
Plant Growth Promotion Using Bacillus cereus

 , , ,
, , ,
Abstract
:1. Introduction
2. Biofilm Formation in Bacillus cereus
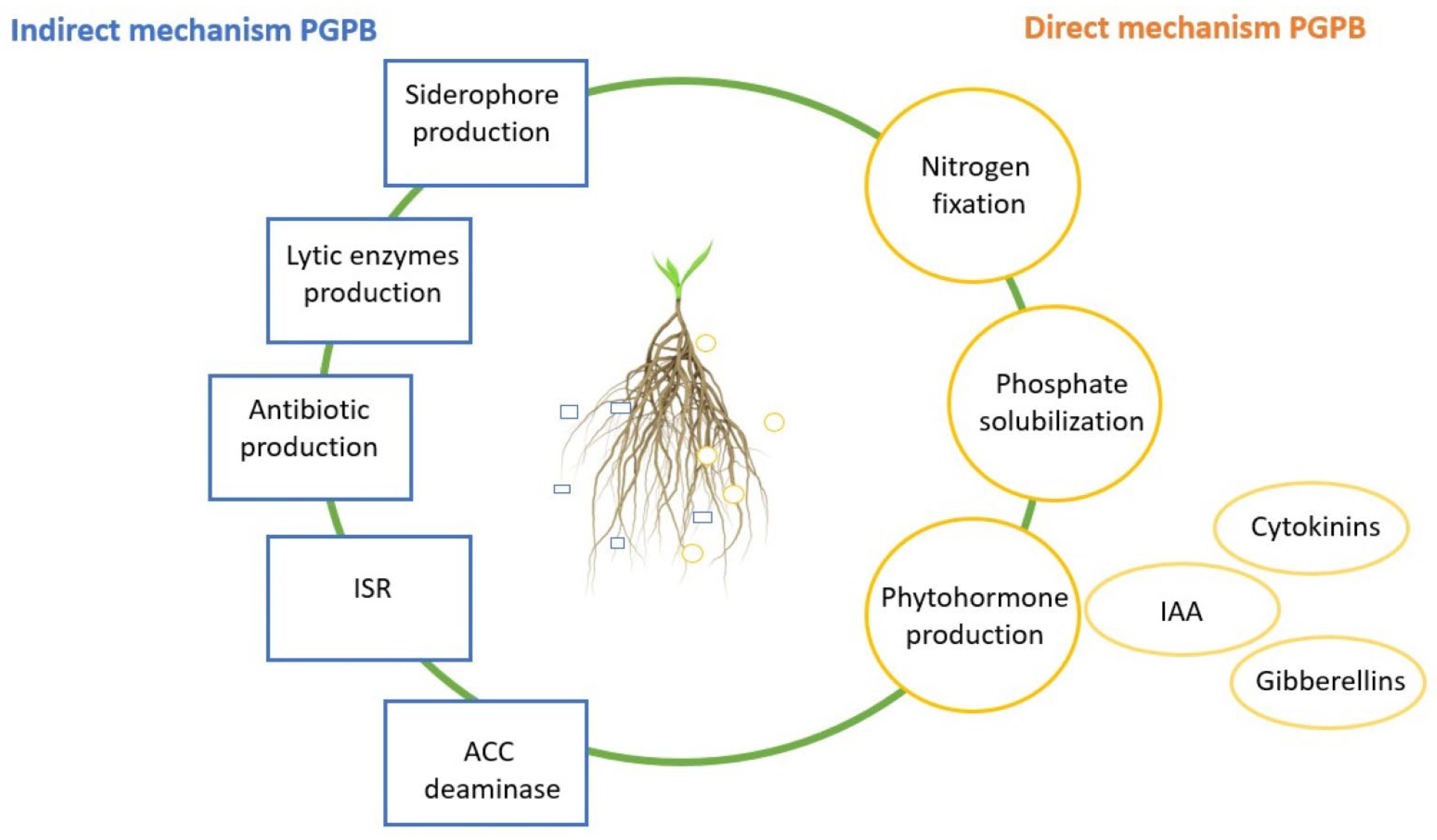
3. Bacillus cereus as a Directly Plant Growth Stimulating Bacteria
4. Plant Growth Promotion by Bacillus cereus in Combination with Other PGPB Species
5. Bacillus cereus in the Alleviation of Abiotic Stress
6. Biocontrol Using Bacillus cereus
7. Biocontrol Using Bacillus cereus in Combination with Other PGPB Species
8. Bacillus cereus Impact on Native Microbiota
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasperska-Wołowicz, W.; Rolbiecki, S.; Sadan, H.A.; Rolbiecki, R.; Jagosz, B.; Stachowski, P.; Liberacki, D.; Bolewski, T.; Prus, P.; Pal-Fam, F. Impact of the Projected Climate Change on Soybean Water Needs in the Kuyavia Region in Poland. J. Water Land Dev. 2021, 51, 199–207. [Google Scholar] [CrossRef]
- Wierzchowski, P.S.; Dobrzyński, J.; Mazur, K.; Kierończyk, M.; Wardal, W.J.; Sakowski, T.; Barszczewski, J. Chemical Properties and Bacterial Community Reaction to Acidified Cattle Slurry Fertilization in Soil from Maize Cultivation. Agronomy 2021, 11, 601. [Google Scholar] [CrossRef]
- Zielewicz, W.; Swędrzyński, A.; Dobrzyński, J.; Swędrzyńska, D.; Kulkova, I.; Wierzchowski, P.S.; Wróbel, B. Effect of Forage Plant Mixture and Biostimulants Application on the Yield, Changes of Botanical Composition, and Microbiological Soil Activity. Agronomy 2021, 11, 1786. [Google Scholar] [CrossRef]
- Dobrzyński, J.; Wierzchowski, P.S.; Stępień, W.; Górska, E.B. The Reaction of Cellulolytic and Potentially Cellulolytic Spore-Forming Bacteria to Various Types of Crop Management and Farmyard Manure Fertilization in Bulk Soil. Agronomy 2021, 11, 772. [Google Scholar] [CrossRef]
- Heyi, E.A.; Dinka, M.O.; Mamo, G. Assessing the Impact of Climate Change on Water Resources of Upper Awash River Sub-Basin, Ethiopia. J. Water Land Dev. 2022, 52, 232–244. [Google Scholar] [CrossRef]
- Gupta, K.; Dubey, N.K.; Singh, S.P.; Kheni, J.K.; Gupta, S.; Varshney, A. Plant Growth-Promoting Rhizobacteria (PGPR): Current and Future Prospects for Crop Improvement. In Current Trends in Microbial Biotechnology for Sustainable Agriculture; Environmental and Microbial Biotechnology; Springer: Singapore, 2021; pp. 203–226. [Google Scholar] [CrossRef]
- Pathania, P.; Rajta, A.; Singh, P.C.; Bhatia, R. Role of Plant Growth-Promoting Bacteria in Sustainable Agriculture. Biocatal. Agric. Biotechnol. 2020, 30, 101842. [Google Scholar] [CrossRef]
- Santoyo, G.; Guzman-Guzman, P.; Parra-Cota, F.I.; de los Santos-Villalobos, S.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant growth stimulation by microbial consortia. Agronomy 2021, 11, 219. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [Green Version]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Ferrusquía-Jiménez, N.I.; González-Arias, B.; Rosales, A.; Esquivel, K.; Escamilla-Silva, E.M.; Ortega-Torres, A.E.; Guevara-González, R.G. Elicitation of Bacillus cereus-Amazcala (B.c-A) with SiO2 Nanoparticles Improves Its Role as a Plant Growth-Promoting Bacteria (PGPB) in Chili Pepper Plants. Plants 2022, 11, 3445. [Google Scholar] [CrossRef]
- Sarmiento-López, L.G.; López-Meyer, M.; Maldonado-Mendoza, I.E.; Quiroz-Figueroa, F.R.; Sepúlveda-Jiménez, G.; Rodríguez-Monroy, M. Production of Indole-3-Acetic Acid by Bacillus Circulans E9 in a Low-Cost Medium in a Bioreactor. J. Biosci. Bioeng. 2022, 134, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyński, J.; Wróbel, B.; Górska, E.B. Cellulolytic Properties of a Potentially Lignocellulose-Degrading Bacillus sp. 8E1A Strain Isolated from Bulk Soil. Agronomy 2022, 12, 665. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.; Reddy, M.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 1, 1140. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, M.-Y.; Khan, N.; Tan, L.-L.; Yang, S. Potentials, Utilization, and Bioengineering of Plant Growth-Promoting Methylobacterium for Sustainable Agriculture. Sustainability 2021, 13, 3941. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The Role of Mycorrhizae and Plant Growth Promoting Rhizobacteria (PGPR) in Improving Crop Productivity under Stressful Environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7, 7–3. [Google Scholar] [CrossRef]
- Fayad, N.; Kallassy Awad, M.; Mahillon, J. Diversity of Bacillus cereus Sensu Lato Mobilome. BMC Genom. 2019, 20, 436. [Google Scholar] [CrossRef] [Green Version]
