSH were not tested [30].
For kinetic characterization of sugarcane wt UGPase and its Cys102Ser mutant, we have determined Km values for UDPG and PPi by varying the concentration of one substrate at the constant fixed level of the other substrate. The analyses were done both for the non-treated and GSSG-treated proteins (Figure 3). For either wt and the mutant, their Km values with UDPG were roughly similar (0.08 and 0.11 mM, respectively), and both proteins displayed a decreased affinity for UDPG (increased Km) after oxidation by GSSG (Figure 3). The Km with PPi, however, was much lower for wt UGPase than for the Cys102Ser mutant (0.05 mM versus 0.23 mM). Similar results were obtained earlier for purified barley UGPase and its Cys99Ser mutant, where the Km with PPi of wt protein was 7–10-fold lower than that of the mutant [36,37].
Oxidation of sugarcane wt UGPase by GSSG led to a two–three-fold increase in its Km values with UDPG and PPi, while the Kms of Cys102Ser protein also increased but were less strongly affected by GSSG (Figure 3). Nevertheless, in the case of the mutant, the GSSG-induced increase of its Km values suggests that oxidation of Cys residues other than Cys102 may have also contributed to the lower substrate affinity of the oxidized enzyme (see Section 3).
2.2. Redox Modulation of Barley UGPase
Barley UGPase has been the primary model to study plant UGPases, both at the protein structure and enzyme regulation levels [5,36,37,38]. Thus, given the evidence on redox control of sugarcane UGPase (Figure 1, Figure 2 and Figure 3), it was of interest whether barley UGPase is also responsive to redox control.
Compared to sugarcane wt UGPase, the barley wt enzyme was more sensitive to H2O2 treatment. Its activity was at least two-fold inhibited by oxidation by as little as 0.5 mM H2O2 (Figure 4A). This needs to be compared with the activity of sugarcane wt UGPase, which was inhibited by less than 50% only by 20 mM H2O2 (Figure 1). On the other hand, the Cys99Ser mutant was resistant to H2O2 treatment, showing only about 15% inhibition at 5 mM H2O2 (Figure 4B). The H2O2-oxidized wt and mutant proteins could regain all their activity after treatment with DTT (Figure 4B). The restoration of activity by DTT has indicated the reversibility of redox modulation of barley UGPase, similar to the sugarcane enzyme that was oxidized by GSSG and reduced by DTT or GSH (Figure 2).
The Km values for barley UGPase were determined for purified wt and Cys99Ser proteins which were pre-incubated with either H2O2 or DTT (Figure 5). These conditions differed from those for Km determinations of sugarcane UGPase, where purified wt and Cys102Ser proteins were pre-incubated with or without GSSG (Figure 3). However, in both cases, oxidation by either H2O2 (barley enzyme) or GSSG (sugarcane enzyme) resulted in similar effects on Km values. Thus, the oxidation led to a ca. two-fold increase in the Km with UDPG for both wt and mutant enzymes and a several-fold increase in the Km with PPi for wt UGPases, but not for the mutants (Figure 3 and Figure 5). Treatment with H2O2 or DTT had no effect on the Km values of the barley Cys99Ser mutant, and those values corresponded to the Kms of oxidized wt protein (Figure 5). This suggests that Cys99, and no other Cys residues, is essential for redox modulation of barley UGPase. Supporting this is little or no effect of 5 mM H2O2 on the activity of the mutant (Figure 4B).
3. Discussion
3.1. Redox Compounds Affecting Plant UGPase Activity
Generally, we found that plant UGPase is sensitive to oxidation by H2O2 and GSSG (e.g., Figure 1). These two compounds are believed to be the main players during oxidative stress conditions, and they regulate the activities of hundreds of proteins via post-translational modification of selected Cys residues [18]. The oxidation resulted in lower activities for both barley and sugarcane UGPases (Figure 1, Figure 2 and Figure 4). This decrease in activity could be reverted by the treatment of oxidized UGPase with DTT or GSH (Figure 2 and Figure 4).
Both H2O2 and GSSG cause oxidation of proteinaceous Cys residues, but their modes of action are different. Whereas H2O2 oxidizes the -SH group of a Cys to sulfenic acid (-SOH) [38], the effect of GSSG is to oxidize (glutathionylate) the -SH group to a bulky -SSG [27]. In most organisms and in cell culture, the concentration of H2O2 was estimated at extremely low levels, from pico- to nanomolar [39,40]. At low levels, it acts as a signaling molecule, and at high levels, it induces cell death [41]. There are exceptions, however, e.g., in sugarcane leaves but not sugarcane stem internodes, H2O2 was reported at up to 50 mM [30]. Glutathione, on the other hand, is present at millimolar concentrations in all aerobic organisms, and the GSSG/GSH ratio is an important indicator of oxidative stress conditions [42].
UGPase is one of many proteins that undergo S-glutathionylation, as found for Arabidopsis [25]. The GSSG (along with its reduced form, GSH) is believed to be involved in redox regulation in vivo of a variety of proteins, especially those located in the cytosol, in both plant and animal cells [43,44]. In most cases, the process involves the use of GSSG for a reversible modification of protein cysteinyl residues that can directly modulate a given protein activity [29]. In barley, using a genetically-encoded biosensor of cytosolic glutathione redox potential, this potential was found to be highly robust under combined salt and osmotic stresses [45]. This is consistent with plant UGPase activity being responsive to these stresses [46,47,48].
3.2. Redox and Substrate Affinity of Plant UGPases
In our earlier work on barley UGPase mutants, the Cys99Ser mutant had characteristically increased Km values (i.e., lower substrate affinity) with UDPG and, especially, with PPi, when compared to Kms of the wt enzyme [36,37]. Similar results were obtained in the present study for both barley Cys99Ser and sugarcane Cys102Ser mutants. As seen in Table 1, which lists Km values for redox-modulated sugarcane and barley UGPases (based on Figure 3 and Figure 5), the untreated or reduced mutants had increased Km values for both UDPG and PPi, when compared to their respective wt proteins. This was especially evident for Km values with PPi, which were ca. six- and 10-fold higher for Cys102Ser and Cys99Ser mutants when compared to those of their wt counterparts. Upon oxidation of wt enzymes, their Kms with UDPG and PPi notably increased, especially in the case of Km with PPi for barley wt UGPase (Table 1). On the other hand, oxidation of the mutants led only to a moderate increase in Km values for the sugarcane Cys102Ser protein but had no effect on the substrate affinity of the Cys99Ser barley mutant (Table 1).
For both sugarcane and barley wt UGPases, the differences in Km values between non-treated/reduced and oxidized enzymes suggest that redox conditions affect their substrate affinity. For the sugarcane enzyme, the redox status of its Cys102 appears to be the major, but not the only, factor involved in these effects. In contrast, for barley UGPase, its Cys99 appears to be responsible for most, if not all, of redox effects.
Redox-induced changes in substrate affinity (Table 1) may have important consequences for in vivo activities of plant UGPase, especially with respect to its affinity for PPi. Furthermore, PPi serves as a UGPase substrate only when complexed with Mg2+ (as MgPPi), whereas free PPi is a strong inhibitor of the enzyme [37,49]. Thus, under oxidative stress, the lower affinity of UGPase for PPi may lead to an increase in the cytosolic concentration of free PPi, resulting in a further decrease in the activity of this enzyme.