- Etikala, A.; Thamburaj, S.; Johnson, A.M.; Sarma, C.; Mummaleti, G.; Kalakandan, S.K. Incidence, Toxin Gene Profile, Antibiotic Resistance and Antibacterial Activity of Allium Parvum and Allium Cepa Extracts on Bacillus cereus Isolated from Fermented Millet-Based Food. LWT 2022, 160, 113314. [Google Scholar] [CrossRef]
- Ku, Y.; Xu, G.; Tian, X.; Xie, H.; Yang, X.; Cao, C. Root Colonization and Growth Promotion of Soybean, Wheat and Chinese Cabbage by Bacillus cereus YL6. PLoS ONE 2018, 13, e0200181. [Google Scholar] [CrossRef] [Green Version]
- Sansinenea, E. Applications and Patents of Bacillus spp. in Agriculture. In Intellectual Property Issues in Microbiology; Singh, H.B., Keswani, C., Singh, S.P., Eds.; Springer: Singapore, 2019; pp. 133–146. [Google Scholar] [CrossRef]
- Wijman, J.G.; de Leeuw, P.P.; Moezelaar, R.; Zwietering, M.H.; Abee, T. Air-Liquid Interface Biofilms of Bacillus cereus: Formation, Sporulation, and Dispersion. Appl. Environ. Microbiol. 2007, 73, 1481–1488. [Google Scholar] [CrossRef] [Green Version]
- Mohsin, M.Z.; Omer, R.; Huang, J.; Mohsin, A.; Guo, M.; Qian, J.; Zhuang, Y. Advances in Engineered Bacillus subtilis Biofilms and Spores, and Their Applications in Bioremediation, Biocatalysis, and Biomaterials. Sythn. Syst. Biotechnol. 2021, 6, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Ding, M.; Yang, C.-H.; Fan, H.; Chai, Y.; Li, Y. The Phosphotransferase System Gene PtsH Plays an Important Role in MnSOD Production, Biofilm Formation, Swarming Motility, and Root Colonization in Bacillus cereus 905. Res. Microbiol. 2019, 170, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Majed, R.; Faille, C.; Kallassy, M.; Gohar, M. Bacillus cereus Biofilms—Same, Only Different. Front. Microbiol. 2016, 7, 1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, T.; Foulston, L.; Chai, Y.; Wang, Q.; Losick, R. Alternative Modes of Biofilm Formation by Plant-Associated Bacillus cereus. Microbiol. Open 2015, 4, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Briandet, R.; Kovács, Á.T. Bacillus cereus Sensu Lato Biofilm Formation and Its Ecological Importance. Biofilm 2022, 4, 100070. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Yu, Y.; Gozzi, K.; Chen, Y.; Guo, J.; Chai, Y. Genome-Wide Investigation of Biofilm Formation in Bacillus cereus. Appl. Environ. Microbiol. 2017, 83, e00561-17. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Yu, Y.; Wang, L.; Luo, Y.; Guo, J.; Chai, Y. The ComER Gene Plays an Important Role in Biofilm Formation and Sporulation in Both Bacillus subtilis and Bacillus cereus. Front. Microbiol. 2016, 7, 1025. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yang, N.; Zheng, S.; Yan, F.; Jiang, C.; Yu, Y.; Guo, J.; Chai, Y.; Chen, Y. The Spo0A-SinI-SinR Regulatory Circuit Plays an Essential Role in Biofilm Formation, Nematicidal Activities, and Plant Protection in Bacillus cereus AR156. Mol. Plant Microbe Inter. 2017, 30, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Zhang, Z.; Liu, Q.; Liu, F.; Liu, Y.; Zhang, J.; Wang, G. SpoVG Is an Important Regulator of Sporulation and Affects Biofilm Formation by Regulating Spo0A Transcription in Bacillus cereus 0–9. BMC Microbiol. 2021, 21, 172. [Google Scholar] [CrossRef]
- Xu, Y.-B.; Chen, M.; Zhang, Y.; Wang, M.; Wang, Y.; Huang, Q.; Wang, X.; Wang, G. The Phosphotransferase System Gene PtsI in the Endophytic Bacterium Bacillus cereus Is Required for Biofilm Formation, Colonization, and Biocontrol against Wheat Sharp Eyespot. FEMS Microbiol. Lett. 2014, 354, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, H.; Huang, Q.; Zhang, Y.; Zhao, L.; Liu, F.; Wang, G. Four Superoxide Dismutases of Bacillus cereus 0–9 Are Non-Redundant and Perform Different Functions in Diverse Living Conditions. World J. Microbiol. Biotechnol. 2020, 36, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Meng, L.; Zhang, Y.; Sang, L.; Liu, Q.; Zhao, L.; Liu, F.; Wang, G. GapB is involved in biofilm formation dependent on LrgAB but not the SinI/R system in Bacillus cereus 0–9. Front. Microbiol. 2020, 11, 591926. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, H.; Xie, T.; Huang, Q.; Xiong, X.; Liu, Q.; Wang, G. The YmdB protein regulates biofilm formation dependent on the repressor SinR in Bacillus cereus 0–9. World J. Microbiol. Biotechnol. 2020, 36, 165. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Li, Y.; Ding, M.; Chai, Y.; Wang, Q. The phosphotransferase system gene ptsI in Bacillus cereus regulates expression of sodA2 and contributes to colonization of wheat roots. Res. Microbiol. 2017, 168, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Ding, M.; Wang, Q. The recA gene is crucial to mediate colonization of Bacillus cereus 905 on wheat roots. Appl. Microbiol. Biotechnol. 2020, 104, 9251–9265. [Google Scholar] [CrossRef]