3.3. Structural Basis for Redox Control of Plant UGPases
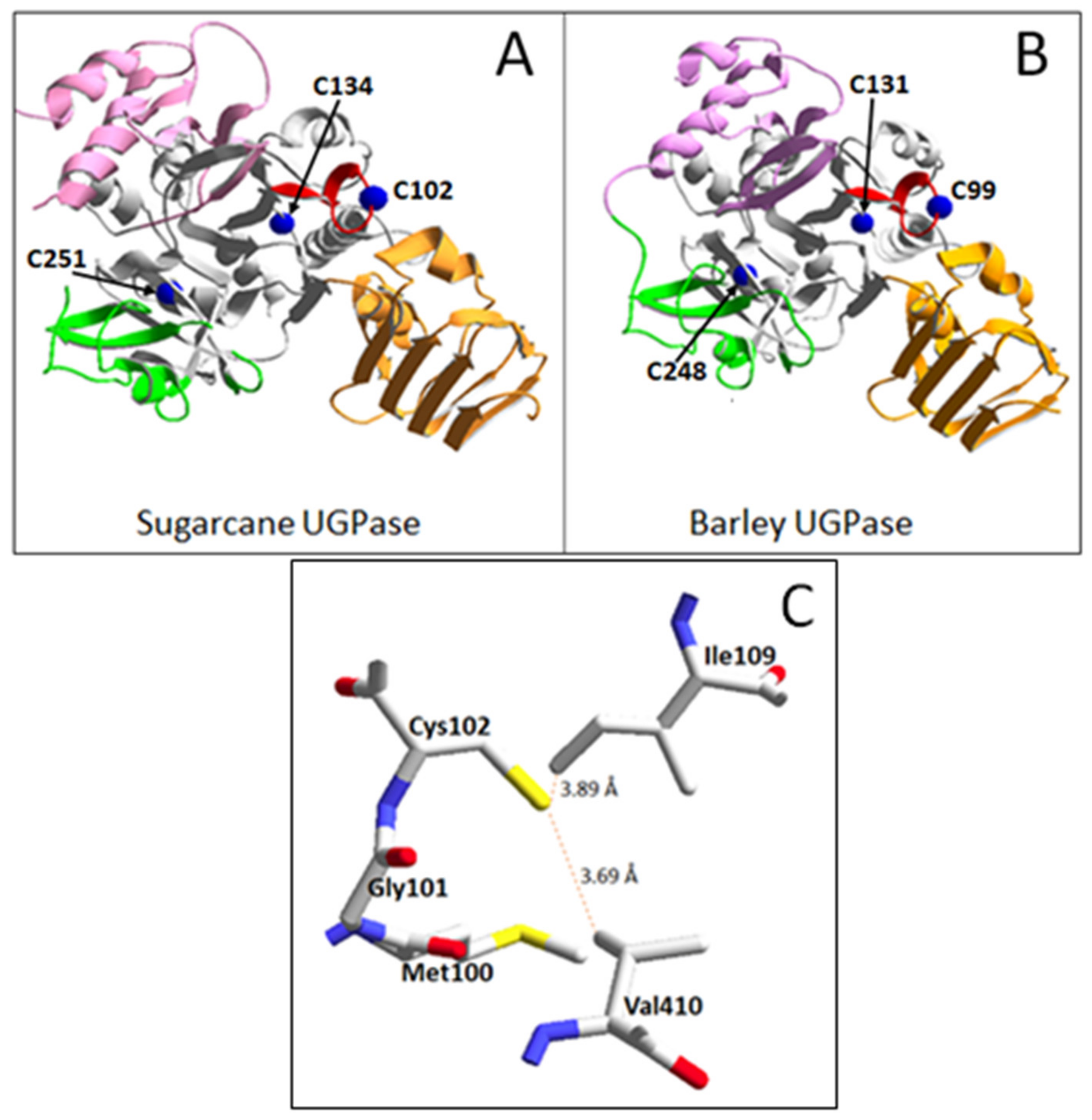
Amino acid sequences of both sugarcane and barley UGPases contain only three Cys residues: Cys102, Cys134, and Cys251 for sugarcane UGPase; and Cys99, Cys131, and Cys248 for the barley protein. Sugarcane UGPase Cys102 and its barley homolog Cys99 are located at the NB loop that is close to the active site of the protein [34,35,37], whereas the other two Cys are more distant and are located in the central domain (Figure 6A,B). For the three Cys residues, only Cys102 (sugarcane protein) and Cys99 (barley protein) are conserved in all eukaryotic UGPases, whereas other Cys are conserved for UGPases only in plants [12,20,30]. If the three Cys molecules are analyzed in pairs, their relative distances range between 23 and 35 Å from each other; this is too far for any pair of these Cys to form a disulfide bridge (S-S). In comparison, a typical distance between Cys residues in a protein structure to form an S-S bond is 2.03 Å [12]. Thus, it seems unlikely that the redox effect on plant UGPase involves disulfide bond formation for its internal Cys residues. More likely, the UGPase Cys residues are reversibly S-glutathionylated and S-sulfenylated by GSSG and H2O2, respectively. The -SH group of Cys that was oxidized by either H2O2 or GSSG can be reduced back by either GSH, DTT, or reduced thioredoxin [50]. Seed UGPases from wheat and Medicago truncatula have already been identified as in vivo targets for thioredoxin reduction [22,23].
Among eukaryotic UGPases, only the enzyme from Entamoeba histolytica, a single-celled parasite, was demonstrated to be redox modulated via reversible disulfide bond formation for two of its Cys residues [12], and this was shown by detailed modeling and SDS-PAGE analyses of oxidized and reduced UGPase protein. The Cys molecules involved were Cys108 and Cys378 [12], the former corresponding to Cys99 and Cys102 of barley and sugarcane UGPases, respectively [30,36].
For sugarcane UGPase, two of the near neighbors of Cys102 are Ile109 (also located on the NB loop) and Val410 (from the C-terminal domain) (Figure 6C). Corresponding configurations are also present in Arabidopsis and barley UGPases. As hydrophobic interactions usually have carbon-carbon distances of 3.3–4.0 Å, the short distances shown in Figure 6C suggest that Cys 102 is involved in hydrophobic interplay with these amino acids. Cysteine, despite possessing a polar sulfhydryl group, frequently tends to behave as a hydrophobic (rather than polar) residue in folded protein structures [51].
It is unknown how the oxidation of a specific Cys residue (Cys102 and Cys99 for sugarcane and barley UGPases, respectively) inhibits UGPase activity and leads to an increase of Km values for its substrates. This Cys, located near the tip of the NB loop (Figure 6), is distant from the active site of each enzyme. However, the loop contains three other amino acids known to coordinate substrate binding. In Arabidopsis UGPase, these amino acids are Leu85—coordinating ribose moiety of UTP; Gly87—uridyl group; and Lys99—phosphate moiety (all numbered as in Arabidopsis UGPase) [34]. These amino acids have their homologs in sugarcane and barley UGPases. Thus, it is tempting to suggest that oxidation of the NB-loop-located Cys may affect the conformation of the NB loop itself, which in turn could affect the binding of substrates in the active site. A similar explanation may also be given for low substrate affinity (higher Km values) of the Cys99Ser and Cys102Ser mutants used in this study. Taking into account the hydrophobic environment for the NB-located Cys (Figure 6C), replacing it with Ser (a hydrophilic amino acid) could not only affect the conformation of the NB loop but also disrupt inter-domain hydrophobic interactions (through Val410). More detailed studies are required to address this.
It also remains to be investigated whether redox modulation of UGPase affects its tertiary structure. Plant UGPases are active as monomers only, but under some conditions, they can form inactive dimers and higher-order oligomers [30,34,35,36,37,52,53,54]. In comparison, human UGPase is active only as an octamer, and octamer formation is essential for its activity [13,55], even though the human and plant enzymes share over 50% identity at their amino acid sequences [38]. In sugarcane, UGPase was present as both a monomer and a dimer in the leaves but only as a monomer in the stem internodes [30]. In the same study, sugarcane leaves, but not internodes, were reported to accumulate large amounts (up to 50 mM) of H2O2. This inferred that H2O2 might affect the quaternary structure of UGPase, and, thus, its activity [30]. Earlier investigations of UGPase from Euglena gracilis, a simple single-cell photosynthetic organism, have suggested that, after oxidation by H2O2, the enzyme arranges in several enzymatically-inactive structural conformations, both in vitro and in vivo [21]. The reported presence of UGPase monomers and dimers in sugarcane leaves containing high [H2O2] [30] suggests that oligomerization is a regulatory mechanism controlling UGPase activity in response to oxidative stress; however, its exact mechanism is still unknown.
3.4. Plant UGPase Is under Transcriptional, Posttranslational, and Metabolite Control
Over the years, it has become clear that UGPase is regulated at several levels, including transcriptional (at gene expression level), post-translational (at protein level), and metabolic regulation (at activity level) [5]. Each of these modes of regulation on their own may considerably affect UGPase activity and, subsequently, the flow of carbon in overall carbohydrate metabolism in both photosynthetic and non-photosynthetic tissues. Those modes of control are frequently overlapping, thus making UGPase activity even more sensitive to their regulation.
Concerning transcriptional control, UGPase gene expression was already shown as strongly affected by light-dark transition, low temperature, phosphate deficiency, salt stress, heavy metal stress, and exposure to sucrose and other sugars [30,46,53,56,57,58,59,60]. In many cases, the amount of UGPase protein and its activity were roughly correlated with changes in UGPase gene expression. As common in such studies, it is difficult to differentiate whether changes in UGPase activity during such stresses are brought about by direct effects of the stressor on UGPase (e.g., affecting the stability of the protein) or indirectly via effects on UGPase gene expression or both [16]. A good example is H2O2, which besides directly affecting UGPase activity and its substrate affinity, as shown in this study (Figure 3, Figure 4 and Figure 5), is known to affect the expression of a multitude of genes [61]. At low concentrations, H2O2 acts as a messenger molecule triggering tolerance against various abiotic stresses [31], while at high concentrations, it orchestrates programmed cell death [62].