- Gao, T.-T.; Ding, M.-Z.; Li, Y.; Zeng, Q.-C.; Wang, Q. Identification of genes involved in regulating MnSOD2 production and root colonization in Bacillus cereus 905. J. Integr. Agric. 2021, 20, 1570–1584. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent. Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Swiecicka, I. Natural Occurrence of Bacillus Thuringiensis and Bacillus cereus in Eukaryotic Organisms: A Case for Symbiosis. Biocontrol Sci. Technol. 2008, 18, 221–239. [Google Scholar] [CrossRef]
- Kumar, P.; Pahal, V.; Gupta, A.; Vadhan, R.; Chandra, H.; Dubey, R.C. Effect of Silver Nanoparticles and Bacillus cereus LPR2 on the Growth of Zea Mays. Sci. Rep. 2020, 10, 20409. [Google Scholar] [CrossRef]
- Ali, A.M.; Awad, M.Y.M.; Hegab, S.A.; Gawad, A.M.A.E.; Eissa, M.A. Effect of Potassium Solubilizing Bacteria (Bacillus cereus) on Growth and Yield of Potato. J. Plant Nutr. 2021, 44, 411–420. [Google Scholar] [CrossRef]
- Sherpa, M.T.; Bag, N.; Das, S.; Haokip, P.; Sharma, L. Isolation and Characterization of Plant Growth Promoting Rhizobacteria Isolated from Organically Grown High Yielding Pole Type Native Pea (Pisum sativum L.) Variety Dentami of Sikkim, India. Curr. Res. Microb. Sci. 2021, 2, 100068. [Google Scholar] [CrossRef]
- Ibrahim, M.S.; Ikhajiagbe, B. The Growth Response of Rice (Oryza sativa L. Var. FARO 44) in vitro after Inoculation with Bacterial Isolates from a Typical Ferruginous Ultisol. Bull. Natl. Res. Cent. 2021, 45, 70. [Google Scholar] [CrossRef]
- Baliyan, N.; Dhiman, S.; Dheeman, S.; Kumar, S.; Arora, N.K.; Maheshwari, D.K. Optimization of Gibberellic Acid Production in Endophytic Bacillus cereus Using Response Surface Methodology and Its Use as Plant Growth Regulator in Chickpea. J. Plant Growth Regul. 2022, 41, 3019–3029. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Ayangbenro, A.S.; Babalola, O.O. Genomic Analysis of Endophytic Bacillus cereus T4S and Its Plant Growth-Promoting Traits. Plants 2021, 10, 1776. [Google Scholar] [CrossRef]
- Lopes, R.; Tsui, S.; Gonçalves, P.J.R.O.; de Queiroz, M.V.A. Look into a Multifunctional Toolbox: Endophytic Bacillus Species Provide Broad and Underexploited Benefits for Plants. World J. Microbiol. Biotechnol. 2018, 34, 94. [Google Scholar] [CrossRef] [PubMed]
- Sivasankari, B.; Anandharaj, M. Isolation and Molecular Characterization of Potential Plant Growth Promoting Bacillus cereus GGBSTD1 and Pseudomonas spp. GGBSTD3 from Vermisources. Adv. Agric. 2014, 2014, e248591. [Google Scholar] [CrossRef] [Green Version]
- Elsayed, A.; Abdelsattar, A.M.; Heikal, Y.M.; El-Esawi, M.A. Synergistic Effects of Azospirillum Brasilense and Bacillus cereus on Plant Growth, Biochemical Attributes and Molecular Genetic Regulation of Steviol Glycosides Biosynthetic Genes in Stevia Rebaudiana. Plant Physiol. Biochem. 2022, 189, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.K.; Maheshwari, D.K.; Dheeman, S.; Bajpai, V.K. Termitarium-Inhabiting Bacillus spp. Enhanced Plant Growth and Bioactive Component in Turmeric (Curcuma longa L.). Curr. Microbiol. 2017, 74, 184–192. [Google Scholar] [CrossRef]
- Hassan, T.U.; Bano, A. Biofertilizer: A novel formulation for improving wheat growth, physiology and yield. Pak. J. Bot. 2016, 48, 2233–2241. [Google Scholar]
- Singh, V.K.; Singh, A.K.; Singh, P.P.; Kumar, A. Interaction of plant growth promoting bacteria with tomato under abiotic stress: A review. Agric. Ecosyst. Environ. 2018, 267, 129–140. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Thermotolerance Effect of Plant Growth-Promoting Bacillus cereus SA1 on Soybean during Heat Stress. BMC Microbiol. 2020, 20, 175. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ma, H.; Liu, B.; Du, Z.; Ma, B.; Jing, D. Effects of Plant Growth-Promoting Rhizobacteria on the Physioecological Characteristics and Growth of Walnut Seedlings under Drought Stress. Agronomy 2023, 13, 290. [Google Scholar] [CrossRef]
- Bhatt, S.; Pandhi, N.; Raghav, R. Improved Salt Tolerance and Growth Parameters of Groundnut (Arachis hypogaea L.) Employing Halotolerant Bacillus cereus SVSCD1 Isolated from Saurashtra Region, Gujarat. Ecol. Environ. Cos. 2020, 26, S199–S212. [Google Scholar]