Plant UGPases have been shown to be post-translationally modified in a number of ways. Examples include phosphorylation of Ser419 of sugarcane UGPase [30], binding to 14-3-3 proteins for the enzymes from barley and Arabidopsis [63,64], rice UGPase acetylation [65], and N-glycosylation of rice and maize UGPases [66,67]. It appears now that also redox modulation, as shown here for sugarcane and barley UGPases, needs to be added to this ever-growing list.
Substrate availability and product inhibition may also affect UGPase activity [15,68]. UGPase, by carrying a fully reversible reaction, may be involved in anabolic pathways by using UTP and glucose-1-P to produce PPi and UDPG (the latter serving as a key precursor of oligo- and polysaccharides), but also—in its reverse reaction—in catabolic processes, where glucose-1-P is broken down during respiration [5,8,49]. Thus, the direction of the UGPase reaction depends on the overall metabolic status of a tissue and may change depending on environmental/stress factors. Strong product inhibition by PPi of the forward UGPase reaction is a well-known metabolic mechanism [15,49], modulating the direction of carbohydrate metabolism in vivo [68]. A number of chemical inhibitors have been described that strongly inhibit UGPase activity (at a µM range) both in vitro and in planta [69,70]; however, all of them also affect the activities of so-called UDP-sugar pyrophosphorylases, which are related to UGPase in that they can non-specifically produce UDPG or use it as a substrate [71].
Finally, the UGPase reaction and its reversibility depend very much on the concentration of free magnesium (Mg2+), which regulates the formation of both MgATP and MgPPi complexes [72] that are true substrates/products of UGPase [49,73]. Changes in [Mg2+] reflect the energy status of a given tissue and arise mostly from intracellular pools of adenylates, uridylates, and other nucleotides [74,75,76]. Adequate intracellular [Mg2+] has already been shown to ease oxidative stress by affecting the phloem transport of sugars and controlling optimal CO2 fixation, especially under high-light conditions [77].
More detailed studies are required on the redox status of the NB-located Cys and other Cys residues of plant UGPase, involving MALDI-TOF mass spectrometry analyses. This should also include an investigation of the redox status of methionine (Met) residues. Met can have the same function as Cys residues when it comes to redox regulation [78]. There is also a possibility that plant UGPases contain a non-catalytic binding site(s) for MgPPi, as reported, e.g., for E. coli ATPase [79]. Overall, this would give a more profound picture of redox regulation at the UGPase active site and potentially on alternative binding sites.
In conclusion, redox modulation of plant UGPase, as shown here for the enzymes from sugarcane and barley, adds yet another mode of regulation to a long list of mechanisms affecting UGPase activity. There is a variety of abiotic factors that give rise to oxidative stress conditions, e.g., drought, high temperature, high light intensity, heavy metals, salinity, or ultraviolet radiation, and they all cause yield and quality losses in crops. Some of these abiotic factors are transient, as they are part of normal weather conditions, but some may persist and overlap during a plant life span [80]. With UGPase positioned at the very center of primary plant metabolism, its redox regulation may actually turn out to be a major mechanism controlling plant yield.
4. Materials and Methods
4.1. Materials
Constructs containing wt and Cys102Ser mutant of sugarcane UGPase were prepared as in [30,35] and were kindly provided by Dr. José Sérgio Soares, Functional Genome Lab., State Univ. Campinas, Brazil. Recombinant sugarcane UGPases were purified by the Umeå University Protein Expertise Platform (PEP) by immobilized metal (nickel) affinity chromatography, and the polyHis tail was cleaved off by TEV protease. Aliquots of purified enzyme, in concentrations of 0.3–0.45 mg/mL, were stored at −80 °C in 29 mM phosphate buffer (pH 8.0), 150 mM NaCl and 1 mM β-mercaptoethanol. Purified recombinant barley UGPase and its Cys99Ser mutant were produced as described in [36,37].
4.2. Assays
The recombinant sugarcane UGPase was assayed at room temperature (RT) in the reaction mixture containing 100 mM Hepes-NaOH (pH 7.5), 5 mM MgCl2, 0.6 mM NADP, 1 unit each of coupling enzymes: phosphoglucomutase and glucose 6-phosphate dehydrogenase. Unless otherwise stated, PPi and UDPG (substrates used) were kept at 1 mM and 2 mM, respectively. The reactions, at a final volume of 100 µL, were started with an aliquot of UGPase. Assays were performed in 96 well plates (Sarstedt, Numbrecht, Germany), and the formation of NADPH was determined at 340 nm on a Spectramax 190 plate reader (Molecular Devices, Sunnyvale, CA, USA). The assays of recombinant barley UGPase were carried out essentially as those for sugarcane enzyme, with the exception that, unless otherwise stated, UDPG and PPi were at 1 mM each.
One unit of UGPase activity refers to 1 µmol NADPH formed per minute under assay conditions, corresponding to 1 µmol of glucose-1-P (product of UGPase) formed during the reaction. Specific activities of sugarcane UGPase, as found in this study, were 480 unit/mg and 390 unit/mg for wt and Cys102Ser proteins, respectively; for barley UGPase, they were 1200 unit/mg for wt and 650 unit/mg protein for Cys99Ser mutant, respectively [36]. Assay chemicals were obtained from Sigma-Aldrich Sweden AB, Stockholm, Sweden.
4.3. Kinetic Studies
For the determination of Michaelis–Menten constant (Km) values for UDPG and PPi, assays were done by varying the concentration of one substrate at the constant fixed level of the other substrate. For Km determinations of sugarcane UGPase, fixed concentrations of substrates were: 2 mM UDPG and 1 mM PPi. For barley UGPase, both UDPG and PPi were fixed at 1 mM (for wt) and at 2 and 5 mM, respectively, for the Cys99Ser mutant. The Km values were calculated using the Excel solver add-in. Kinetic points were means of at least two repeats that were reproducible within ±10%. Prior to assays, sugarcane UGPase was pre-incubated overnight at RT either untreated or with 10 mM GSSG (oxidized enzyme). Before assays of barley UGPase, the enzyme was preincubated overnight at RT with 2 mM DTT (reduced enzyme) or 2 mM H2O2 (oxidized enzyme).
4.4. Structural Modeling
Barley UGPase 3D structure was homology-modeled, based on the crystal structure of Arabidopsis UGPase1 [34], as in [37,54]. Both the barley UGPase homology model and the crystal structure of sugarcane UGPase (pdb 5WEG) [35] were displayed by DeepView/Swiss-PdbViewer (v4.1) (https://spdbv.unil.ch/) [81].