- Mukhtar, T.; Rehman, S.U.; Smith, D.; Sultan, T.; Seleiman, M.F.; Alsadon, A.A.; Amna; Ali, S.; Chaudhary, H.J.; Solieman, T.H.I.; et al. Mitigation of Heat Stress in Solanum lycopersicum L. by ACC-Deaminase and Exopolysaccharide Producing Bacillus cereus: Effects on Biochemical Profiling. Sustainability 2020, 12, 2159. [Google Scholar] [CrossRef] [Green Version]
- Alotaibi, B.S.; Khan, M.; Shamim, S. Unraveling the Underlying Heavy Metal Detoxification Mechanisms of Bacillus Species. Microorganisms 2021, 9, 1628. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, Z.; Xi, Y.; Wang, X.; Chen, S.; He, M.; Chen, Y.; Guo, Y. Improvement of Salt Tolerance of Arabidopsis Thaliana Seedlings Inoculated with Endophytic Bacillus cereus KP120. J. Plant Interact. 2022, 17, 884–893. [Google Scholar] [CrossRef]
- Azeem, M.; Haider, M.Z.; Javed, S.; Saleem, M.H.; Alatawi, A. Drought Stress Amelioration in Maize (Zea mays L.) by Inoculation of Bacillus spp. Strains under Sterile Soil Conditions. Agriculture 2022, 12, 50. [Google Scholar] [CrossRef]
- Andy, A.K.; Rajput, V.D.; Burachevskaya, M.; Gour, V.S. Exploring the Identity and Properties of Two Bacilli Strains and Their Potential to Alleviate Drought and Heavy Metal Stress. Horticulturae 2023, 9, 46. [Google Scholar] [CrossRef]
- Meenakshi; Annapurna, K.; Govindasamy, V.; Ajit, V.; Choudhary, D.K. Mitigation of Drought Stress in Wheat Crop by Drought Tolerant Endophytic Bacterial Isolates. Vegetos 2019, 32, 486–493. [Google Scholar] [CrossRef]
- Wróbel, M.; Śliwakowski, W.; Kowalczyk, P.; Kramkowski, K.; Dobrzyński, J. Bioremediation of Heavy Metals by the Genus Bacillus. Int. J. Environ. Res. Public Health 2023, 20, 4964. [Google Scholar] [CrossRef]
- Martins, S.J.; Rocha, G.A.; de Melo, H.C.; de Castro Georg, R.; Ulhôa, C.J.; de Campos Dianese, É.; Oshiquiri, L.H.; da Cunha, M.G.; da Rocha, M.R.; de Araújo, L.G.; et al. Plant-Associated Bacteria Mitigate Drought Stress in Soybean. Environ. Sci. Pollut. Res. 2018, 25, 13676–13686. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Babar, M.A. The Stimulatory Effects of Plant Growth Promoting Rhizobacteria and Plant Growth Regulators on Wheat Physiology Grown in Sandy Soil. Arch. Microbiol. 2019, 201, 769–785. [Google Scholar] [CrossRef] [PubMed]
- Hassan, T.U.; Bano, A.; Naz, I.; Hussain, M. Bacillus cereus: A competent plant growth promoting bacterium of saline sodic field. Pak. J. Bot. 2018, 50, 1029–1037. [Google Scholar]
- Zhou, Y.; Sang, T.; Tian, M.; Jahan, M.S.; Wang, J.; Li, X.; Guo, S.; Liu, H.; Wang, Y.; Shu, S. Effects of Bacillus cereus on Photosynthesis and Antioxidant Metabolism of Cucumber Seedlings under Salt Stress. Horticulturae 2022, 8, 463. [Google Scholar] [CrossRef]
- Wang, Q.; Peng, X.; Lang, D.; Ma, X.; Zhang, X. Physio-Biochemical and Transcriptomic Analysis Reveals That the Mechanism of Bacillus cereus G2 Alleviated Oxidative Stress of Salt-Stressed Glycyrrhiza Uralensis Fisch. Seedlings. Ecotoxicol. Environ. Saf. 2022, 247, 114264. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, Z.; Irshad, F.; Habib, A.; Hussain, N.; Sajjad, M.; Mumtaz, S.; Rehman, S.; Haider, W.; Hassan, M.N. Alleviation of cadmium stress in rice by inoculation of Bacillus cereus. PeerJ 2022, 10, e13131. [Google Scholar] [CrossRef] [PubMed]
- Sahile, A.A.; Khan, M.A.; Hamayun, M.; Imran, M.; Kang, S.-M.; Lee, I.-J. Novel Bacillus cereus Strain, ALT1, Enhance Growth and Strengthens the Antioxidant System of Soybean under Cadmium Stress. Agronomy 2021, 11, 404. [Google Scholar] [CrossRef]
- Akhtar, N.; Ilyas, N.; Yasmin, H.; Sayyed, R.Z.; Hasnain, Z.A.; Elsayed, E.; El Enshasy, H.A. Role of Bacillus cereus in Improving the Growth and Phytoextractability of Brassica nigra (L.) K. Koch in Chromium Contaminated Soil. Molecules 2021, 26, 1569. [Google Scholar] [CrossRef]
- Abou-Aly, H.E.; Youssef, A.M.; Tewfike, T.A.; El-Alkshar, E.A.; El-Meihy, R.M. Reduction of Heavy Metals Bioaccumulation in Sorghum and Its Rhizosphere by Heavy Metals-Tolerant Bacterial Consortium. Biocatal. Agric. Biotechnol. 2021, 31, 101911. [Google Scholar] [CrossRef]
- Yanti, Y.; Habazar, T.; Reflinaldon, R.; Nasution, C.R.; Felia, S. Indigenous Bacillus spp. Ability to Growth Promoting Activities and Control Bacterial Wilt Disease (Ralstonia solanacearum). Biodivers. J. Biol. Divers. 2017, 18, 1562–1567. [Google Scholar] [CrossRef]
- Malik, M.S.; Haider, S.; Rehman, A.; Rehman, S.U.; Jamil, M.; Naz, I.; Anees, M. Biological Control of Fungal Pathogens of Tomato (Lycopersicon esculentum) by Chitinolytic Bacterial Strains. J. Basic Microbiol. 2022, 62, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Hyder, S.; Gondal, A.S.; Rizvi, Z.F.; Ahmad, R.; Alam, M.M.; Hannan, A.; Ahmed, W.; Fatima, N.; Inam-ul-Haq, M. Characterization of Native Plant Growth Promoting Rhizobacteria and Their Anti-Oomycete Potential against Phytophthora Capsici Affecting Chilli Pepper (Capsicum annum L.). Sci. Rep. 2020, 10, 13859. [Google Scholar] [CrossRef] [PubMed]
- Antil, S.; Kumar, R.; Pathak, D.V.; Kumar, A.; Panwar, A.; Kumari, A. Plant Growth-Promoting Rhizobacteria—Bacillus cereus KMT-5 and B. Megaterium KMT-8 Effectively Suppressed Meloidogyne Javanica Infection. Appl. Soil Ecol. 2022, 174, 104419. [Google Scholar] [CrossRef]
- Arif, S.; Liaquat, F.; Yang, S.; Shah, I.H.; Zhao, L.; Xiong, X.; Garcia, D.; Zhang, Y. Exogenous Inoculation of Endophytic Bacterium Bacillus cereus Suppresses Clubroot (Plasmodiophora brassicae) Occurrence in Pak Choi (Brassica Campestris sp. Chinensis L.). Planta 2021, 253, 25. [Google Scholar] [CrossRef]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 386. [Google Scholar] [CrossRef]
- Jiang, C.H.; Huang, Z.Y.; Xie, P.; Gu, C.; Li, K.; Wang, D.C.; Yu, Y.Y.; Fan, Z.H.; Wang, C.J.; Wang, Y.P.; et al. Transcription factors WRKY70 and WRKY11 served as regulators in rhizobacterium Bacillus cereus AR156-induced systemic resistance to Pseudomonas syringae pv. tomato DC3000 in Arabidopsis. J. Exp. Bot. 2016, 67, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Fan, Z.; Li, Z.; Niu, D.; Li, Y.; Zheng, M.; Wang, Q.; Jin, H.; Guo, J. Bacillus cereus AR156 Triggers Induced Systemic Resistance against Pseudomonas Syringae Pv. Tomato DC3000 by Suppressing MiR472 and Activating CNLs-Mediated Basal Immunity in Arabidopsis. Mol. Plant Pathol. 2020, 21, 854–870. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Huerta, J.; Tamez-Guerra, P.; Gomez-Flores, R.; Delgado-Gardea, M.C.E.; Robles-Hernández, L.; Gonzalez-Franco, A.C.; Infante-Ramirez, R. Pepper growth promotion and biocontrol against Xanthomonas euvesicatoria by Bacillus cereus and Bacillus thuringiensis formulations. PeerJ 2023, 11, e14633. [Google Scholar] [CrossRef]
- Ananda, M.; Rusmana, I.; Akhdiya, A. Quorum quenching of Bacillus cereus INT1c against Pseudomonas syringae. J. Phys. Conf. Ser. 2019, 1277, 012010. [Google Scholar] [CrossRef] [Green Version]
- Rajesh, P.S.; Rai, V.R. Inhibition of QS-regulated virulence factors in Pseudomonas aeruginosa PAO1 and Pectobacterium carotovorum by AHL-lactonase of endophytic bacterium Bacillus cereus VT96. Biocatal. Agric. Biotechnol. 2016, 7, 154–163. [Google Scholar] [CrossRef]
- Wang, N.; Wang, L.; Zhu, K.; Hou, S.; Chen, L.; Mi, D.; Gui, Y.; Qi, Y.; Jiang, C.; Guo, J.-H. Plant Root Exudates Are Involved in Bacillus cereus AR156 Mediated Biocontrol Against Ralstonia solanacearum. Front. Microbiol. 2019, 10, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Dheeman, S.; Dubey, R.C.; Maheshwari, D.K.; Baliyan, N. Cyclic siloxane biosurfactant-producing Bacillus cereus BS14 biocontrols charcoal rot pathogen Macrophomina phaseolina and induces growth promotion in Vigna mungo L. Arch. Microbiol. 2021, 203, 5043–5054. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, D.; Thomas, A.; Banwarie, N. Antagonistic potential of lipopeptide producing Bacillus amyloliquefaciens against major vegetable pathogens. Eur. J. Plant Pathol. 2019, 154, 319–335. [Google Scholar] [CrossRef]
- Sarangi, T.; Ramakrishnan, S.; Nakkeeran, S. Antimicrobial Peptide Genes Present in Indigenous Isolates of Bacillus spp. Exhibiting Antimicrobical Properties. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1361–1369. [Google Scholar] [CrossRef] [Green Version]
- Ramarathnam, R.; Fernando, W.D. Molecular and biochemical detection of lipopeptide antibiotics producing Bacillus spp., antagonistic to common fungal pathogens of canola (Brassica napus L.). In Proceedings of the 12th International Rapeseed Congress, Wuhan, China, 26–30 March 2007; p. 180. [Google Scholar]
- Amruta, N.; Kumar, M.P.; Puneeth, M.E.; Sarika, G.; Kandikattu, H.K.; Vishwanath, K.; Narayanaswamy, S. Exploring the potentiality of novel rhizospheric bacterial strains against the rice blast fungus Magnaporthe oryzae. Plant Pathol. J. 2018, 34, 126. [Google Scholar] [CrossRef] [PubMed]
- Urdaci, M.; Pinchuk, I. Antimicrobial Activity of Bacillus Probiotics. In Bacterial Spore Formers—Probiotics and Emerging Applications; Horizon Bioscience: Norfolk, UK, 2004; pp. 171–182. [Google Scholar]
- Chauhan, A.K.; Maheshwari, D.K.; Kim, K.; Bajpai, V.K. Termitarium-inhabiting Bacillus endophyticus TSH42 and Bacillus cereus TSH77 colonizing Curcuma longa L.: Isolation, characterization, and evaluation of their biocontrol and plant-growth-promoting activities. Can. J. Microbiol. 2016, 62, 880–892. [Google Scholar] [CrossRef]