Author Contributions
D.D. and L.A.K. designed the experiments. D.D., J.A. and L.A.K. carried out all experiments. M.W. analyzed the UGPase structure. D.D. and L.A.K. analyzed data. L.A.K. wrote the manuscript with input from D.D., J.A. and M.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the strategic fund from Umeå University (to L.A.K.) as well as by grants from the Stiftelsen J.C. Kempes Minnes Stipendiefond (to D.D.) and the Lars Hierta Memorial Foundation (to D.D.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Acknowledgments
We are very grateful to José Sérgio Soares, Functional Genome Lab., State Univ. State Univ. Campinas, Brazil, for providing us with cDNA clones for sugarcane UGPase.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zrenner, R.; Willmitzer, L.; Sonnewald, U. Analysis of the expression of potato uridinediphosphate-glucose pyrophosphorylase and its inhibition by antisense RNA. Planta 1993, 190, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Geisler, M.; Johansson, H.; Harholt, J.; Scheller, H.V.; Mellerowicz, E.J.; Kleczkowski, L.A. UDP-glucose pyrophosphorylase is not rate-limiting, but is essential in Arabidopsis. Plant Cell Physiol. 2009, 50, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Park, J.I.; Ishimizu, T.; Suwabe, K.; Sudo, K.; Masuko, H.; Hakozaki, H.; Nou, I.S.; Suzuki, G.; Watanabe, M. UDP-glucose pyrophosphorylase is rate limiting in vegetative and reproductive phases in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 981–996. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Decker, D. Sugar activation for production of nucleotide sugars as substrates for glycosyltransferases in plants. J. Appl. Glycosci. 2015, 62, 25–36. [Google Scholar] [CrossRef]
- Decker, D.; Kleczkowski, L.A. UDP-sugar producing pyrophosphorylases—Distinct and essential enzymes with overlapping substrate specificities, providing de novo precursors for glycosylation reactions. Front. Plant Sci. 2019, 9, 1822. [Google Scholar] [CrossRef] [PubMed]
- Stucki, J.W. The thermodynamic-buffer enzymes. Eur. J. Biochem. 1980, 109, 257–267. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Metabolic systems maintain stable non-equilibrium via thermodynamic buffering. BioEssays 2009, 31, 1091–1099. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Igamberdiev, A.U. Optimization of nucleotide sugar supply for polysaccharide formation via thermodynamic buffering. Biochem. J. 2020, 477, 341–356. [Google Scholar] [CrossRef]
- Feingold, D.S.; Avigad, G. Sugar nucleotide transformation in plants. In The Biochemistry of Plants; Stumpf, P.K., Conn, E.E., Eds.; Academic Press: New York, NY, USA, 1980; Volume 3, pp. 101–170. [Google Scholar] [CrossRef]
- Feingold, D.S.; Barber, G.A. Nucleotide sugars. In Methods in Plant Biochemistry; Dey, P.M., Harborne, J.B., Eds.; Academic Press: London, UK, 1990; Volume 2, pp. 39–78. [Google Scholar] [CrossRef]
- Knop, J.K.; Hansen, R.G. Uridine diphosphate glucose pyrophosphorylase. IV. Crystallization and properties of the enzyme from human liver. J. Biol. Chem. 1970, 245, 2499–2504. [Google Scholar] [CrossRef]
- Martínez, L.I.; Piattoni, C.V.; Garay, S.A.; Rodrígues, D.E.; Guerrero, S.A.; Iglesias, A.A. Redox regulation of UDP-glucose pyrophosphorylase from Entamoeba histolytica. Biochimie 2011, 93, 260–268. [Google Scholar] [CrossRef]
- Führing, J.A.; Cramer, J.T.; Schneider, J.; Baruch, P.; Gerardy-Schahn, R.; Fedorov, R. A quaternary mechanism enables the complex biological functions of octameric human UDP-glucose pyrophosphorylase, a key enzyme in cell metabolism. Sci. Rep. 2015, 5, 9618. [Google Scholar] [CrossRef] [PubMed]
- Nakano, K.; Omura, Y.; Tagaya, M.; Fukui, T. UDP-glucose pyrophosphorylase from potato tuber: Purification and characterization. J. Biochem. 1989, 106, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A. Glucose activation and metabolism through UDP-glucose pyrophosphorylase in plants. Phytochemistry 1994, 37, 1507–1515. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Geisler, M.; Ciereszko, I.; Johansson, H. UDP-glucose pyrophosphorylase. An old protein with new tricks. Plant Physiol. 2004, 134, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Elling, L.; Kula, M.R. Purification of UDP-glucose pyrophosphorylase from germinated barley (malt). J. Biotechnol. 1994, 34, 157–163. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Keech, O.; Gardeström, P.; Kleczkowski, L.A.; Rouhier, N. The redox control of photorespiration: From biochemical and physiological aspects to biotechnological considerations. Plant Cell Environ. 2017, 40, 553–569. [Google Scholar] [CrossRef]
- Ebrecht, A.C.; Asención Diez, M.D.; Piattoni, C.V.; Guerrero, S.A.; Iglesias, A.A. The UDP-glucose pyrophosphorylase from Giardia lamblia is redox regulated and exhibits promiscuity to use galactose-1-phosphate. Biochim. Biophys. Acta 2015, 1850, 88–96. [Google Scholar] [CrossRef]
- Muchut, R.J.; Calloni, R.D.; Herrera, F.E.; Garay, S.A.; Arias, D.G.; Iglesias, A.A.; Guerrero, S.A. Elucidating paramylon and other carbohydrate metabolism in Euglena gracilis: Kinetic characterization, structure and cellular localization of UDP-glucose pyrophosphorylase. Biochimie 2018, 154, 176–186. [Google Scholar] [CrossRef]
- Wong, J.H.; Cai, N.; Balmer, Y.; Tanaka, C.K.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin targets of developing wheat seeds identified by complementary proteomic approaches. Phytochemistry 2004, 65, 1629–1640. [Google Scholar] [CrossRef]
- Alkhalfioui, F.; Renard, M.; Vensel, W.H.; Wong, J.; Tanaka, C.K.; Hurkman, W.J.; Buchanan, B.B.; Montrichard, F. Thioredoxin-linked proteins are reduced during germination of Medicago truncatula seeds. Plant Physiol. 2007, 144, 1559–1579. [Google Scholar] [CrossRef]
- Collet, J.F.; Messens, J. Structure, function, and mechanism of thioredoxin proteins. Antioxid. Redox Signal. 2010, 13, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Skipsey, M.; Grundy, N.M.; Edwards, R. Stress-induced protein S-glutathionylation in Arabidopsis. Plant Physiol. 2005, 138, 2233–2244. [Google Scholar] [CrossRef]
- Wolosiuk, R.A.; Buchanan, B.B. Thioredoxin and glutathione regulate photosynthesis in chloroplasts. Nature 1977, 266, 565–567. [Google Scholar] [CrossRef]
- Lee, K.; Lee, J.; Kim, Y.; Bae, D.; Kang, K.Y.; Yoon, S.C.; Lim, D. Defining the plant disulfide proteome. Electrophoresis 2004, 25, 532–541. [Google Scholar] [CrossRef]
- Fratelli, M.; Gianazza, E.; Ghezzi, P. Redox proteomics: Identification and functional role of glutathionylated proteins. Expert Rev. Proteom. 2004, 1, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, P. Protein glutathionylation in health and disease. Biochim. Biophys. Acta 2013, 1830, 3165–3172. [Google Scholar] [CrossRef]
- Soares, J.S.; Gentile, A.; Scorsato, V.; Lima, A.C.; Kiyota, E.; Dos Santos, M.L.; Piattoni, C.V.; Huber, S.C.; Aparicio, R.; Menossi, M. Oligomerization, membrane association, and in vivo phosphorylation of sugarcane UDP-glucose pyrophosphorylase. J. Biol. Chem. 2014, 289, 33364–33377. [Google Scholar] [CrossRef]
- Karpinski, S.; Reynolds, H.; Karpinska, B.; Wingsle, G.; Creissen, G.; Mullineaux, P. Systemic signaling and acclimation in response to excess excitation energy in Arabidopsis. Science 1999, 284, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Randall, D.D. Light and thiol activation of maize leaf glycerate kinase. The stimulating effect of reduced thioredoxins and ATP. Plant Physiol. 1985, 79, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B. The path to thioredoxin and redox regulation in chloroplasts. Annu. Rev. Plant Biol. 2016, 67, 1–24. [Google Scholar] [CrossRef]
- McCoy, J.G.; Bitto, E.; Bingman, C.A.; Wesenberg, G.E.; Bannen, R.M.; Kondrashov, D.A.; Phillips, G.N. Structure and dynamics of UDPglucose pyrophosphorylase from Arabidopsis thaliana with bound UDPglucose and UTP. J. Mol. Biol. 2007, 366, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Cotrim, C.A.; Soares, J.S.M.; Kobe, B.; Menossi, M. Crystal structure and insights into the oligomeric state of UDP-glucose pyrophosphorylase from sugarcane. PLoS ONE 2018, 13, e0193667. [Google Scholar] [CrossRef]
- Martz, F.; Wilczynska, M.; Kleczkowski, L.A. Oligomerization status, with the monomer as active species, defines catalytic efficiency of UDP-glucose pyrophosphorylase. Biochem. J. 2002, 367, 295–300. [Google Scholar] [CrossRef]
- Meng, M.; Fitzek, E.; Gajowniczek, A.; Wilczynska, M.; Kleczkowski, L.A. Domain-specific determinants of catalysis/substrate binding and the oligomerization status of barley UDP-glucose pyrophosphorylase. Biochim. Biophys. Acta 2009, 1794, 1734–1742. [Google Scholar] [CrossRef]