- Ramarathnam, R.; Fernando, W.G.D.; de Kievit, T. The Role of Antibiosis and Induced Systemic Resistance, Mediated by Strains of Pseudomonas chlororaphis, Bacillus cereus and B. Amyloliquefaciens, in Controlling Blackleg Disease of Canola. BioControl 2011, 56, 225–235. [Google Scholar] [CrossRef]
- Płaza, G.; Chojniak, J.; Rudnicka, K.; Paraszkiewicz, K.; Bernat, P. Detection of biosurfactants in Bacillus species: Genes and products identification. J. Appl. Microbiol. 2015, 119, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, P.; Kashyap, P.L.; Srivastava, A.K.; Tiwari, R.K. Plant Growth Promoting and Antifungal Activity in Endophytic Bacillus Strains from Pearl Millet (Pennisetum glaucum). Braz. J. Microbiol. 2020, 51, 229–241. [Google Scholar] [CrossRef]
- Ajesh, K.; Sudarslal, S.; Arunan, C.; Sreejith, K. Kannurin, a novel lipopeptide from Bacillus cereus strain AK 1: Isolation, structural evaluation and antifungal activities. J. Appl. Microbiol. 2013, 115, 1287–1296. [Google Scholar] [CrossRef]
- Abriouel, H.; Franz, C.M.; Omar, N.B.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Kuipers, O.P. Identification and Classification of Known and Putative Antimicrobial Compounds Produced by a Wide Variety of Bacillales Species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.-Y.; Si, F.-J.; Wang, N.; Wang, T.; Jin, Y.; Zheng, Y.; Yang, W.; Luo, Y.-M.; Niu, D.-D.; Guo, J.-H.; et al. Bacillus-Secreted Oxalic Acid Induces Tomato Resistance Against Gray Mold Disease Caused by Botrytis cinerea by Activating the JA/ET Pathway. Mol. Plant Microbe Interact. 2022, 35, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Madriz-Ordeñana, K.; Pazarlar, S.; Jørgensen, H.J.L.; Nielsen, T.K.; Zhang, Y.; Nielsen, K.L.; Hansen, L.H.; Thordal-Christensen, H. The Bacillus cereus Strain EC9 Primes the Plant Immune System for Superior Biocontrol of Fusarium oxysporum. Plants 2022, 11, 687. [Google Scholar] [CrossRef]
- Piechulla, B.; Lemfack, M.C.; Kai, M. Effects of Discrete Bioactive Microbial Volatiles on Plants and Fungi. Plant Cell Environ. 2017, 40, 2042–2067. [Google Scholar] [CrossRef]
- Ramírez, V.; Martínez, J.; del Bustillos-Cristales, M.R.; Catañeda-Antonio, D.; Munive, J.; Baez, A. Bacillus cereus MH778713 Elicits Tomato Plant Protection against Fusarium oxysporum. J. Appl. Microbiol. 2022, 132, 470–482. [Google Scholar] [CrossRef]
- Nie, P.; Li, X.; Wang, S.; Guo, J.; Zhao, H.; Niu, D. Induced systemic resistance against Botrytis cinerea by Bacillus cereus AR156 through a JA/ET- and NPR1-dependent signaling pathway and activates pamp-triggered immunity in Arabidopsis. Front. Plant Sci. 2017, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G.M.; Ramaraj, G.B.; Ponnusamy, P.; Sekar, J.; Ramalingam, P.V. Biocontrol efficiency of native plant growth promoting rhizobacteria against rhizome rot disease of turmeric. Biol. Control 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Zhou, H.; Ren, Z.; Zu, X.; Yu, X.; Zhu, H.; Li, X.; Zhong, J.; Liu, E. Efficacy of Plant Growth-Promoting Bacteria Bacillus cereus YN917 for Biocontrol of Rice Blast. Front. Microbiol. 2021, 12, 684888. [Google Scholar] [CrossRef]
- Janahiraman, V.; Anandham, R.; Kwon, S.W.; Sundaram, S.; Karthik Pandi, V.; Krishnamoorthy, R.; Kim, K.; Samaddar, S.; Sa, T. Control of Wilt and Rot Pathogens of Tomato by Antagonistic Pink Pigmented Facultative Methylotrophic Delftia Lacustris and Bacillus spp. Front. Plant Sci. 2016, 7, 1626. [Google Scholar] [CrossRef] [Green Version]
- Lizárraga-Sánchez, G.J.; Leyva-Madrigal, K.Y.; Sánchez-Peña, P.; Quiroz-Figueroa, F.R.; Maldonado-Mendoza, I.E. Bacillus cereus Sensu Lato Strain B25 Controls Maize Stalk and Ear Rot in Sinaloa, Mexico. Field Crops Res. 2015, 176, 11–21. [Google Scholar] [CrossRef]
- Figueroa-López, A.M.; Cordero-Ramírez, J.D.; Martínez-Álvarez, J.C.; López-Meyer, M.; Lizárraga-Sánchez, G.J.; Félix-Gastélum, R.; Castro-Martínez, C.; Maldonado-Mendoza, I.E. Rhizospheric Bacteria of Maize with Potential for Biocontrol of Fusarium verticillioides. SpringerPlus 2016, 5, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydi Ben Abdallah, R.; Mokni-Tlili, S.; Nefzi, A.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Biocontrol of Fusarium Wilt and Growth Promotion of Tomato Plants Using Endophytic Bacteria Isolated from Nicotiana Glauca Organs. Biol. Control 2016, 97, 80–88. [Google Scholar] [CrossRef]