- Geisler, M.; Wilczynska, M.; Karpinski, S.; Kleczkowski, L.A. Toward a blueprint for UDP-glucose pyrophosphorylase structure/function properties: Homology-modeling analyses. Plant Mol. Biol. 2004, 56, 783–794. [Google Scholar] [CrossRef]
- Lyublinskaya, O.; Antunes, F. Measuring intracellular concentration of hydrogen peroxide with the use of genetically encoded H2O2 biosensor HyPer. Redox Biol. 2019, 24, 101200. [Google Scholar] [CrossRef]
- Sies, H. Role of metabolic H2O2 generation: Redox signaling and oxidative stress. J. Biol. Chem. 2014, 289, 8735–8741. [Google Scholar] [CrossRef]
- Gechev, T.S.; Hille, J. Hydrogen peroxide as a signal controlling plant programmed cell death. J. Cell Biol. 2005, 168, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Marrs, K.A. The functions and regulation of glutathione S-transferases in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef]
- Lu, S.C. Regulation of glutathione synthesis. Mol. Asp. Med. 2009, 30, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Bohle, F.; Klaus, A.; Tegethof, H.; Schwarzländer, M.; Hochholdinger, F.; Meyer, A.J.; Acosta, I.F.; Müller-Schüssele, S.J. High robustness of cytosolic glutathione redox potential under combined salt and osmotic stress in barley as revealed by the biosensor Grx1-roGFP2. bioRxiv 2022. [Google Scholar] [CrossRef]
- Yan, S.; Tang, Z.; Su, W.; Sun, W. Proteomic analysis of salt stress-responsive proteins in rice roots. Proteomics 2005, 5, 235–244. [Google Scholar] [CrossRef]
- Carpentier, S.C.; Witters, E.; Laukens, K.; Van Onckelen, H.; Swennen, R.; Panis, B. Banana (Musa spp.) as a model to study the meristem proteome: Acclimation to osmotic stress. Proteomics 2007, 7, 92–105. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Decker, D. Effects of magnesium, pyrophosphate and phosphonates on pyrophosphorolytic reaction of UDP-glucose pyrophosphorylase. Plants 2022, 11, 1611. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B. Formation and functions of protein sulfenic acids. Curr. Protoc. Toxicol. 2003, 17, 1–15. [Google Scholar] [CrossRef]
- Iyer, B.R.; Mahalakshmi, R. Hydrophobic characteristic is energetically preferred for cysteine in a model membrane protein. Biophys. J. 2019, 117, 25–35. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Martz, F.; Wilczynska, M. Factors affecting oligomerization status of UDP-glucose pyrophosphorylase. Phytochemistry 2005, 66, 2815–2821. [Google Scholar] [CrossRef]
- Meng, M.; Wilczynska, M.; Kleczkowski, L.A. Molecular and kinetic characterization of two UDP-glucose pyrophosphorylases, products of distinct genes, from Arabidopsis. Biochim. Biophys. Acta 2008, 1784, 967–972. [Google Scholar] [CrossRef]
- Decker, D.; Meng, M.; Gornicka, A.; Hofer, A.; Wilczynska, M.; Kleczkowski, L.A. Substrate kinetics and substrate effects on the quaternary structure of barley UDP-glucose pyrophosphorylase. Phytochemistry 2012, 79, 39–45. [Google Scholar] [CrossRef]
- Führing, J.; Damerow, S.; Fedorov, R.; Schneider, J.; Münster-Kühnel, A.K.; Gerardy-Schahn, R. Octamerization is essential for enzymatic function of human UDP-glucose pyrophosphorylase. Glycobiology 2013, 23, 426–437. [Google Scholar] [CrossRef]
- Ciereszko, I.; Johansson, H.; Hurry, V.; Kleczkowski, L.A. Phosphate status affects the gene expression, protein content and enzymatic activity of UDP-glucose pyrophosphorylase in wild-type and pho mutants of Arabidopsis. Planta 2001, 212, 598–605. [Google Scholar] [CrossRef]
- Ciereszko, I.; Johansson, H.; Kleczkowski, L.A. Sucrose and light regulation of a cold-inducible UDP-glucose pyrophosphorylase gene via a hexokinase-independent and abscisic acid-insensitive pathway in Arabidopsis. Biochem. J. 2001, 354, 67–72. [Google Scholar] [CrossRef]
- Repetto, O.; Bestel-Corre, G.; Dumas-Gaudot, E.; Berta, G.; Gianinazzi-Pearson, V.; Gianinazzi, S. Targeted proteomics to identify cadmium-induced protein modifications in Glomus mosseae-inoculated pea roots. New Phytol. 2003, 157, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Ciereszko, I.; Johansson, H.; Kleczkowski, L.A. Interactive effects of phosphate deficiency, sucrose and light/dark conditions on gene expression of UDP-glucose pyrophosphorylase in Arabidopsis. J. Plant Physiol. 2005, 162, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Geisler, M.; Johansson, H.; Mellerowicz, E.J.; Karpinski, S.; Kleczkowski, L.A. Differential tissue/organ-dependent expression of two sucrose- and cold-responsive genes for UDP-glucose pyrophosphorylase in Populus. Gene 2007, 389, 186–195. [Google Scholar] [CrossRef]
- Vandenabeele, S.; Van Der Kelen, K.; Dat, J.; Gadjev, I.; Boonefaes, T.; Morsa, S.; Rottiers, P.; Slooten, L.; Van Montagu, M.; Zabeau, M.; et al. A comprehensive analysis of hydrogen peroxide-induced gene expression in tobacco. Proc. Natl. Acad. Sci. USA 2003, 100, 16113–16118. [Google Scholar] [CrossRef]
- Dat, J.F.; Pellinen, R.; Beeckman, T.; Kangasjärvi, J.; Langebartels, C.; Inzé, D.; Van Breusegem, F. Changes in hydrogen peroxide homeostasis trigger an active cell death process in tobacco. Plant J. 2003, 33, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.D.; Morris, P.C. A proteomic analysis of 14-3-3 binding proteins from developing barley grains. Proteomics 2006, 6, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Swatek, K.N.; Graham, K.; Agrawal, G.K.; Thelen, J.J. The 14-3-3 isoforms chi and epsilon differentially bind client proteins from developing Arabidopsis seed. J. Proteome Res. 2011, 10, 4076–4087. [Google Scholar] [CrossRef]
- Chen, R.; Zhao, X.; Shao, Z.; Zhu, L.; He, G. Multiple isoforms of UDP-glucose pyrophosphorylase in rice. Physiol. Plant. 2007, 129, 725–736. [Google Scholar] [CrossRef]
- Komatsu, S.; Yamada, E.; Furukawa, K. Cold stress changes the concanavalin A positive glycosylation pattern of proteins expressed in the basal parts of rice leaf sheaths. Amino Acids 2009, 36, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Silva-Sanchez, C.; Chen, S.; Li, J.; Chourey, P.S. A comparative glycoproteome study of developing endosperm in the hexose-deficient miniature1 (mn1) seed mutant and its wild type Mn1 in maize. Front. Plant Sci. 2014, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Ferjani, A.; Kawade, K.; Asaoka, M.; Oikawa, A.; Okada, T.; Mochizuki, A.; Maeshima, M.; Hirai, M.Y.; Saito, K.; Tsukaya, H. Pyrophosphate inhibits gluconeogenesis by restricting UDP-glucose formation in vivo. Sci. Rep. 2018, 8, 14696. [Google Scholar] [CrossRef]
- Decker, D.; Öberg, C.; Kleczkowski, L.A. Identification and characterization of inhibitors of UDP-glucose and UDP-sugar pyrophosphorylases for in vivo studies. Plant J. 2017, 90, 1093–1107. [Google Scholar] [CrossRef]
- Decker, D.; Kleczkowski, L.A. Substrate specificity and inhibitor sensitivity of plant UDP-sugar producing pyrophosphorylases. Front. Plant Sci. 2017, 8, 1610. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Geisler, M.; Fitzek, E.; Wilczynska, M. A common structural blueprint for plant UDP-sugar producing pyrophosphorylases. Biochem. J. 2011, 439, 375–379. [Google Scholar] [CrossRef]
- Rose, I.A. The state of magnesium in cells as estimated from the adenylate kinase equilibrium. Proc. Natl. Acad. Sci. USA 1968, 61, 1079–1086. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Pyrophosphate as an alternative energy currency in plants. Biochem. J. 2021, 478, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Kleczkowski, L.A. Magnesium and cell energetics in plants under anoxia. Biochem. J. 2011, 437, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Igamberdiev, A.U. Magnesium signaling in plants. Int. J. Mol. Sci. 2021, 22, 1159. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Igamberdiev, A.U. Magnesium and cell energetics: At the junction of metabolism of adenylate and non-adenylate nucleotides. J. Plant Physiol. 2023, 280, 153901. [Google Scholar] [CrossRef]
- Cakmak, I.; Kirkby, E.A. Role of magnesium in carbon partitioning and alleviating photooxidative damage. Physiol. Plant. 2008, 133, 692–704. [Google Scholar] [CrossRef]
- Kim, G.; Weiss, S.J.; Levine, R.L. Methionine oxidation and reduction in proteins. Biochim. Biophys. Acta 2014, 1840, 901–905. [Google Scholar] [CrossRef]
- Weber, J.; Senior, A.E. Location and properties of pyrophosphate-binding sites in Escherichia coli F1-ATPase. J. Biol. Chem. 1995, 270, 12653–12658. [Google Scholar] [CrossRef]
- Qamer, Z.; Chaudhary, M.T.; Du, X.; Hinze, L.; Azhar, M.T. Review of oxidative stress and antioxidative defense mechanisms in Gossypium hirsutum L. in response to extreme abiotic conditions. J. Cotton Res. 2021, 4, 9. [Google Scholar] [CrossRef]
- Guex, N.; Diemand, A.; Peitsch, M.C. Protein modelling for all. Trends Biochem. Sci. 1999, 24, 364–367. [Google Scholar] [CrossRef]
Figure 1.
Effects of oxidized glutathione (GSSG) and H2O2 on activities of wt and Cys102Ser (C102S) mutant of sugarcane UGPase. The proteins were incubated for 30 min (at RT) at a given concentration of the oxidant or in water (no oxidant).
Figure 1.
Effects of oxidized glutathione (GSSG) and H2O2 on activities of wt and Cys102Ser (C102S) mutant of sugarcane UGPase. The proteins were incubated for 30 min (at RT) at a given concentration of the oxidant or in water (no oxidant).
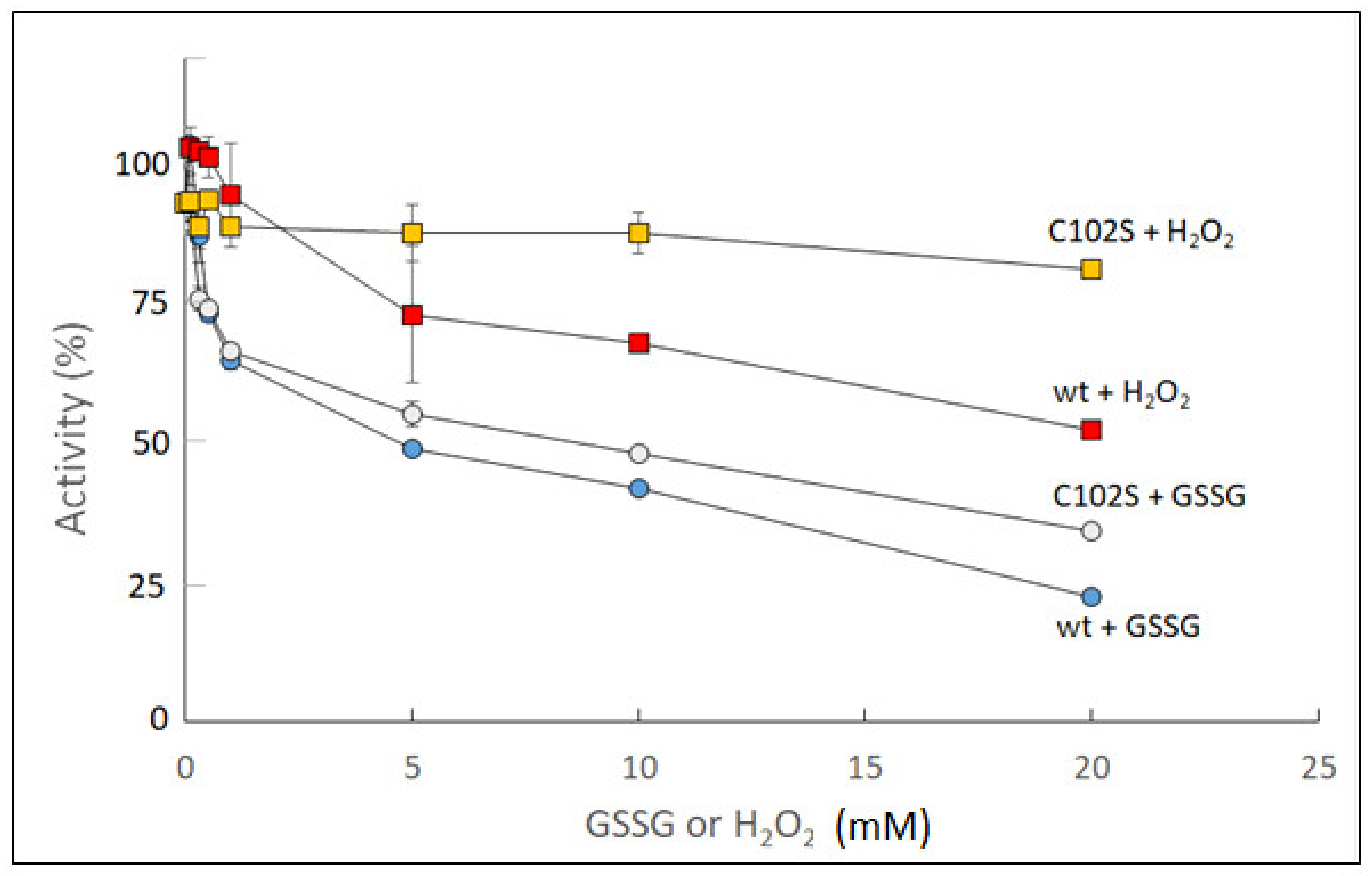
Figure 2.
Effects of dithiothreitol (DTT) and glutathione (GSH) on S-glutathionylated wt sugarcane UGPase (A) and its Cys102Ser (C102S) mutant (B). The proteins were incubated at RT for 30 min ± 10 mM GSSG (control). This was followed by treatment with 20 mM DTT or 20 mM GSH for 3 h.
Figure 2.
Effects of dithiothreitol (DTT) and glutathione (GSH) on S-glutathionylated wt sugarcane UGPase (A) and its Cys102Ser (C102S) mutant (B). The proteins were incubated at RT for 30 min ± 10 mM GSSG (control). This was followed by treatment with 20 mM DTT or 20 mM GSH for 3 h.
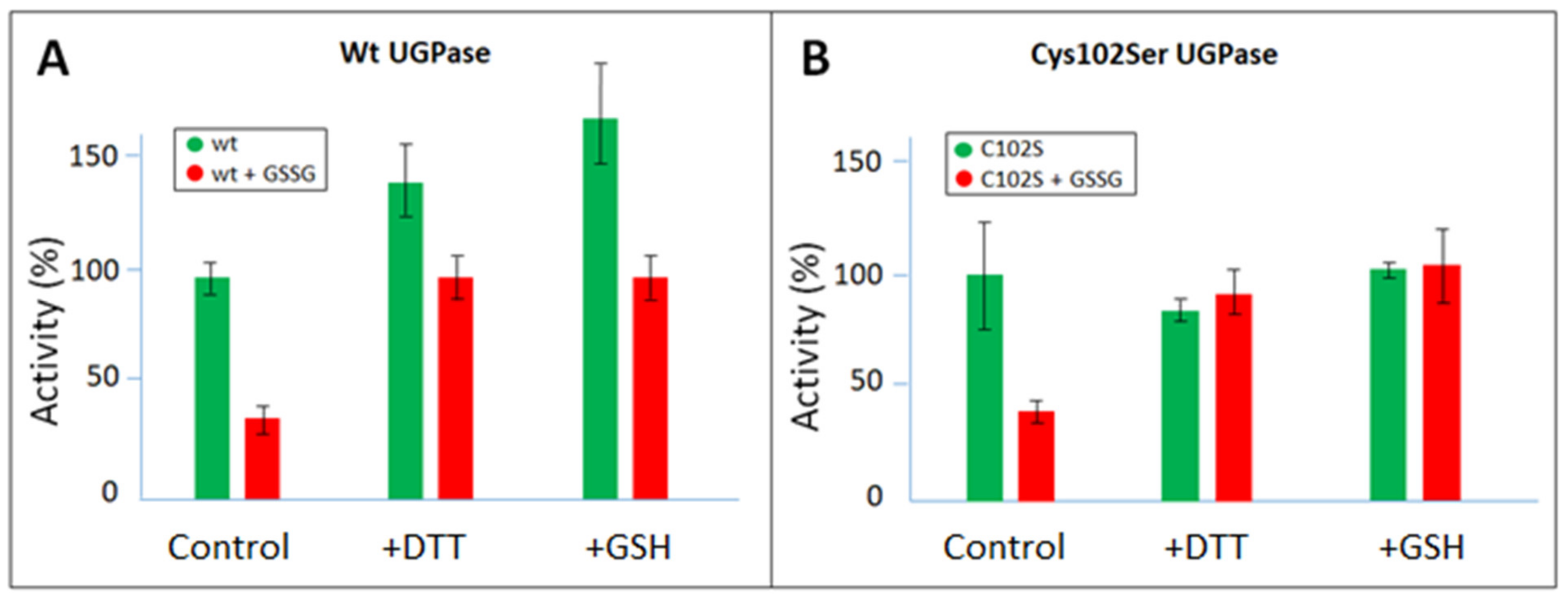
Figure 3.