- Hammami, I.; Siala, R.; Jridi, M.; Ktari, N.; Nasri, M.; Triki, M.A. Partial purification and characterization of chiIO8, a novel antifungal chitinase produced by Bacillus cereus IO8. J. Appl. Microbiol. 2013, 115, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-T.; Chen, Y.-C.; Jao, C.-L. Antifungal Activity and Enhancement of Plant Growth by Bacillus cereus Grown on Shellfish Chitin Wastes. Bioresour. Technol. 2007, 98, 1224–1230. [Google Scholar] [CrossRef]
- Li, J.-G.; Jiang, Z.-Q.; Xu, L.-P.; Sun, F.-F.; Guo, J.-H. Characterization of Chitinase Secreted by Bacillus cereus Strain CH2 and Evaluation of Its Efficacy against Verticillium Wilt of Eggplant. BioControl 2008, 53, 931–944. [Google Scholar] [CrossRef]
- Chen, W.; Wang, J.; Huang, D.; Cheng, W.; Shao, Z.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Volatile organic compounds from Bacillus aryabhattai MCCC 1K02966 with multiple modes against Meloidogyne incognita. Molecules 2022, 27, 103. [Google Scholar] [CrossRef]
- Duong, B.; Marraccini, P.; Etienne, H.; Villain, L.; Hoang, G.T.; Khong, G.N.; Lebrun, M.; Duponnois, R. Potential of the coffee endophytic Bacillus cereus sensu lato strain CCBLR15 to control the plant-parasitic nematode Radopholus duriophilus. Biocontrol Sci. Technol. 2022, 32, 971–988. [Google Scholar] [CrossRef]
- Wang, C.; Hu, H.J.; Li, X.; Wang, Y.F.; Tang, Y.Y.; Chen, S.L.; Yan, S.Z. Effects of varying environmental factors on the biological control of Meloidogyne incognita in tomato by Bacillus cereus strain BCM2. Biocontrol Sci. Technol. 2018, 28, 359–376. [Google Scholar] [CrossRef]
- Li, X.; Hu, H.-J.; Li, J.-Y.; Wang, C.; Chen, S.-L.; Yan, S.-Z. Effects of the Endophytic Bacteria Bacillus cereus BCM2 on Tomato Root Exudates and Meloidogyne incognita Infection. Plant Dis. 2019, 103, 1551–1558. [Google Scholar] [CrossRef]
- Hu, H.; Gao, Y.; Li, X.; Chen, S.; Yan, S.; Tian, X. Identification and nematicidal characterization of proteases secreted by endophytic bacteria Bacillus cereus BCM2. Phytopathology 2020, 110, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.J.; Tan, S.Y.; Shen, Q.R.; Ran, W. Bacillus cereus X5 suppresses root-knot: Nematode of tomato by colonizing in roots and soil. Afr. J. Microbiol. Res. 2012, 6, 2321–2327. [Google Scholar] [CrossRef]
- Yin, N.; Zhao, J.L.; Liu, R.; Li, Y.; Ling, J.; Yang, Y.-H.; Xie, B.-Y.; Mao, Z.-C. Biocontrol efficacy of Bacillus cereus strain Bc-cm103 against Meloidogyne incognita. Plant Dis. 2021, 105, 2061–2070. [Google Scholar] [CrossRef]
- Gao, H.; Qi, G.; Yin, R.; Zhang, H.; Li, C.; Zhao, X. Bacillus cereus strain S2 shows high nematicidal activity against Meloidogyne incognita by producing sphingosine. Sci. Rep. 2016, 6, 28756. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, Y.; Yuan, H.; Sun, B.; Li, H. Biological control of the cereal cyst nematode (Heterodera filipjevi) by Achromobacter xylosoxidans isolate 09X01 and Bacillus cereus isolate 09B18. Biol. Control 2016, 92, 1–6. [Google Scholar] [CrossRef]
- Naureen, Z.; Hafeez, F.Y.; Roberts, M.R.A. Consortium of Siderophore-Producing Bacterial Strains from the Rhizosphere of Rice Plants Induces Systemic Resistance in Rice against Sheath Blight Disease. Asp. Appl. Biol. 2009, 98, 17–18. [Google Scholar]
- Zhang, L.-N.; Wang, D.-C.; Hu, Q.; Dai, X.-Q.; Xie, Y.-S.; Li, Q.; Liu, H.-M.; Guo, J.-H. Consortium of Plant Growth-Promoting Rhizobacteria Strains Suppresses Sweet Pepper Disease by Altering the Rhizosphere Microbiota. Front. Microbiol. 2019, 10, 1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Zheng, L.; Liu, H.X.; Wang, K.B.; Yu, Y.Y.; Luo, Y.M.; Guo, J.H. Evaluation of the effectiveness of a consortium of three plant-growth promoting rhizobacteria for biocontrol of cotton Verticillium wilt. Biocontrol Sci. Technol. 2014, 24, 489–502. [Google Scholar] [CrossRef]
- Zhang, J.; Ahmed, W.; Dai, Z.; Zhou, X.; He, Z.; Wei, L.; Ji, G. Microbial Consortia: An Engineering Tool to Suppress Clubroot of Chinese Cabbage by Changing the Rhizosphere Bacterial Community Composition. Biology 2022, 11, 918. [Google Scholar] [CrossRef]
- Mülner, P.; Bergna, A.; Wagner, P.; Sarajlić, D.; Gstöttenmayr, B.; Dietel, K.; Grosch, R.; Cernava, T.; Berg, G. Microbiota Associated with Sclerotia of Soilborne Fungal Pathogens—A Novel Source of Biocontrol Agents Producing Bioactive Volatiles. Phytobiomes J. 2019, 3, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Rilling, J.I.; Acuña, J.J.; Nannipieri, P.; Cassan, F.; Maruyama, F.; Jorquera, M.A. Current Opinion and Perspectives on the Methods for Tracking and Monitoring Plant Growth—promoting Bacteria. Soil Biol. Biochem. 2019, 130, 205–219. [Google Scholar] [CrossRef]