Comparison of Km values for untreated and oxidized sugarcane wt UGPase and its Cys102Ser mutant (C102S). Panels (A,B) refer to Km values with UDPG and with PPi, respectively. The Kms were obtained by the double reciprocal plot method (see Section 4 for details). Prior to assays, the proteins were incubated overnight with either water (“untreated” enzyme, green boxes) or 10 mM GSSG (“oxidized” enzyme”, red boxes). The determined Km values are indicated on top of each box.
Figure 3.
Comparison of Km values for untreated and oxidized sugarcane wt UGPase and its Cys102Ser mutant (C102S). Panels (A,B) refer to Km values with UDPG and with PPi, respectively. The Kms were obtained by the double reciprocal plot method (see Section 4 for details). Prior to assays, the proteins were incubated overnight with either water (“untreated” enzyme, green boxes) or 10 mM GSSG (“oxidized” enzyme”, red boxes). The determined Km values are indicated on top of each box.
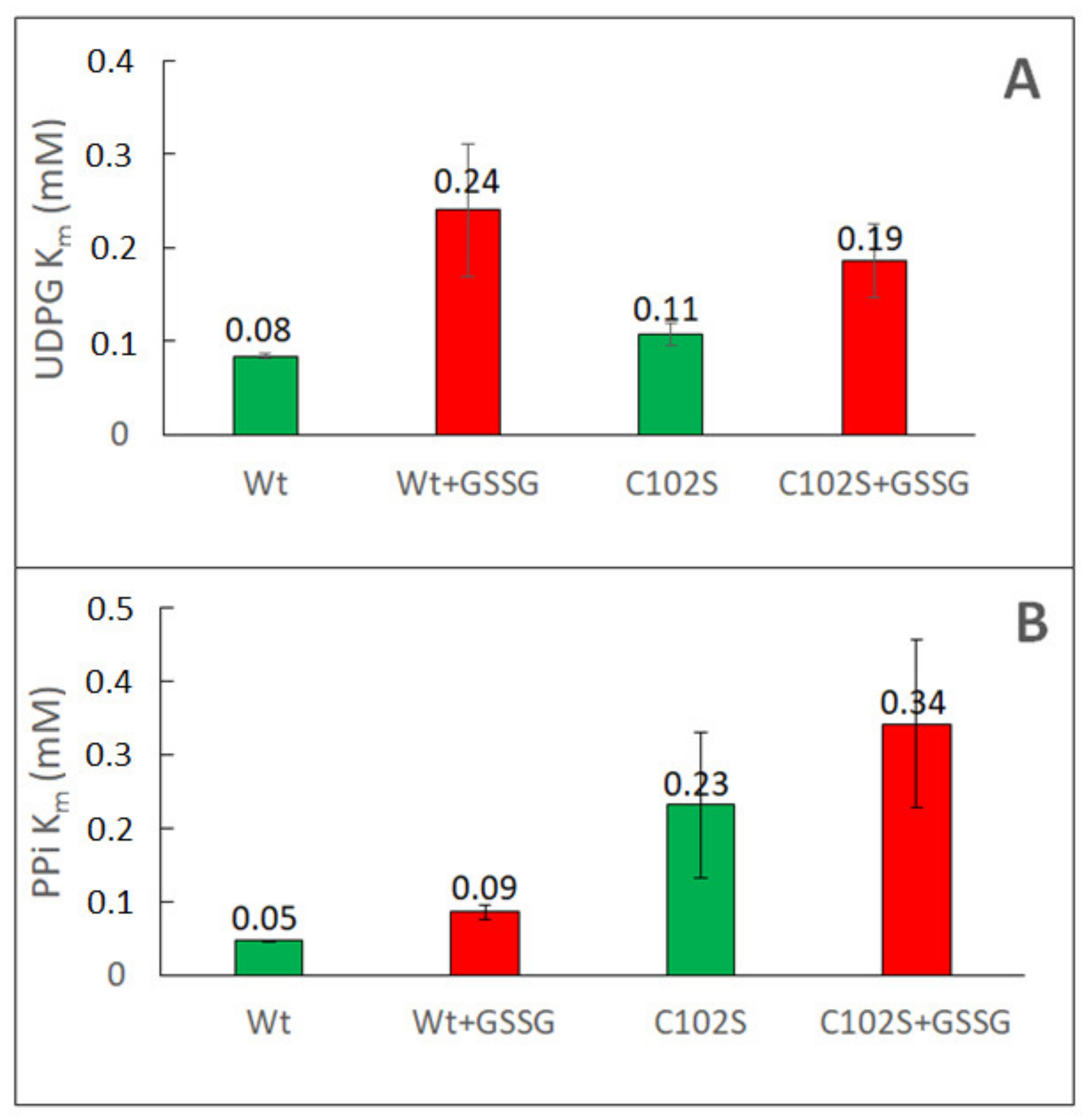
Figure 4.
Redox modulation of wt and Cys99Ser mutant of barley UGPase. The proteins were incubated at RT for 10 h either untreated or with 0.5 mM H2O2 (for wt) or 5 mM H2O2 (for Cys99Ser mutant), followed by incubation with 10 mM DTT for 40 min (at RT). Assays contained 0.85 mM UDPG and 0.5 mM PPi. Panels (A,B) refer to wt UGPase and the Cys99Ser mutant, respectively. The asterix (*) refers to statistical significance between the samples (p < 0.05).
Figure 4.
Redox modulation of wt and Cys99Ser mutant of barley UGPase. The proteins were incubated at RT for 10 h either untreated or with 0.5 mM H2O2 (for wt) or 5 mM H2O2 (for Cys99Ser mutant), followed by incubation with 10 mM DTT for 40 min (at RT). Assays contained 0.85 mM UDPG and 0.5 mM PPi. Panels (A,B) refer to wt UGPase and the Cys99Ser mutant, respectively. The asterix (*) refers to statistical significance between the samples (p < 0.05).
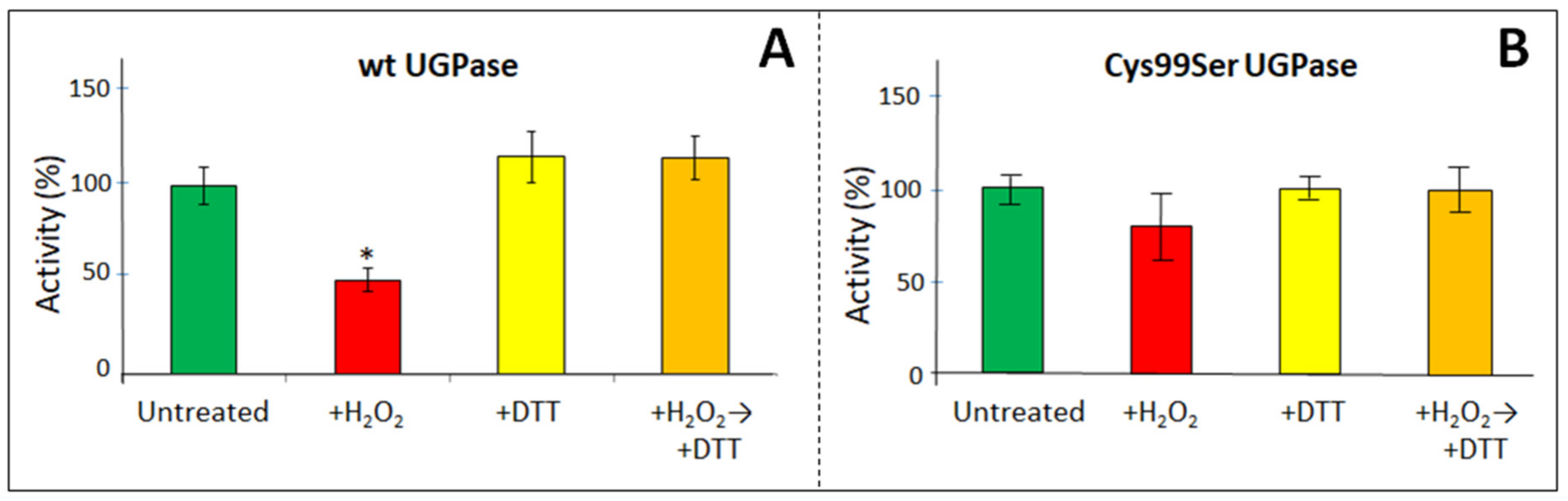
Figure 5.
Determination of Km values for barley wt UGPase and its Cys99Ser mutant (C99S) with UDPG (panels A,C) and PPi (panels B,D). Double reciprocal plots are shown, where UGPase activity (V) was measured at fixed concentrations of either PPi or UDPG, with the other substrate concentration varied. Prior to assays, the enzymes were incubated overnight at RT with either 2 mM H2O2 (“oxidized” enzyme) or 2 mM DTT (“reduced” enzyme). See Section 4 for other details.
Figure 5.
Determination of Km values for barley wt UGPase and its Cys99Ser mutant (C99S) with UDPG (panels A,C) and PPi (panels B,D). Double reciprocal plots are shown, where UGPase activity (V) was measured at fixed concentrations of either PPi or UDPG, with the other substrate concentration varied. Prior to assays, the enzymes were incubated overnight at RT with either 2 mM H2O2 (“oxidized” enzyme) or 2 mM DTT (“reduced” enzyme). See Section 4 for other details.
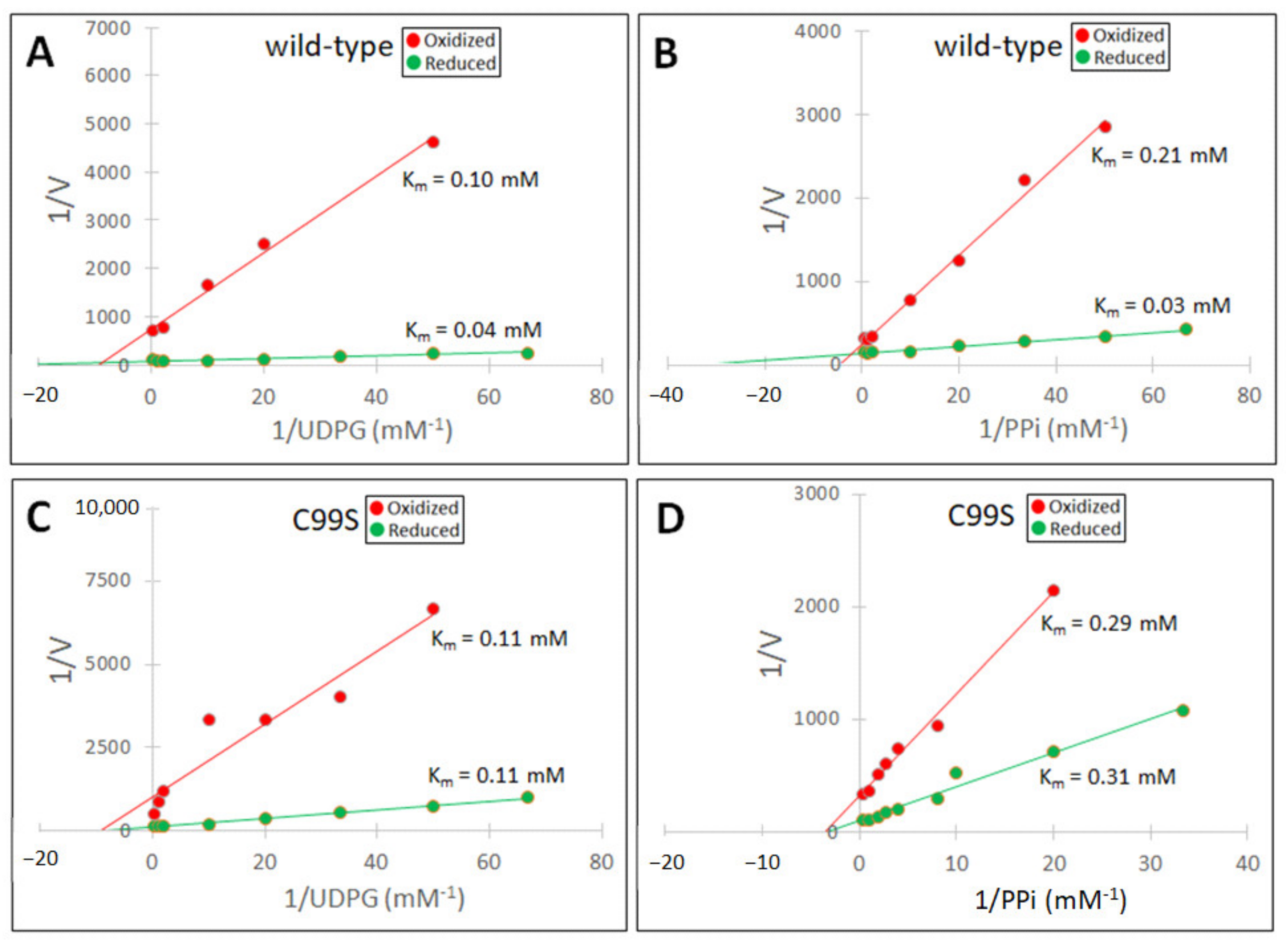
Figure 6.
Structures of sugarcane and barley UGPases. Structures shown are based on the crystal structure of sugarcane UGPase (pdb 5WEG) [35] and for homology-modeled barley UGPase, based on Arabidopsis UGPase crystal structure (pdb 1z90) [37]. (A) General view of sugarcane UGPase, with the N-terminal domain on the upper left (pink color), central domain (gray), and C-terminal domain on the lower right (orange). Shown also is the sugar-binding domain (green) and nucleotide-binding (NB) loop (red). The position and numbering of three cysteine molecules are indicated by blue balls corresponding to their α-carbons. (B) A similar view of barley UGPase, with its three Cys numbered and shown as blue balls. UGPase active site is composed of specific amino acids from the N-terminal domain, sugar-binding domain, central domain, NB loop, and C-terminal domain [34]. (C) The environment of Cys102 in sugarcane UGPase (pdb 5WEG) [35]. Shown are the closest amino acids at a distance of 4 Å from the sulfur atom of Cys 102. Broken red lines refer to distances from Cys102 to Ile109 (located on NB loop, central domain) and Val410 (C-terminal domain), which are likely to interplay with the Cys via hydrophobic bonds.
Figure 6.
Structures of sugarcane and barley UGPases. Structures shown are based on the crystal structure of sugarcane UGPase (pdb 5WEG) [35] and for homology-modeled barley UGPase, based on Arabidopsis UGPase crystal structure (pdb 1z90) [37]. (A) General view of sugarcane UGPase, with the N-terminal domain on the upper left (pink color), central domain (gray), and C-terminal domain on the lower right (orange). Shown also is the sugar-binding domain (green) and nucleotide-binding (NB) loop (red). The position and numbering of three cysteine molecules are indicated by blue balls corresponding to their α-carbons. (B) A similar view of barley UGPase, with its three Cys numbered and shown as blue balls. UGPase active site is composed of specific amino acids from the N-terminal domain, sugar-binding domain, central domain, NB loop, and C-terminal domain [34]. (C) The environment of Cys102 in sugarcane UGPase (pdb 5WEG) [35]. Shown are the closest amino acids at a distance of 4 Å from the sulfur atom of Cys 102. Broken red lines refer to distances from Cys102 to Ile109 (located on NB loop, central domain) and Val410 (C-terminal domain), which are likely to interplay with the Cys via hydrophobic bonds.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the redox effects on Km values of UGPases from sugarcane (wt and Cys102Ser) and barley (wt and Cys99Ser mutant). Compiled from Figure 3 and Figure 5.
| Sugarcane UGPase | Wild type | Cys102Ser | |||
| Untreated | Oxidized | Untreated | Oxidized | ||
| Km UDPG (mM) | 0.08 | 0.24 | 0.11 | 0.19 | |
| Km PPi (mM) | 0.04 | 0.09 | 0.23 | 0.34 | |
| Barley UGPase | Wild type | Cys99Ser | |||
| Reduced | Oxidized | Reduced | Oxidized | ||
| Km UDPG (mM) | 0.04 | 0.10 | 0.11 | 0.11 | |
| Km PPi (mM) | 0.03 | 0.21 | 0.31 | 0.29 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Decker, D.; Aubert, J.; Wilczynska, M.; Kleczkowski, L.A. Exploring Redox Modulation of Plant UDP-Glucose Pyrophosphorylase. Int. J. Mol. Sci. 2023, 24, 8914. https://doi.org/10.3390/ijms24108914
AMA Style
Decker D, Aubert J, Wilczynska M, Kleczkowski LA. Exploring Redox Modulation of Plant UDP-Glucose Pyrophosphorylase. International Journal of Molecular Sciences. 2023; 24(10):8914. https://doi.org/10.3390/ijms24108914
Chicago/Turabian StyleDecker, Daniel, Juliette Aubert, Malgorzata Wilczynska, and Leszek A. Kleczkowski. 2023. "Exploring Redox Modulation of Plant UDP-Glucose Pyrophosphorylase" International Journal of Molecular Sciences 24, no. 10: 8914. https://doi.org/10.3390/ijms24108914
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.