- Manfredini, A.; Malusà, E.; Costa, C.; Pallottino, F.; Mocali, S.; Pinzari, F.; Canfora, L. Current Methods, Common Practices, and Perspectives in Tracking and Monitoring Bioinoculants in Soil. Front. Microbiol. 2021, 12, 698491. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyński, J.; Jakubowska, Z.; Dybek, B. Potential of Bacillus pumilus to Directly Promote Plant Growth. Front. Microbiol. 2022, 13, 1069053. [Google Scholar] [CrossRef]
- Kang, Y.; Shen, M.; Wang, H.; Zhao, Q. A possible mechanism of action of plant growth-promoting rhizobacteria (PGPR) strain Bacillus pumilus WP8 via regulation of soil bacterial community structure. J. Gen. Appl. Microbiol. 2013, 59, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, R.M.; Cueva-Yesquén, L.G.; Garboggini, F.F.; Desoignies, N.; Rigobelo, E.C. Inoculum Concentration and Mineral Fertilization: Effects on the Endophytic Microbiome of Soybean. Front. Microbiol. 2022, 13, 900980. [Google Scholar] [CrossRef]
- Azeem, M.; Hassan, T.U.; Tahir, M.I.; Ali, A.; Jeyasundar, P.G.S.A.; Hussain, Q.; Bashir, S.; Mehmood, S.; Zhang, Z. Tea Leaves Biochar as a Carrier of Bacillus cereus Improves the Soil Function and Crop Productivity. Appl. Soil Ecol. 2021, 157, 103732. [Google Scholar] [CrossRef]
- Win, K.T.; Okazaki, K.; Ohkama-Ohtsu, N.; Yokoyama, T.; Ohwaki, Y. Short-Term Effects of Biochar and Bacillus pumilus TUAT-1 on the Growth of Forage Rice and Its Associated Soil Microbial Community and Soil Properties. Biol. Fertil. Soils 2020, 56, 481–497. [Google Scholar] [CrossRef]
- Wang, X.; Xie, H.; Ku, Y.; Yang, X.; Chen, Y.; Yang, N.; Mei, X.; Cao, C. Chemotaxis of Bacillus cereus YL6 and its colonization of Chinese cabbage seedlings. Plant Soil 2020, 447, 413–430. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Inducing the Rhizosphere Microbiome by Biofertilizer Application to Suppress Banana Fusarium Wilt Disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strains B. cereus | Source | Plant Species | Experimental Conditions | Plant Disease and Effects | Lytic Enzymes | References | |||
|---|---|---|---|---|---|---|---|---|---|
| Cellulase | Chitinase | β-1,3-Glucanase | Protease | ||||||
| B. cereus YN917 | Rice leaf | Rice | Greenhouse | Antifungal activity against Magnaporthe oryzae; promoting seed germination and seedling plant growth | + | - | + | + | [103] |
| B. cereus PPB-1 | Phyllopla-ne of crop plants | Tomato | In vitro, greenhouse | Antifungal activity against Fusarium oxysporum, Sclerotium rolfsii, Pythium ultimum, and Rhizoctonia solani | - | + | + | - | [104] |
| B. cereus B25 | Rhizosphe-re of maize | Maize | In vitro, in planta, field | Antifungal activity against Fusarium verticillioides; increase in grain, yield, plant height | - | + | + | + | [105,106] |
| B. cereus S42 | Tobacco organs | Tomato | In vitro, in vivo, greenhouse | Antifungal activity against Fusarium wilt; increase in height of plants, fresh weight, root length, root fresh. | - | + | - | + | [107] |
| B. cereus IO8 | Soil | Tomato | In vitro and in vivo | Antifungal activity against Alternaria solani, Fusarium solani, Fusarium sambucinum, Alternaria citri, Penicillium occitanis, Aspergillus nidulans, Verticillium dahliae, and Botrytis cinerea | - | + | - | - | [108] |
| B. cereus QQ308 | Soil | Chinese cabbage | Pots on the balcony | Antifungal activity against Fusarium oxysporum, Fusarium solani, and Pythium ultimum; increase in total weight and total height | - | + | - | + | [109] |
| B. cereus CH2 | Rhizosphere of eggplant | Eggplant | Greenhouse | Activity against Verticillium wilt; increase in plant biomass, both fresh and dry | - | + | - | - | [110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulkova, I.; Dobrzyński, J.; Kowalczyk, P.; Bełżecki, G.; Kramkowski, K. Plant Growth Promotion Using Bacillus cereus. Int. J. Mol. Sci. 2023, 24, 9759. https://doi.org/10.3390/ijms24119759
Kulkova I, Dobrzyński J, Kowalczyk P, Bełżecki G, Kramkowski K. Plant Growth Promotion Using Bacillus cereus. International Journal of Molecular Sciences. 2023; 24(11):9759. https://doi.org/10.3390/ijms24119759
Chicago/Turabian StyleKulkova, Iryna, Jakub Dobrzyński, Paweł Kowalczyk, Grzegorz Bełżecki, and Karol Kramkowski. 2023. "Plant Growth Promotion Using Bacillus cereus" International Journal of Molecular Sciences 24, no. 11: 9759. https://doi.org/10.3390/ijms24119759